
Rational Design for the Complete Synthesis of Stevioside in Saccharomyces cerevisiae


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Reagents
2.2. DNA Manipulation
2.3. Strain Construction, Transformation, Screening, and Culture
2.4. Metabolite Extraction and Quantification
2.5. Fed-Batch Fermentation of Yeast Strain SST-302III-ST2
3. Results
3.1. Construction of S. cerevisiae to Produce Steviol
3.2. Studies on Diterpenoid Synthases
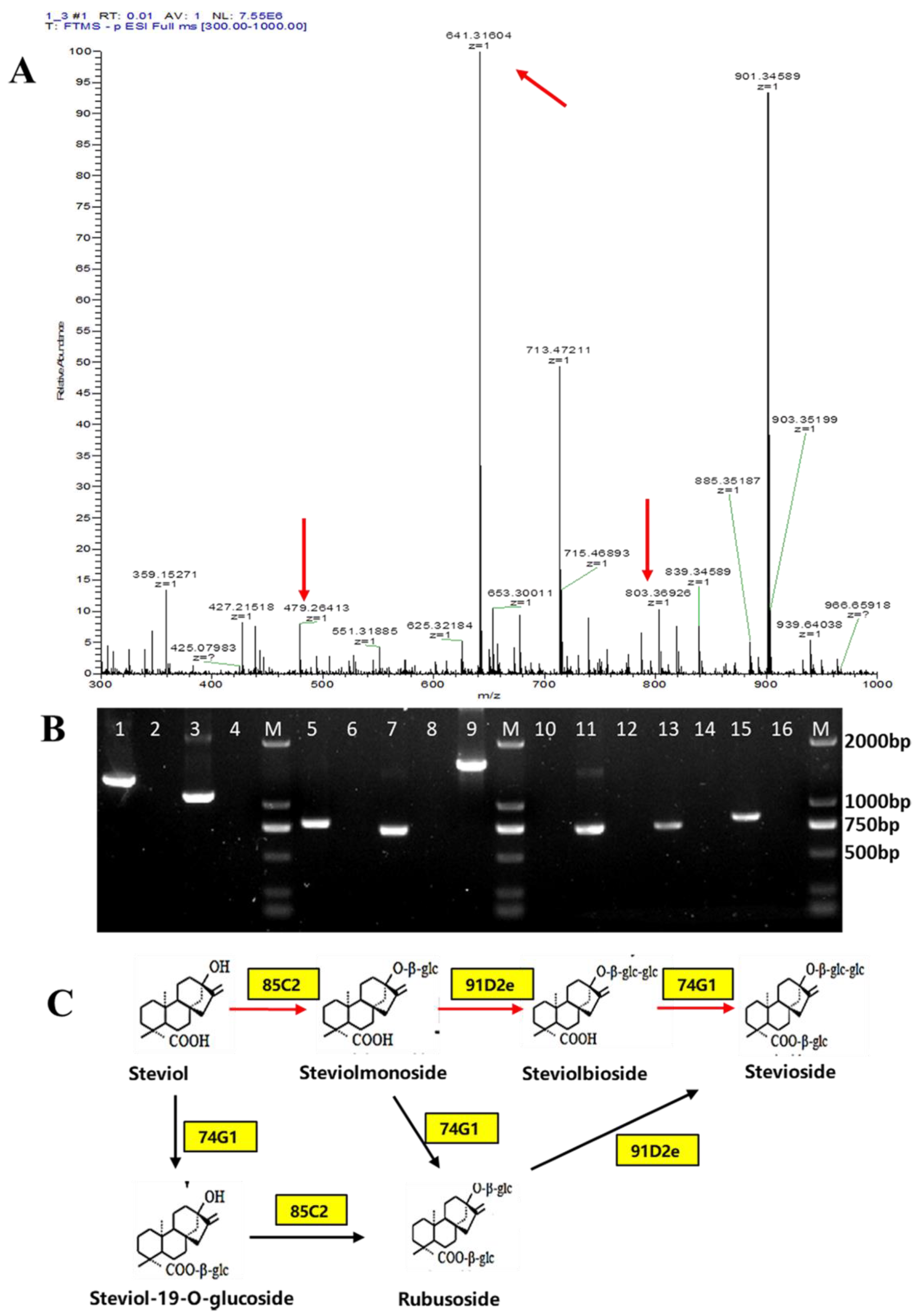
3.3. Complete Biosynthesis of Stevioside in S. cerevisiae
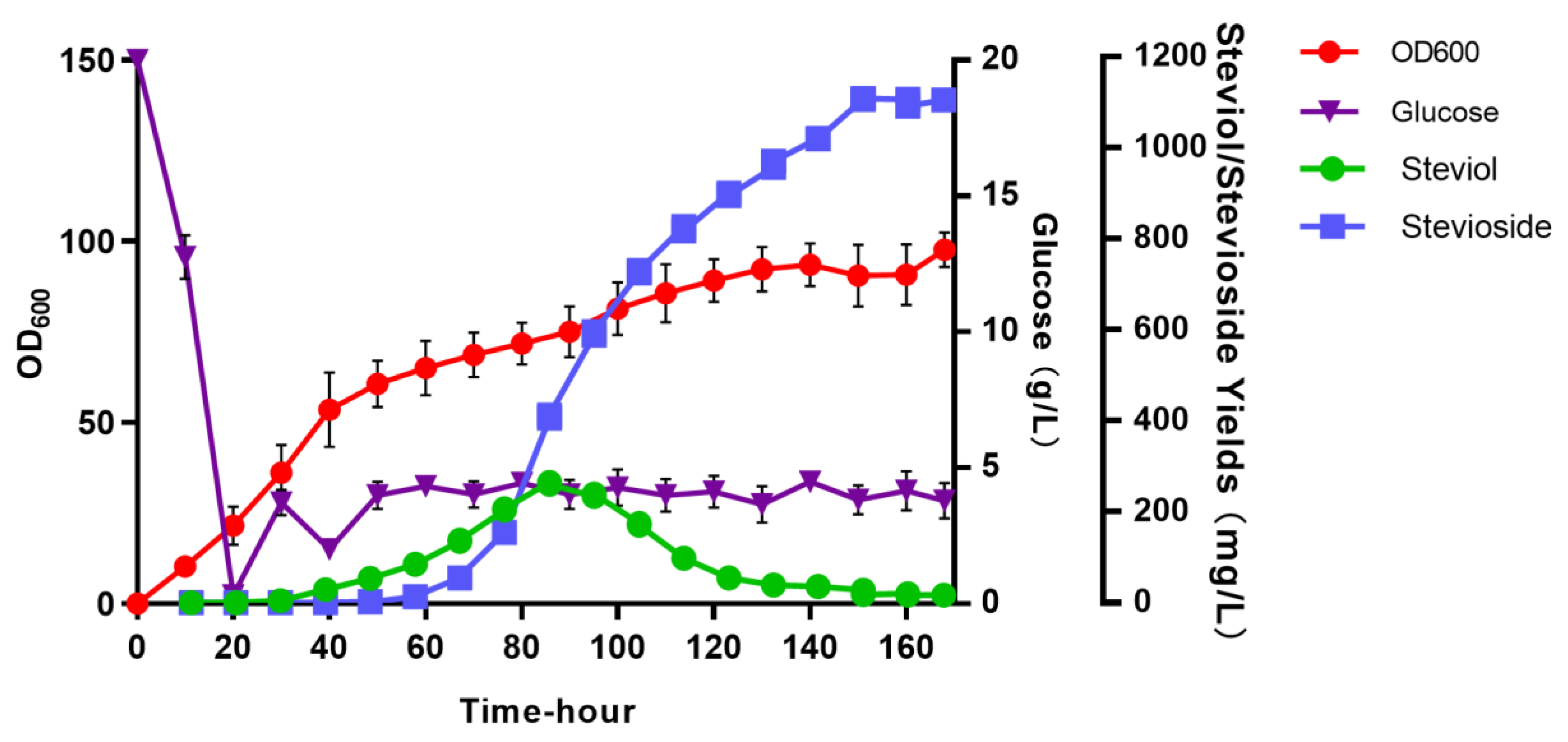
3.4. Batch Feeding Culture in a 10 L Bioreactor
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Brandle, J.E.; Telmer, P.G. Steviol glycoside biosynthesis. Phytochemistry 2007, 68, 1855–1863. [Google Scholar] [CrossRef] [PubMed]
- Geuns, J.M. Stevioside. Phytochemistry 2003, 64, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Bondarev, N.I.; Sukhanova, M.A.; Reshetnyak, O.V.; Nosov, A.M. Steviol glycoside content in different organs of Stevia rebaudiana and its dynamics during ontogeny. Biol. Plantarum. 2002, 47, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Kaul, K.; Bajpai-Gupta, S.; Kaul, V.K.; Kumar, S. A comprehensive analysis of fifteen genes of steviol glycosides biosynthesis pathway in Stevia rebaudiana (Bertoni). Gene 2012, 492, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Richman, A.S.; Gijzen, M.; Starratt, A.N.; Yang, Z.; Brandle, J.E. Diterpene synthesis in Stevia rebaudiana: Recruitment and up-regulation of key enzymes from the gibberellin biosynthetic pathway. Plant J. 1999, 19, 411–421. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Wang, Y.; Chen, L.; Yan, M.; Chen, K.; Xu, L.; Ouyang, P. Production of Rebaudioside A from Stevioside Catalyzed by the Engineered Saccharomyces cerevisiae. Appl. Biochem. Biotechnol. 2016, 178, 1586–1598. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Jin, J.; Zheng, J.; Wong, L.; Chua, N.H.; Jang, I.C. Comparative Transcriptomics Unravel Biochemical Specialization of Leaf Tissues of Stevia for Diterpenoid Production. Plant Physiol. 2015, 169, 2462–2480. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hou, K.; Qin, P.; Liu, H.; Yi, B.; Yang, W.; Wu, W. RNA-Seq for gene identification and transcript profiling of three Stevia rebaudiana genotypes. BMC Genom. 2014, 15, 571. [Google Scholar] [CrossRef]
