

Application of Cyanobacteria as Chassis Cells in Synthetic Biology


Abstract
1. Introduction
2. Optimization Strategy of Cyanobacteria Chassis Cells
2.1. Develop Cyanobacteria Chassis Organisms with Fast Growth, Strong Tolerance, and High Yield

{kind=link}
| Species | Features | Cultural Conditions (Optimum) | References |
|---|---|---|---|
| Synechococcus sp. PCC 11801 | Fast-growing (Doubling time: 2.3 h), Tolerant to high temperature, light intensities, Higher rates of photosynthesis and carbon fixation | 38 °C Carbon-free 1000 µmol photons/m2/s | [10] |
| Synechococcus sp. PCC 11802 | Fast-growing (Doubling time: 2.8 h), Tolerant to high temperature, light and carbon dioxide intensities | 38 °C 1% CO2 1000 µmol photons/m2/s | [10] |
| Synechococcus sp. PCC 7002 | Fast-growing (Doubling time: 2.27 h), Tolerant to high salt light and temperature intensities | 38 °C | [19] |
| Synechococcus sp. PCC 11901 | Fast-growing (Doubling time: 2 h), Tolerant to high temperature and light intensities, Showed the highest biomass accumulation | Mixed nutrient culture (use the modified AD7 medium) | [20] |
| Anabaena sp. PCC 7120 | Heterocystic specialized cells, High nitrogen fixation efficiency | 23 °C | [21] |
| Synechococcus elongatus PCC 7942 | Fast-growing (Doubling time: 4.9 h), Small genome, Easy to convert, wide salt tolerance | 38 °C 300 µmol photons/m2/s, Seawater culture | [22] |
| Synechocystis sp. PCC 6803 | Fast-growing (Doubling time: 6.6 h), Easy to convert, wide salt tolerance | 30 °C 300 µmol photons/m2/s | [23] |
| Synechococcus sp. NKBG 15041c | Fast-growing, high glycogen accumulation under nitrogen deficient culture | Seawater culture | [24] |
| Synechococcus elongatus UTEX 2973 | Fast-growing (Doubling time: 1.93 h), Tolerant to high temperatures and light intensities, Easy to convert, Showed the highest biomass accumulation | 38 °C Freshwater culture | [12] |
| Prochlorococcus | Small genome, Adapted to darkness, Tolerance to high temperature and nitrate | Seawater culture | [25] |
2.2. Adaptive Evolutionary Strategy
2.3. Semicontinuous Algal Culture Supported by Machine Learning and Synthesis
3. Modification of Cyanobacteria Chassis Cells for Synthetic Biology
3.1. Altered Endogenous Gene Expression
3.2. Introducing Heterologous Metabolic Pathways
3.3. Artificial Intelligence Design Genetic Elements
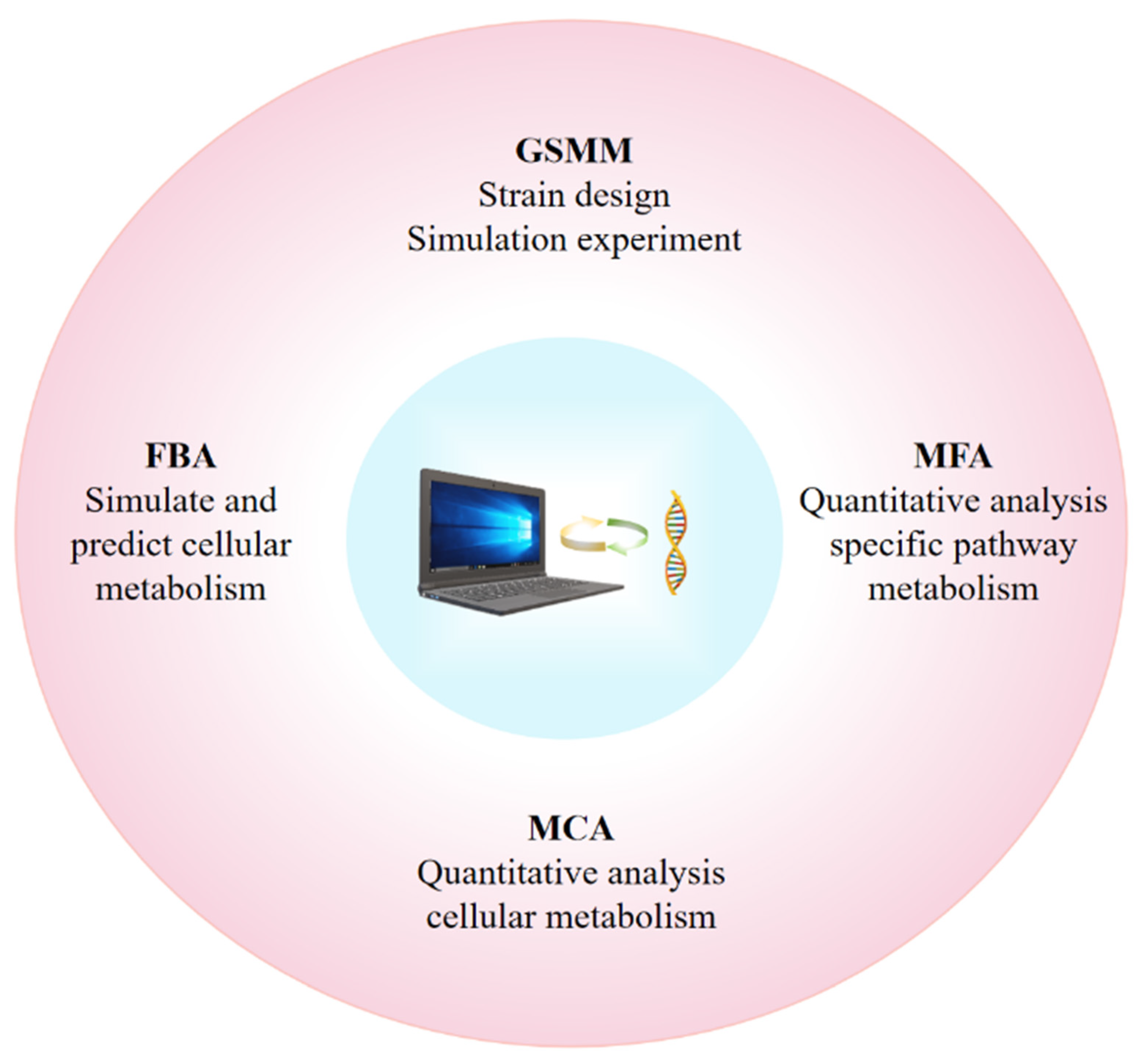
3.4. Using Artificial Intelligence and Mathematical Models to Identify and Optimize Metabolic Stuck Point
4. “Green Engineering” Built in the Chassis Cells of Cyanobacteria
4.1. Macromolecular Substance
4.1.1. PHA
4.1.2. PHB
4.2. Water-Insoluble Liquid Substance
4.2.1. Limonene
4.2.2. Biodiesel
4.2.3. FFAs
4.3. Gas
4.3.1. Ethylene
4.3.2. Hydrogen
4.4. Water-Soluble but High-Value Products
4.4.1. Ethanol
4.4.2. Sucrose
4.4.3. Lysine
4.4.4. Succinic Acid
5. Application of Modified Cyanobacteria Chassis Cells in Environmental Monitoring and Remediation
5.1. Application of Modified Cyanobacteria in Environmental Monitoring
5.2. Application of Modified Cyanobacteria in Environmental Remediation
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pandey, A.; Srivastava, S.; Kumar, S. Development and cost-benefit analysis of a novel process for biofuel production from microalgae using pre-treated high-strength fresh cheese whey wastewater. Environ. Sci. Pollut. Res. 2020, 27, 23963–23980. [Google Scholar] [CrossRef] [PubMed]
- Gowrisankar, A.; Priyanka, T.M.C.; Saha, A.; Rondoni, L.; Kamrul Hassan, M.; Banerjee, S. Greenhouse gas emissions: A rapid submerge of the world. Chaos 2022, 32, 061104. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.F.; Ling, H.; Foo, J.L.; Chang, M.W. Synthetic Biology Toolkits for Metabolic Engineering of Cyanobacteria. Biotechnol. J. 2019, 14, e1800496. [Google Scholar] [CrossRef]
- Bourgade, B.; Stensjö, K. Synthetic biology in marine cyanobacteria: Advances and challenges. Front. Microbiol. 2022, 13, 994365. [Google Scholar] [CrossRef] [PubMed]
- Ng, I.S.; Keskin, B.B.; Tan, S.I. A Critical Review of Genome Editing and Synthetic Biology Applications in Metabolic Engineering of Microalgae and Cyanobacteria. Biotechnol. J. 2020, 15, e1900228. [Google Scholar] [CrossRef] [PubMed]
- Yadav, I.; Rautela, A.; Kumar, S. Approaches in the photosynthetic production of sustainable fuels by cyanobacteria using tools of synthetic biology. World J. Microbiol. Biotechnol. 2021, 37, 201. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, D.; Sahasrabuddhe, D.; Wangikar, P.P. Cyanobacteria as cell factories: The roles of host and pathway engineering and translational research. Curr. Opin. Biotechnol. 2022, 73, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Ramey, C.J.; Barón-Sola, Á.; Aucoin, H.R.; Boyle, N.R. Genome Engineering in Cyanobacteria: Where We Are and Where We Need to Go. ACS Synth. Biol. 2015, 4, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, S.; Chen, L.; Sun, T.; Zhang, W. Fast-growing cyanobacterial chassis for synthetic biology application. Crit. Rev. Biotechnol. 2023, 44, 414–428. [Google Scholar] [CrossRef]
- Jaiswal, D.; Sengupta, A.; Sengupta, S.; Madhu, S.; Pakrasi, H.B.; Wangikar, P.P. A Novel Cyanobacterium Synechococcus elongatus PCC 11802 has Distinct Genomic and Metabolomic Characteristics Compared to its Neighbor PCC 11801. Sci. Rep. 2020, 10, 191. [Google Scholar] [CrossRef]