- Ashok, K.K.Y.; Singh, S.; Dhyani, D.; Ahuja, P.S. A review on the improvement of stevia [Stevia rebaudiana (Bertoni)]. Can. J. Plant Sci. 2011, 91, 1–27. [Google Scholar] [CrossRef]
- Wang, J. Discussing the mechanism of sweetness, sweetness and bitter aftertaste of stevia glycosides. Food Ind. Technol. 2010, 31, 417–420. [Google Scholar] [CrossRef]
- Ceunen, S.; Geuns, J.M. Steviol glycosides: Chemical diversity, metabolism, and function. J. Nat. Prod. 2013, 76, 1201–1228. [Google Scholar] [CrossRef]
- Olsson, K.; Carlsen, S.; Semmler, A.; Simón, E.; Mikkelsen, M.D.; Møller, B.L. Microbial production of next-generation stevia sweeteners. Microb. Cell Fact. 2016, 15, 207. [Google Scholar] [CrossRef]
- Carothers, J.M.; Goler, J.A.; Keasling, J.D. Chemical synthesis using synthetic biology. Curr. Opin. Biotechnol. 2009, 20, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Ro, D.K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 2006, 440, 940–943. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Y.; Nielsen, J. Synthetic Biology of Yeast. Biochemistry 2019, 58, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, S.; Xiong, Z.; Wang, Y. Pathway mining-based integration of critical enzyme parts for de novo biosynthesis of steviolglycosides sweetener in Escherichia coli. Cell Res. 2016, 26, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, X.; Zhang, C.; Zhou, X.; Xu, X.; Han, L.; Lv, X.; Liu, Y.; Liu, S.; Li, J.; et al. De novo biosynthesis of rubusoside and rebaudiosides in engineered yeasts. Nat. Commun. 2022, 13, 3040. [Google Scholar] [CrossRef]
- Mao, Y.; Chen, Z.; Ren, Y.; Sun, Y.; Wang, Y. Whole-Cell Biocatalyst for Rubusoside Production in Saccharomyces cerevisiae. J. Agric. Food Chem. 2021, 69, 13155–13163. [Google Scholar] [CrossRef]
- Ye, L.D.; Lv, X.M.; Yu, H.W. Assembly of biosynthetic pathways in Saccharomyces cerevisiae using a marker recyclable integrative plasmid toolbox. Front. Chem. Sci. Eng. 2017, 11, 126–132. [Google Scholar] [CrossRef]
- Reider Apel, A.; d’Espaux, L.; Wehrs, M.; Sachs, D.; Li, R.A.; Tong, G.J.; Garber, M.; Nnadi, O.; Zhuang, W.; Hillson, N.J.; et al. A Cas9-based toolkit to program gene expression in Saccharomyces cerevisiae. Nucleic Acids Res. 2017, 45, 496–508. [Google Scholar] [CrossRef]
- Nan, W.; Zhao, F.; Zhang, C.; Ju, H.; Lu, W. Promotion of compound K production in Saccharomyces cerevisiae by glycerol. Microb. Cell Fact. 2020, 19, 41. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, Y.; Liu, M.; Qu, J.; Yao, M.; Li, B.; Ding, M.; Liu, H.; Xiao, W.; Yuan, Y. Primary and Secondary Metabolic Effects of a Key Gene Deletion (ΔYPL062W) in Metabolically Engineered Terpenoid-Producing Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2019, 85, e01990-18. [Google Scholar] [CrossRef] [PubMed]
- Amiri, P.; Shahpiri, A.; Asadollahi, M.A.; Momenbeik, F.; Partow, S. Metabolic engineering of Saccharomyces cerevisiae for linalool production. Biotechnol. Lett. 2016, 38, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Scalcinati, G.; Knuf, C.; Partow, S.; Chen, Y.; Maury, J.; Schalk, M.; Daviet, L.; Nielsen, J.; Siewers, V. Dynamic control of gene expression in Saccharomyces cerevisiae engineered for the production of plant sesquitepene α-santalene in a fed-batch mode. Metab. Eng. 2012, 14, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Zhou, J.; Tong, Y.; Su, P.; Li, X.; Liu, Y.; Liu, N.; Wu, X.; Zhang, Y.; Wang, J.; et al. Engineering chimeric diterpene synthases and isoprenoid biosynthetic pathways enables high-level production of miltiradiene in yeast. Metab. Eng. 2020, 60, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Gao, W.; Rong, Q.; Jin, G.; Chu, H.; Liu, W.; Yang, W.; Zhu, Z.; Li, G.; Zhu, G.; et al. Modular pathway engineering of diterpenoid synthases and the mevalonic acid pathway for miltiradiene production. J. Am. Chem. Soc. 2012, 134, 3234–3241. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P.; Phillips, A.L. Gibberellin metabolism: New insights revealed by the genes. Trends Plant Sci. 2000, 5, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Brown, R.; Köllner, T.G.; Fu, J.; Chen, X.; Wong, G.K.; Gershenzon, J.; Peters, R.J.; Chen, F. Origin and early evolution of the plant terpene synthase family. Proc. Natl. Acad. Sci. USA 2022, 119, e2100361119. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.C.; Woo, H.M. Biosynthesis of the Calorie-Free Sweetener Precursor ent-Kaurenoic Acid from CO2 Using Engineered Cyanobacteria. ACS Synth. Biol. 2020, 9, 2979–2985. [Google Scholar] [CrossRef]
- Li, W.; Cui, L.; Mai, J.; Shi, T.Q.; Lin, L.; Zhang, Z.G.; Ledesma-Amaro, R.; Dong, W.; Ji, X.J. Advances in Metabolic Engineering Paving the Way for the Efficient Biosynthesis of Terpenes in Yeasts. J. Agric. Food Chem. 2022, 70, 9246–9261. [Google Scholar] [CrossRef]
- Zhou, K.; Gao, Y.; Hoy, J.A.; Mann, F.M.; Honzatko, R.B.; Peters, R.J. Insights into diterpene cyclization from structure of bifunctional abietadiene synthase from Abies grandis. J. Biol. Chem. 2012, 287, 6840–6850. [Google Scholar] [CrossRef] [PubMed]
- Richman, A.; Swanson, A.; Humphrey, T.; Chapman, R.; McGarvey, B.; Pocs, R.; Brandle, J. Functional genomics uncovers three glucosyltransferases involved in the synthesis of the major sweet glucosides of Stevia rebaudiana. Plant J. 2005, 41, 56–67. [Google Scholar] [CrossRef]
- Kishore, G.M.L.; Motion, M.L.; Hicks, P.M.L.; Houghton-Larsen, J.L.; Hansen, J.L.; Halkjear Hansen, E.L.; Tavares, S.L.; Blom, C.L. Recombinant Production of Steviol. Glycosides. Patent WO2013022989A2, 14 February 2013. [Google Scholar]
- Chen, G.H.; Li, C.; Feng, X.D. Advances in UDP-Glc biosynthesis. J. Biol. 2023, 40, 95–100. [Google Scholar] [CrossRef]
- Li, X.; Yu, W.W.; Lv, X.Q. Metabolic engineering of Saccharomyces cerevisiae to synthesize β-glucan efficiently. Food Ferment. Ind. 2022, 48, 1–7. [Google Scholar] [CrossRef]
- Zhuang, Y.; Yang, G.Y.; Chen, X.; Liu, Q.; Zhang, X.; Deng, Z.; Feng, Y. Biosynthesis of plant-derived ginsenoside Rh2 in yeast via repurposing a key promiscuous microbial enzyme. Metab. Eng. 2017, 42, 25–32. [Google Scholar] [CrossRef]