- Luan, G.; Zhang, S.; Lu, X. Engineering cyanobacteria chassis cells toward more efficient photosynthesis. Curr. Opin. Biotechnol. 2020, 62, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Liberton, M.; Cliften, P.F.; Head, R.D.; Jacobs, J.M.; Smith, R.D.; Koppenaal, D.W.; Brand, J.J.; Pakrasi, H.B. Synechococcus elongatus UTEX 2973, a fast growing cyanobacterial chassis for biosynthesis using light and CO2. Sci. Rep. 2015, 5, srep08132. [Google Scholar] [CrossRef] [PubMed]
- Bryant, D.A.; Ludwig, M. Transcription Profiling of the Model Cyanobacterium Synechococcus sp. Strain PCC 7002 by Next-Gen (SOLiD™) Sequencing of cDNA. Front. Microbiol. 2011, 2, 9540. [Google Scholar] [CrossRef]
- Gao, X.; Sun, T.; Pei, G.; Chen, L.; Zhang, W. Cyanobacterial chassis engineering for enhancing production of biofuels and chemicals. Appl. Microbiol. Biotechnol. 2016, 100, 3401–3413. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Sun, T.; Chen, L.; Zhang, W. Salt-Tolerant Synechococcus elongatus UTEX 2973 Obtained via Engineering of Heterologous Synthesis of Compatible Solute Glucosylglycerol. Front. Microbiol. 2021, 12, 650217. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Sun, T.; Li, S.; Xie, Y.; Song, X.; Wang, F.; Chen, L.; Zhang, W. Improved Salt Tolerance and Metabolomics Analysis of Synechococcus elongatus UTEX 2973 by Overexpressing Mrp Antiporters. Front. Bioeng. Biotechnol. 2020, 8, 500. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarzyk, D.; Anfelt, J.; Särnegrim, A.; Hudson, E.P. Overexpression of sigma factor SigB improves temperature and butanol tolerance of Synechocystis sp. PCC6803. J. Biotechnol. 2014, 182–183, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Zhang, H.; Zhou, Z.; Li, K.; Hou, G.; Xu, Q.; Chuai, W.; Zhang, C.; Han, D.; Hu, Q. Ultrahigh-cell-density heterotrophic cultivation of the unicellular green microalga Scenedesmus acuminatus and application of the cells to photoautotrophic culture enhance biomass and lipid production. Biotechnol. Bioeng. 2019, 117, 96–108. [Google Scholar] [CrossRef]
- Ruffing, A.M.; Jensen, T.J.; Strickland, L.M. Genetic tools for advancement of Synechococcus sp. PCC 7002 as a cyanobacterial chassis. Microb. Cell Factories 2016, 15, 190. [Google Scholar] [CrossRef]
- Włodarczyk, A.; Selão, T.T.; Norling, B.; Nixon, P.J. Newly discovered Synechococcus sp. PCC 11901 is a robust cyanobacterial strain for high biomass production. Commun. Biol. 2020, 3, 215. [Google Scholar] [CrossRef]
- Malatinszky, D.; Steuer, R.; Jones, P.R. A Comprehensively Curated Genome-Scale Two-Cell Model for the Heterocystous Cyanobacterium Anabaena sp. PCC 7120. Plant Physiol. 2017, 173, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Tsinoremas, N.F.; Kutach, A.K.; Strayer, C.A.; Golden, S.S. Efficient gene transfer in Synechococcus sp. strains PCC 7942 and PCC 6301 by interspecies conjugation and chromosomal recombination. J. Bacteriol. 1994, 176, 6764–6768. [Google Scholar] [CrossRef] [PubMed]
- Zang, X.; Liu, B.; Liu, S.; Arunakumara, K.K.; Zhang, X. Optimum conditions for transformation of Synechocystis sp. PCC 6803. J. Microbiol. 2007, 45, 241–245. [Google Scholar]
- Badary, A.; Takamatsu, S.; Nakajima, M.; Ferri, S.; Lindblad, P.; Sode, K. Glycogen Production in Marine Cyanobacterial Strain Synechococcus sp. NKBG 15041c. Mar. Biotechnol. 2018, 20, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.W. Prochlorococcus. Curr. Biol. 2017, 27, R447–R448. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-Y.; Teng, W.-K.; Zhao, L.; Hu, C.-X.; Zhou, Y.-K.; Han, B.-P.; Song, L.-R.; Shu, W.-S. Comparative genomics reveals insights into cyanobacterial evolution and habitat adaptation. ISME J. 2021, 15, 211–227. [Google Scholar] [CrossRef]
- Six, C.; Ratin, M.; Marie, D.; Corre, E. Marine Synechococcus picocyanobacteria: Light utilization across latitudes. Proc. Natl. Acad. Sci. USA 2021, 118, e2111300118. [Google Scholar] [CrossRef]
- Sun, H.; Luan, G.; Ma, Y.; Lou, W.; Chen, R.; Feng, D.; Zhang, S.; Sun, J.; Lu, X. Engineered hypermutation adapts cyanobacterial photosynthesis to combined high light and high temperature stress. Nat. Commun. 2023, 14, 1238. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shi, M.; Niu, X.; Zhang, X.; Gao, L.; Chen, L.; Wang, J.; Zhang, W. Metabolomic basis of laboratory evolution of butanol tolerance in photosynthetic Synechocystis sp. PCC 6803. Microb. Cell Factories 2014, 13, 151. [Google Scholar] [CrossRef]
- Srivastava, V.; Amanna, R.; Rowden, S.J.L.; Sengupta, S.; Madhu, S.; Howe, C.J.; Wangikar, P.P. Adaptive laboratory evolution of the fast-growing cyanobacterium Synechococcus elongatus PCC 11801 for improved solvent tolerance. J. Biosci. Bioeng. 2021, 131, 491–500. [Google Scholar] [CrossRef]
- Matsusako, T.; Toya, Y.; Yoshikawa, K.; Shimizu, H. Identification of alcohol stress tolerance genes of Synechocystis sp. PCC 6803 using adaptive laboratory evolution. Biotechnol. Biofuels 2017, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, J.; Kanesaki, Y.; Iwata, N.; Asakura, R.; Funamizu, K.; Tasaki, R.; Agatsuma, M.; Tahara, H.; Matsuhashi, A.; Yoshikawa, H.; et al. Genomic analysis of parallel-evolved cyanobacterium Synechocystis sp. PCC 6803 under acid stress. Photosynth. Res. 2015, 125, 243–254. [Google Scholar] [CrossRef]