- Redden, H.; Morse, N.; Alper, H.S. The synthetic biology toolbox for tuning gene expression in yeast. FEMS Yeast Res. 2015, 15, 1–10. [Google Scholar] [CrossRef]
- Wu, G.; Yan, Q.; Jones, J.A.; Tang, Y.J.; Fong, S.S.; Koffas, M.A.G. Metabolic Burden: Cornerstones in Synthetic Biology and Metabolic Engineering Applications. Trends Biotechnol. 2016, 34, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Vogt, T. Glycosyltransferases in secondary plant metabolism: Tranquilizers and stimulant controllers. Planta 2001, 213, 164–174. [Google Scholar] [CrossRef]
- Gachon, C.M.; Langlois-Meurinne, M.; Saindrenan, P. Plant secondary metabolism glycosyltransferases: The emerging functional analysis. Trends Plant Sci. 2005, 10, 542–549. [Google Scholar] [CrossRef]
- Bowles, D.; Isayenkova, J.; Lim, E.K.; Poppenberger, B. Glycosyltransferases: Managers of small molecules. Curr. Opin. Plant Biol. 2005, 8, 254–263. [Google Scholar] [CrossRef]
- Lian, J.; Mishra, S.; Zhao, H. Recent advances in metabolic engineering of Saccharomyces cerevisiae: New tools and their applications. Metab. Eng. 2018, 50, 85–108. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, W.; Liu, Y.; Ma, X.; Ma, C.; Jiang, Y.; Su, J. Rational Design for the Complete Synthesis of Stevioside in Saccharomyces cerevisiae. Microorganisms 2024, 12, 1125. https://doi.org/10.3390/microorganisms12061125
Huang W, Liu Y, Ma X, Ma C, Jiang Y, Su J. Rational Design for the Complete Synthesis of Stevioside in Saccharomyces cerevisiae. Microorganisms. 2024; 12(6):1125. https://doi.org/10.3390/microorganisms12061125
Chicago/Turabian StyleHuang, Wei, Yongheng Liu, Xiaomei Ma, Cilang Ma, Yuting Jiang, and Jianyu Su. 2024. "Rational Design for the Complete Synthesis of Stevioside in Saccharomyces cerevisiae" Microorganisms 12, no. 6: 1125. https://doi.org/10.3390/microorganisms12061125
APA StyleHuang, W., Liu, Y., Ma, X., Ma, C., Jiang, Y., & Su, J. (2024). Rational Design for the Complete Synthesis of Stevioside in Saccharomyces cerevisiae. Microorganisms, 12(6), 1125. https://doi.org/10.3390/microorganisms12061125