- Xu, C.; Sun, T.; Li, S.; Chen, L.; Zhang, W. Adaptive laboratory evolution of cadmium tolerance in Synechocystis sp. PCC 6803. Biotechnol. Biofuels 2018, 11, 205. [Google Scholar] [CrossRef] [PubMed]
- Luan, G.; Lu, X. Tailoring cyanobacterial cell factory for improved industrial properties. Biotechnol. Adv. 2018, 36, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Decorosi, F.; Ratti, C.; De Philippis, R.; Adessi, A. Semi-continuous cultivation of EPS-producing marine cyanobacteria: A green biotechnology to remove dissolved metals obtaining metal-organic materials. New Biotechnol. 2024, 82, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Karemore, A.; Yuan, Y.; Porubsky, W.; Chance, R. Biomass and pigment production for Arthrospira platensis via semi-continuous cultivation in photobioreactors: Temperature effects. Biotechnol. Bioeng. 2020, 117, 3081–3093. [Google Scholar] [CrossRef]
- Long, B.; Fischer, B.; Zeng, Y.; Amerigian, Z.; Li, Q.; Bryant, H.; Li, M.; Dai, S.Y.; Yuan, J.S. Machine learning-informed and synthetic biology-enabled semi-continuous algal cultivation to unleash renewable fuel productivity. Nat. Commun. 2022, 13, 541. [Google Scholar] [CrossRef]
- Lin, P.-C.; Saha, R.; Zhang, F.; Pakrasi, H.B. Metabolic engineering of the pentose phosphate pathway for enhanced limonene production in the cyanobacterium Synechocystis sp. PCC 6803. Sci. Rep. 2017, 7, 17503. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.W.; Carvalho, B.J.; Jones, H.D.T.; Singh, S. The role of photo-osmotic adaptation in semi-continuous culture and lipid particle release from Dunaliella viridis. J. Appl. Phycol. 2014, 27, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, D.; Chen, H.; Wang, Q. Advances in Genetic Engineering in Improving Photosynthesis and Microalgal Productivity. Int. J. Mol. Sci. 2023, 24, 1898. [Google Scholar] [CrossRef]
- Su, H.Y.; Chou, H.H.; Chow, T.J.; Lee, T.M.; Chang, J.S.; Huang, W.L.; Chen, H.J. Improvement of outdoor culture efficiency of cyanobacteria by over-expression of stress tolerance genes and its implication as bio-refinery feedstock. Bioresour. Technol. 2017, 244 Pt 2, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Angermayr, S.A.; Gorchs Rovira, A.; Hellingwerf, K.J. Metabolic engineering of cyanobacteria for the synthesis of commodity products. Trends Biotechnol. 2015, 33, 352–361. [Google Scholar] [CrossRef]
- Chaurasia, A.K.; Apte, S.K. Overexpression of the groESL operon enhances the heat and salinity stress tolerance of the nitrogen-fixing cyanobacterium Anabaena sp. strain PCC7120. Appl. Environ. Microbiol. 2009, 75, 6008–6012. [Google Scholar] [CrossRef]
- Gonzalez-Esquer, C.R.; Vermaas, W.F. ClpB1 overproduction in Synechocystis sp. strain PCC 6803 increases tolerance to rapid heat shock. Appl. Environ. Microbiol. 2013, 79, 6220–6227. [Google Scholar] [CrossRef] [PubMed]
- Sakthivel, K.; Watanabe, T.; Nakamoto, H. A small heat-shock protein confers stress tolerance and stabilizes thylakoid membrane proteins in cyanobacteria under oxidative stress. Arch. Microbiol. 2009, 191, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Vachiranuvathin, P.; Tharasirivat, V.; Hemnusornnanon, T.; Jantaro, S. Native SodB Overexpression of Synechocystis sp. PCC 6803 Improves Cell Growth under Alcohol Stresses Whereas Its Gpx2 Overexpression Impacts on Growth Recovery from Alcohol Stressors. Appl. Biochem. Biotechnol. 2022, 194, 5748–5766. [Google Scholar] [CrossRef] [PubMed]
- Narayan, O.P.; Kumari, N.; Bhargava, P.; Rajaram, H.; Rai, L.C. A single gene all3940 (Dps) overexpression in Anabaena sp. PCC 7120 confers multiple abiotic stress tolerance via proteomic alterations. Funct. Integr. Genom. 2016, 16, 67–78. [Google Scholar] [CrossRef]
- Raghavan, P.S.; Potnis, A.A.; Gupta, S.; Gadly, T.; Kushwah, N.; Rajaram, H. Interlink between ExoD (Alr2882), exopolysaccharide synthesis and metal tolerance in Nostoc sp. strain PCC 7120: Insight into its role, paralogs and evolution. Int. J. Biol. Macromol. 2023, 242, 125014. [Google Scholar] [CrossRef]
- Sun, T.; Li, S.; Pei, G.; Chen, L.; Zhang, W. Towards understanding the mechanism of n-hexane tolerance in Synechocystis sp. PCC 6803. Chin. J. Chem. Eng. 2023, 59, 128–134. [Google Scholar] [CrossRef]
- Lou, W.; Tan, X.; Song, K.; Zhang, S.; Luan, G.; Li, C.; Lu, X. A Specific Single Nucleotide Polymorphism in the ATP Synthase Gene Significantly Improves Environmental Stress Tolerance of Synechococcus elongatus PCC 7942. Appl. Environ. Microbiol. 2018, 84, e01222-18. [Google Scholar] [CrossRef]
- Dexter, J.; Dziga, D.; Lv, J.; Zhu, J.; Strzalka, W.; Maksylewicz, A.; Maroszek, M.; Marek, S.; Fu, P. Heterologous expression of mlrA in a photoautotrophic host-Engineering cyanobacteria to degrade microcystins. Environ. Pollut. 2018, 237, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Taton, A.; Ecker, A.; Diaz, B.; Moss, N.A.; Anderson, B.; Reher, R.; Leão, T.F.; Simkovsky, R.; Dorrestein, P.C.; Gerwick, L.; et al. Heterologous Expression of Cryptomaldamide in a Cyanobacterial Host. ACS Synth. Biol. 2020, 9, 3364–3376. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo Martinez, D.; Betterle, N.; Melis, A. Phycocyanin Fusion Constructs for Heterologous Protein Expression Accumulate as Functional Heterohexameric Complexes in Cyanobacteria. ACS Synth. Biol. 2022, 11, 1152–1166. [Google Scholar] [CrossRef] [PubMed]
- Ergün, B.G.; Çalık, P. Hybrid-architectured promoter design to engineer expression in yeast. Methods Enzym. 2021, 660, 81–104. [Google Scholar] [CrossRef]
- Seo, E.; Choi, Y.-N.; Shin, Y.R.; Kim, D.; Lee, J.W. Design of synthetic promoters for cyanobacteria with generative deep-learning model. Nucleic Acids Res. 2023, 51, 7071–7082. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Yu, H.; Deng, Y.; Qiu, Y.; Li, F.; Wang, X.; He, J.; Liang, W.; Lan, Y.; Qiao, L.; et al. Pathway Evolution Through a Bottlenecking-Debottlenecking Strategy and Machine Learning-Aided Flux Balancing. Adv. Sci. 2024, 11, e2306935. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, H.; Xu, H.; Wei, L.; Liu, L.; Hu, Z.; Wang, X. Deep flanking sequence engineering for efficient promoter design using DeepSEED. Nat. Commun. 2023, 14, 6309. [Google Scholar] [CrossRef]
- Lindblad, P.; Fuente, D.; Borbe, F.; Cicchi, B.; Conejero, J.A.; Couto, N.; Čelešnik, H.; Diano, M.M.; Dolinar, M.; Esposito, S.; et al. CyanoFactory, a European consortium to develop technologies needed to advance cyanobacteria as chassis for production of chemicals and fuels. Algal Res. 2019, 41, 101510. [Google Scholar] [CrossRef]
- Babele, P.K.; Young, J.D. Applications of stable isotope-based metabolomics and fluxomics toward synthetic biology of cyanobacteria. WIREs Syst. Biol. Med. 2019, 12, e1472. [Google Scholar] [CrossRef]
- Pathania, R.; Srivastava, A.; Srivastava, S.; Shukla, P. Metabolic systems biology and multi-omics of cyanobacteria: Perspectives and future directions. Bioresour. Technol. 2022, 343, 126007. [Google Scholar] [CrossRef]
- Ye, C.; Wei, X.; Shi, T.; Sun, X.; Xu, N.; Gao, C.; Zou, W. Genome-scale metabolic network models: From first-generation to next-generation. Appl. Microbiol. Biotechnol. 2022, 106, 4907–4920. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Chen, J.; Jiao, X.; Gong, H.; Pan, D.; Liu, L.; Zhang, Y.; Tan, T. Genome-scale metabolic network models for industrial microorganisms metabolic engineering: Current advances and future prospects. Biotechnol. Adv. 2024, 72, 108319. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Paz, S.; Schmitz, J.; Martins dos Santos, V.A.P.; Suarez-Diez, M. Enzyme-constrained models predict the dynamics of Saccharomyces cerevisiae growth in continuous, batch and fed-batch bioreactors. Microb. Biotechnol. 2022, 15, 1434–1445. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.D.; Thiele, I.; Palsson, B. What is flux balance analysis? Nat. Biotechnol. 2010, 28, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Pakrasi, H.B.; Moon, T.S.; Maranas, C.D.; Immethun, C.M.; Saha, R.; Berla, B.M. Synthetic biology of cyanobacteria: Unique challenges and opportunities. Front. Microbiol. 2013, 4, 246. [Google Scholar] [CrossRef]
- Antoniewicz, M.R. A guide to metabolic flux analysis in metabolic engineering: Methods, tools and applications. Metab. Eng. 2021, 63, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Cheah, Y.E.; Xu, Y.; Sacco, S.A.; Babele, P.K.; Zheng, A.O.; Johnson, C.H.; Young, J.D. Systematic identification and elimination of flux bottlenecks in the aldehyde production pathway of Synechococcus elongatus PCC 7942. Metab. Eng. 2020, 60, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Angermayr, S.A.; Hellingwerf, K.J. On the Use of Metabolic Control Analysis in the Optimization of Cyanobacterial Biosolar Cell Factories. J. Phys. Chem. B 2013, 117, 11169–11175. [Google Scholar] [CrossRef]
- Wang, X.; Liu, W.; Xin, C.; Zheng, Y.; Cheng, Y.; Sun, S.; Li, R.; Zhu, X.-G.; Dai, S.Y.; Rentzepis, P.M.; et al. Enhanced limonene production in cyanobacteria reveals photosynthesis limitations. Proc. Natl. Acad. Sci. USA 2016, 113, 14225–14230. [Google Scholar] [CrossRef]
- Nishiguchi, H.; Hiasa, N.; Uebayashi, K.; Liao, J.; Shimizu, H.; Matsuda, F. Transomics data-driven, ensemble kinetic modeling for system-level understanding and engineering of the cyanobacteria central metabolism. Metab. Eng. 2019, 52, 273–283. [Google Scholar] [CrossRef]
- Sengupta, S.; Jaiswal, D.; Sengupta, A.; Shah, S.; Gadagkar, S.; Wangikar, P.P. Metabolic engineering of a fast-growing cyanobacterium Synechococcus elongatus PCC 11801 for photoautotrophic production of succinic acid. Biotechnol. Biofuels 2020, 13, 89. [Google Scholar] [CrossRef]
- Santos-Merino, M.; Singh, A.K.; Ducat, D.C. New Applications of Synthetic Biology Tools for Cyanobacterial Metabolic Engineering. Front. Bioeng. Biotechnol. 2019, 7, 33. [Google Scholar] [CrossRef]
- Shabestary, K.; Hudson, E.P. Computational metabolic engineering strategies for growth-coupled biofuel production by Synechocystis. Metab. Eng. Commun. 2016, 3, 216–226. [Google Scholar] [CrossRef]
- Vikromvarasiri, N.; Noda, S.; Shirai, T.; Kondo, A. Investigation of two metabolic engineering approaches for (R,R)-2,3-butanediol production from glycerol in Bacillus subtilis. J. Biol. Eng. 2023, 17, 3. [Google Scholar] [CrossRef]
- Vongsangnak, W.; Figueiredo, L.F.; Förster, J.; Weber, T.; Thykaer, J.; Stegmann, E.; Wohlleben, W.; Nielsen, J. Genome-scale metabolic representation of Amycolatopsis balhimycina. Biotechnol. Bioeng. 2012, 109, 1798–1807. [Google Scholar] [CrossRef]
- Tan, C.; Xu, P.; Tao, F. Carbon-negative synthetic biology: Challenges and emerging trends of cyanobacterial technology. Trends Biotechnol. 2022, 40, 1488–1502. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Pugh, S.; Nielsen, D.R.; Zhang, W.; Meldrum, D.R. Engineering cyanobacteria for photosynthetic production of 3-hydroxybutyrate directly from CO2. Metab. Eng. 2013, 16, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Roh, H.; Lee, J.S.; Choi, H.I.; Sung, Y.J.; Choi, S.Y.; Woo, H.M.; Sim, S.J. Improved CO2-derived polyhydroxybutyrate (PHB) production by engineering fast-growing cyanobacterium Synechococcus elongatus UTEX 2973 for potential utilization of flue gas. Bioresour. Technol. 2021, 327, 124789. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Tao, F.; Xu, P. Direct carbon capture for the production of high-performance biodegradable plastics by cyanobacterial cell factories. Green. Chem. 2022, 24, 4470–4483. [Google Scholar] [CrossRef]
- Choi, S.Y.; Wang, J.-Y.; Kwak, H.S.; Lee, S.-M.; Um, Y.; Kim, Y.; Sim, S.J.; Choi, J.-I.; Woo, H.M. Improvement of Squalene Production from CO2 in Synechococcus elongatus PCC 7942 by Metabolic Engineering and Scalable Production in a Photobioreactor. ACS Synth. Biol. 2017, 6, 1289–1295. [Google Scholar] [CrossRef]
- Davies, F.K.; Work, V.H.; Beliaev, A.S.; Posewitz, M.C. Engineering Limonene and Bisabolene Production in Wild Type and a Glycogen-Deficient Mutant of Synechococcus sp. PCC 7002. Front. Bioeng. Biotechnol. 2014, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-C.; Zhang, F.; Pakrasi, H.B. Enhanced limonene production in a fast-growing cyanobacterium through combinatorial metabolic engineering. Metab. Eng. Commun. 2021, 12, e00164. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shen, C.R.; Liao, J.C. Isobutanol production as an alternative metabolic sink to rescue the growth deficiency of the glycogen mutant of Synechococcus elongatus PCC 7942. Photosynth. Res. 2014, 120, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Pade, N.; Erdmann, S.; Enke, H.; Dethloff, F.; Dühring, U.; Georg, J.; Wambutt, J.; Kopka, J.; Hess, W.R.; Zimmermann, R.; et al. Insights into isoprene production using the cyanobacterium Synechocystis sp. PCC 6803. Biotechnol. Biofuels 2016, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Diao, J.; Song, X.; Zhang, L.; Cui, J.; Chen, L.; Zhang, W. Tailoring cyanobacteria as a new platform for highly efficient synthesis of astaxanthin. Metab. Eng. 2020, 61, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Mo, H.; Xie, X.; Zhu, T.; Lu, X. Effects of global transcription factor NtcA on photosynthetic production of ethylene in recombinant Synechocystis sp. PCC 6803. Biotechnol. Biofuels 2017, 10, 145. [Google Scholar] [CrossRef]
- Toepel, J.; Karande, R.; Bühler, B.; Bühler, K.; Schmid, A. Photosynthesis driven continuous hydrogen production by diazotrophic cyanobacteria in high cell density capillary photobiofilm reactors. Bioresour. Technol. 2023, 373, 128703. [Google Scholar] [CrossRef] [PubMed]
- Shimada, N.; Okuda, Y.; Maeda, K.; Umeno, D.; Takaichi, S.; Ikeuchi, M. Astaxanthin production in a model cyanobacterium Synechocystis sp. PCC 6803. J. Gen. Appl. Microbiol. 2020, 66, 116–120. [Google Scholar] [CrossRef]
- Kanno, M.; Carroll, A.L.; Atsumi, S. Global metabolic rewiring for improved CO2 fixation and chemical production in cyanobacteria. Nat. Commun. 2017, 8, 14724. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Tan, X.; Liang, Y.; Lu, X. The potential of Synechococcus elongatus UTEX 2973 for sugar feedstock production. Appl. Microbiol. Biotechnol. 2016, 100, 7865–7875. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, T.; Gao, X.; Shi, M.; Wu, L.; Chen, L.; Zhang, W. Biosynthesis of platform chemical 3-hydroxypropionic acid (3-HP) directly from CO2 in cyanobacterium Synechocystis sp. PCC 6803. Metab. Eng. 2016, 34, 60–70. [Google Scholar] [CrossRef]
- Dookeran, Z.A.; Nielsen, D.R. Systematic Engineering of Synechococcus elongatus UTEX 2973 for Photosynthetic Production of l-Lysine, Cadaverine, and Glutarate. ACS Synth. Biol. 2021, 10, 3561–3575. [Google Scholar] [CrossRef]
- Knoot, C.J.; Khatri, Y.; Hohlman, R.M.; Sherman, D.H.; Pakrasi, H.B. Engineered Production of Hapalindole Alkaloids in the Cyanobacterium Synechococcus sp. UTEX 2973. ACS Synth. Biol. 2019, 8, 1941–1951. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Son, J.; Sim, S.J.; Woo, H.M. Metabolic rewiring of synthetic pyruvate dehydrogenase bypasses for acetone production in cyanobacteria. Plant Biotechnol. J. 2020, 18, 1860–1868. [Google Scholar] [CrossRef]
- Liu, X.; Xie, H.; Roussou, S.; Lindblad, P. Current advances in engineering cyanobacteria and their applications for photosynthetic butanol production. Curr. Opin. Biotechnol. 2022, 73, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, Y.; Matsuo, S.; Hamada, H.; Matsuda, F.; Hanai, T. Metabolic engineering of Synechococcus elongatus PCC 7942 for improvement of 1,3-propanediol and glycerol production based on in silico simulation of metabolic flux distribution. Microb. Cell Factories 2017, 16, 212. [Google Scholar] [CrossRef]
- Tan, X.; Du, W.; Lu, X. Photosynthetic and extracellular production of glucosylglycerol by genetically engineered and gel-encapsulated cyanobacteria. Appl. Microbiol. Biotechnol. 2014, 99, 2147–2154. [Google Scholar] [CrossRef]
- Hasunuma, T.; Matsuda, M.; Kato, Y.; Vavricka, C.J.; Kondo, A. Temperature enhanced succinate production concurrent with increased central metabolism turnover in the cyanobacterium Synechocystis sp. PCC 6803. Metab. Eng. 2018, 48, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; You, L.; Liu, D.; Hollinshead, W.; Tang, Y.; Zhang, F. Development of Synechocystis sp. PCC 6803 as a Phototrophic Cell Factory. Mar. Drugs 2013, 11, 2894–2916. [Google Scholar] [CrossRef]
- Jurko, L.; Makuc, D.; Štern, A.; Plavec, J.; Žegura, B.; Bošković, P.; Kargl, R. Cytotoxicity and Antibacterial Efficacy of Betaine- and Choline-Substituted Polymers. ACS Appl. Polym. Mater. 2023, 5, 5270–5279. [Google Scholar] [CrossRef]
- Luengo, J.M.; García, B.; Sandoval, A.; Naharro, G.; Olivera, E.a.R. Bioplastics from microorganisms. Curr. Opin. Microbiol. 2003, 6, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Tyo, K.E.J.; Jin, Y.S.; Espinoza, F.A.; Stephanopoulos, G. Identification of gene disruptions for increased poly-3-hydroxybutyrate accumulation in Synechocystis PCC 6803. Biotechnol. Prog. 2009, 25, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Shinde, S.; Singapuri, S.; Jiang, Z.; Long, B.; Wilcox, D.; Klatt, C.; Jones, J.A.; Yuan, J.S.; Wang, X. Thermodynamics contributes to high limonene productivity in cyanobacteria. Metab. Eng. Commun. 2022, 14, e00193. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, J.; Zhang, W.; Meldrum, D.R. Application of synthetic biology in cyanobacteria and algae. Front. Microbiol. 2012, 3, 31453. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Fallon, S.; Sheng, J.; Curtiss, R., III. CO2-limitation-inducible Green Recovery of fatty acids from cyanobacterial biomass. Proc. Natl. Acad. Sci. USA 2011, 108, 6905–6908. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sheng, J.; Curtiss, R., III. Fatty acid production in genetically modified cyanobacteria. Proc. Natl. Acad. Sci. USA 2011, 108, 6899–6904. [Google Scholar] [CrossRef]
- Eungrasamee, K.; Lindblad, P.; Jantaro, S. Enhanced productivity of extracellular free fatty acids by gene disruptions of acyl-ACP synthetase and S-layer protein in Synechocystis sp. PCC 6803. Biotechnol. Biofuels Bioprod. 2022, 15, 99. [Google Scholar] [CrossRef]
- Kahn, A.; Oliveira, P.; Cuau, M.; Leão, P.N. Incorporation, fate, and turnover of free fatty acids in cyanobacteria. FEMS Microbiol. Rev. 2023, 47, fuad015. [Google Scholar] [CrossRef]
- Du, Y.D.; Gu, X.B.; Wang, J.W.; Niu, W.Q. Yield and gas exchange of greenhouse tomato at different nitrogen levels under aerated irrigation. Sci. Total Environ. 2019, 668, 1156–1164. [Google Scholar] [CrossRef]
- Carbonell, V.; Vuorio, E.; Aro, E.-M.; Kallio, P. Enhanced stable production of ethylene in photosynthetic cyanobacterium Synechococcus elongatus PCC 7942. World J. Microbiol. Biotechnol. 2019, 35, 77. [Google Scholar] [CrossRef]
- Durall, C.; Lindberg, P.; Yu, J.; Lindblad, P. Increased ethylene production by overexpressing phosphoenolpyruvate carboxylase in the cyanobacterium Synechocystis PCC 6803. Biotechnol. Biofuels 2020, 13, 16. [Google Scholar] [CrossRef] [PubMed]
- Tamagnini, P.; Axelsson, R.; Lindberg, P.; Oxelfelt, F.; Wünschiers, R.; Lindblad, P. Hydrogenases and hydrogen metabolism of cyanobacteria. Microbiol. Mol. Biol. Rev. 2002, 66, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Bothe, H.; Schmitz, O.; Yates, M.G.; Newton, W.E. Nitrogen fixation and hydrogen metabolism in cyanobacteria. Microbiol. Mol. Biol. Rev. 2010, 74, 529–551. [Google Scholar] [CrossRef] [PubMed]
- Kruse, O.; Rupprecht, J.; Bader, K.-P.; Thomas-Hall, S.; Schenk, P.M.; Finazzi, G.; Hankamer, B. Improved Photobiological H2 Production in Engineered Green Algal Cells. J. Biol. Chem. 2005, 280, 34170–34177. [Google Scholar] [CrossRef] [PubMed]
- Antal, T.K.; Lindblad, P. Production of H2 by sulphur-deprived cells of the unicellular cyanobacteria Gloeocapsa alpicola and Synechocystis sp. PCC 6803 during dark incubation with methane or at various extracellular pH. J. Appl. Microbiol. 2005, 98, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Sheremetieva, M.E.; Troshina, O.Y.; Serebryakova, L.T.; Lindblad, P. Identification of hox genes and analysis of their transcription in the unicellular cyanobacterium Gloeocapsa alpicola CALU 743 growing under nitrate-limiting conditions. FEMS Microbiol. Lett. 2002, 214, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Serebryakova, L.T.; Tsygankov, A.A. Two-stage system for hydrogen production by immobilized cyanobacterium Gloeocapsa alpicola CALU 743. Biotechnol. Prog. 2007, 23, 1106–1110. [Google Scholar] [CrossRef] [PubMed]
- Al-Haj, L.; Lui, Y.; Abed, R.; Gomaa, M.; Purton, S. Cyanobacteria as Chassis for Industrial Biotechnology: Progress and Prospects. Life 2016, 6, 42. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Tao, F.; Wang, Y.; Yao, F.; Xu, P. A photoautotrophic platform for the sustainable production of valuable plant natural products from CO2. Green Chem. 2016, 18, 3537–3548. [Google Scholar] [CrossRef]
- Li, C.; Zheng, J.; Wu, Y.; Wang, X.; Shao, H.; Yan, D. Light-Driven Synthetic Biology: Progress in Research and Industrialization of Cyanobacterial Cell Factory. Life 2022, 12, 1537. [Google Scholar] [CrossRef]
- Tsygankov, A.A.; Fedorov, A.S.; Kosourov, S.N.; Rao, K.K. Hydrogen production by cyanobacteria in an automated outdoor photobioreactor under aerobic conditions. Biotechnol. Bioeng. 2002, 80, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Yan, C.; Lu, F.; Hu, Q.; Hu, Z. Growth kinetics of 1–2 mm and 3–4 mm colonies of Nostoc sphaeroides (Cyanophyta) in outdoor culture. Biotechnol. Lett. 2008, 30, 1741–1746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, L.; Wang, J.; Chen, Y.; Gao, X.; Zhang, Z.; Liu, T. Attached cultivation for improving the biomass productivity of Spirulina platensis. Bioresour. Technol. 2015, 181, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Jordan, A.; Chandler, J.; MacCready, J.S.; Huang, J.; Osteryoung, K.W.; Ducat, D.C.; Pettinari, M.J. Engineering Cyanobacterial Cell Morphology for Enhanced Recovery and Processing of Biomass. Appl. Environ. Microbiol. 2017, 83, e00053-17. [Google Scholar] [CrossRef] [PubMed]
- Chaves, J.E.; Melis, A. Engineering isoprene synthesis in cyanobacteria. FEBS Lett. 2018, 592, 2059–2069. [Google Scholar] [CrossRef] [PubMed]
- Dexter, J.; Fu, P. Metabolic engineering of cyanobacteria for ethanol production. Energy Environ. Sci. 2009, 2, 857–864. [Google Scholar] [CrossRef]
- Gao, Z.; Zhao, H.; Li, Z.; Tan, X.; Lu, X. Photosynthetic production of ethanol from carbon dioxide in genetically engineered cyanobacteria. Energy Environ. Sci. 2012, 5, 9857–9865. [Google Scholar] [CrossRef]
- Ducat, D.C.; Avelar-Rivas, J.A.; Way, J.C.; Silver, P.A. Rerouting Carbon Flux to Enhance Photosynthetic Productivity. Appl. Environ. Microbiol. 2012, 78, 2660–2668. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-C.; Zhang, F.; Pakrasi, H.B. Enhanced production of sucrose in the fast-growing cyanobacterium Synechococcus elongatus UTEX 2973. Sci. Rep. 2020, 10, 390. [Google Scholar] [CrossRef]
- Gomaa, M.A.; Al-Haj, L.; Abed, R.M.M. Metabolic engineering of Cyanobacteria and microalgae for enhanced production of biofuels and high-value products. J. Appl. Microbiol. 2016, 121, 919–931. [Google Scholar] [CrossRef]
- Jing, F.; Chen, X.; Yang, Z.; Guo, B. Heavy metals status, transport mechanisms, sources, and factors affecting their mobility in Chinese agricultural soils. Environ. Earth Sci. 2018, 77, 104. [Google Scholar] [CrossRef]
- Zahra, Z.; Choo, D.H.; Lee, H.; Parveen, A. Cyanobacteria: Review of Current Potentials and Applications. Environments 2020, 7, 13. [Google Scholar] [CrossRef]
- Subashchandrabose, S.R.; Ramakrishnan, B.; Megharaj, M.; Venkateswarlu, K.; Naidu, R. Mixotrophic cyanobacteria and microalgae as distinctive biological agents for organic pollutant degradation. Environ. Int. 2013, 51, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Hedman, J.E.; Rüdel, H.; Gercken, J.; Bergek, S.; Strand, J.; Quack, M.; Appelberg, M.; Förlin, L.; Tuvikene, A.; Bignert, A. Eelpout (Zoarces viviparus) in marine environmental monitoring. Mar. Pollut. Bull. 2011, 62, 2015–2029. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, I.; Fulk, E.M.; Kalvapalle, P.; Silberg, J.J.; Masiello, C.A.; Stadler, L.B. Translating New Synthetic Biology Advances for Biosensing into the Earth and Environmental Sciences. Front. Microbiol. 2020, 11, 618373. [Google Scholar] [CrossRef] [PubMed]
- Gavrilaș, S.; Ursachi, C.; Perța-Crișan, S.; Munteanu, F.D. Recent Trends in Biosensors for Environmental Quality Monitoring. Sensors 2022, 22, 1513. [Google Scholar] [CrossRef]
- Markhard, A.L.; McCoy, J.G.; To, T.L.; Mootha, V.K. A genetically encoded system for oxygen generation in living cells. Proc. Natl. Acad. Sci. USA 2022, 119, e2207955119. [Google Scholar] [CrossRef]
- Zheng, G.; Zhao, L.; Yuan, D.; Li, J.; Yang, G.; Song, D.; Miao, H.; Shu, L.; Mo, X.; Xu, X.; et al. A genetically encoded fluorescent biosensor for monitoring ATP in living cells with heterobifunctional aptamers. Biosens. Bioelectron. 2022, 198, 113827. [Google Scholar] [CrossRef]
- Tantama, M.; Hung, Y.P.; Yellen, G. Imaging intracellular pH in live cells with a genetically encoded red fluorescent protein sensor. J. Am. Chem. Soc. 2011, 133, 10034–10037. [Google Scholar] [CrossRef]
- Wang, F.; Gao, Y.; Yang, G. Recent advances in synthetic biology of cyanobacteria for improved chemicals production. Bioengineered 2020, 11, 1208–1220. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Z.; Chai, L.; Wang, Y.; Liu, Y.; Xiao, R. Bioleaching remediation of heavy metal-contaminated soils using Burkholderia sp. Z-90. J. Hazard. Mater. 2016, 301, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Peca, L.; Kós, P.B.; Máté, Z.; Farsang, A.; Vass, I. Construction of bioluminescent cyanobacterial reporter strains for detection of nickel, cobalt and zinc. FEMS Microbiol. Lett. 2008, 289, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Wong, L.S.; Lee, Y.H.; Surif, S. Performance of a Cyanobacteria Whole Cell-Based Fluorescence Biosensor for Heavy Metal and Pesticide Detection. Sensors 2013, 13, 6394–6404. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Rasheed, T.; Sosa-Hernández, J.E.; Raza, A.; Nabeel, F.; Iqbal, H.M.N. Biosorption: An Interplay between Marine Algae and Potentially Toxic Elements—A Review. Mar. Drugs 2018, 16, 65. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.Y.; Howe, C.J.; Porter, A.J.; Glover, L.A. Novel cyanobacterial biosensor for detection of herbicides. Appl. Environ. Microbiol. 2002, 68, 5026–5033. [Google Scholar] [CrossRef] [PubMed]
- Antonacci, A.; Scognamiglio, V. Biotechnological Advances in the Design of Algae-Based Biosensors. Trends Biotechnol. 2020, 38, 334–347. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Liu, M.; Wang, S.; Zhang, J.; Wu, H. Multi-Phase Environmental Impact Assessment of Marine Ecological Restoration Project Based on DPSIR-Cloud Model. Int. J. Environ. Res. Public. Health 2022, 19, 13295. [Google Scholar] [CrossRef]
- Ojuederie, O.B.; Babalola, O.O. Microbial and Plant-Assisted Bioremediation of Heavy Metal Polluted Environments: A Review. Int. J. Environ. Res. Public. Health 2017, 14, 1504. [Google Scholar] [CrossRef]
- de Lorenzo, V.; Prather, K.L.; Chen, G.Q.; O’Day, E.; von Kameke, C.; Oyarzún, D.A.; Hosta-Rigau, L.; Alsafar, H.; Cao, C.; Ji, W.; et al. The power of synthetic biology for bioproduction, remediation and pollution control: The UN’s Sustainable Development Goals will inevitably require the application of molecular biology and biotechnology on a global scale. EMBO Rep. 2018, 19, e45658. [Google Scholar] [CrossRef]
- Salama, E.-S.; Kurade, M.B.; Abou-Shanab, R.A.I.; El-Dalatony, M.M.; Yang, I.-S.; Min, B.; Jeon, B.-H. Recent progress in microalgal biomass production coupled with wastewater treatment for biofuel generation. Renew. Sustain. Energy Rev. 2017, 79, 1189–1211. [Google Scholar] [CrossRef]
- Pinto, F.L.; Thapper, A.; Sontheim, W.; Lindblad, P. Analysis of current and alternative phenol based RNA extraction methodologies for cyanobacteria. BMC Mol. Biol. 2009, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Touliabah, H.E.; El-Sheekh, M.M.; Ismail, M.M.; El-Kassas, H. A Review of Microalgae- and Cyanobacteria-Based Biodegradation of Organic Pollutants. Molecules 2022, 27, 1141. [Google Scholar] [CrossRef] [PubMed]
- Kuritz, T.; Wolk, C.P. Use of filamentous cyanobacteria for biodegradation of organic pollutants. Appl. Environ. Microbiol. 1995, 61, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Erbe, J.L.; Taylor, K.B.; Hall, L.M. Expression of mouse metallothionein in the cyanobacterium Synechococcus PCC7942. J. Ind. Microbiol. 1996, 17, 41–46. [Google Scholar] [CrossRef]
| Strain | Strategy | Genes | Objective | References |
|---|---|---|---|---|
| Synechocystis sp. PCC 6803 Synechococcus elongatus PCC 7942 | Overexpression | The shikimate kinase gene | Improved high light and high temperature tolerances | [28] |
| Anabaena sp. PCC 7120 | Overexpression | groESL | Improved high salt and high temperature tolerances | [43] |
| Synechocystis sp. PCC 6803 | Overexpression | clpB1, dnaK2 | Heat tolerance | [44] |
| Synechococcus elongatus PCC 7942 Synechocystis sp. PCC 6803 | Overexpression | hspA | Improved high light, high temperature, high salt, and oxidative tolerances | [41,45] |
| Synechocystis sp. PCC 6803 | Overexpression | sigB | Improved high temperature and butanol tolerances | [17] |
| Synechocystis sp. PCC 6803 | Overexpression | sodB | Improved alcohol tolerance | [46] |
| Anabaena sp. PCC 7120 | Overexpression | all3940 | Improved heavy metals, UV, salts, and temperatures tolerances | [47] |
| Anabaena sp. PCC 7120 | Overexpression | alr2882 | Improved metal tolerance | [48] |
| Anabaena sp. PCC 7120 | Overexpression | all3940 | Improved salinity, heat, heavy metals, pesticide, and nutrient starvation tolerances | [47] |
| Anabaena sp. PCC 7120 | Overexpression | groESL | Improved heat and salinity tolerances | [43] |
| Synechocystis sp. PCC 6803 | Knockout | slr0724 | Improved n-hexane tolerance | [49] |
| Synechococcus elongatus PCC 7942 | Mutation | atpA | Improved stress tolerance | [50] |
| State | Product | Cyanobacterial Chassis | Titer/Culture Time (Recombinant) | References |
|---|---|---|---|---|
| Macromolecular substance | PHA | Synechocystis sp. PCC 6803 | 533 mg/L | [77] |
| PHB | Synechococcus elongatus UTEX 2973 | 420 mg/L (10 d) | [78] | |
| PLA | Synechococcus elongatus PCC 7942 | 108 mg/L | [79] | |
| Water insoluble liquid substance | Squalene | Synechococcus elongatus UTEX 7942 | 79.2 mg/g dry cell weight (DCW) | [80] |
| α-Farnesene | Synechococcus sp. PCC 7002 | 0.6 mg/L (4 d) | [81] | |
| Limonene | Synechococcus elongatus UTEX 2973 | 16.4 mg/L (2 d) | [82] | |
| FFA | Synechocystis sp. PCC 11901 | 1.54 g/L (5 d) | [20] | |
| Isobutanol | Synechococcus elongatus PCC 7942 | 550 mg/L (8 d) | [83] | |
| Isoprene | Synechocystis sp. PCC 6803 | 1.26 g/L (21 d) | [84] | |
| Astaxanthin | Synechocystis sp. PCC 6803 | 29.6 mg/g (DCW) | [85] | |
| Gas | Ethylene | Synechocystis sp. PCC 6803 | 2463 μL (L/h/OD) | [86] |
| Hydrogen | Nostoc punctiforme ATCC 29133 | 300 µmol/L/h | [87] | |
| Water Soluble but High-Value Products | Astaxanthin | Synechocystis sp. PCC 6803 | 1.089 µg/mL/OD730 | [88] |
| 2,3-butanediol | Synechococcus elongatus PCC 7942 | 12.6 g/L | [89] | |
| Sucrose | Synechococcus elongatus UTEX 2973 | 8.7 g/L | [90] | |
| Ethanol | Synechocystis sp. PCC 6803 | 5.5 g/L (26 d) | [14] | |
| 3-hydroxypropionic acid | Synechocystis sp. PCC 6803 | 837.18 mg/L (6 d) | [91] | |
| L-Lysine | Synechococcus elongatus UTEX 2973 | 556 mg/L (5 d) | [92] | |
| Glutarate | Synechococcus elongatus UTEX 2973 | 67.5 mg/L (4 d) | [92] | |
| Cadaverine | Synechococcus elongatus UTEX 2973 | 55.3 mg/L (4 d) | [92] | |
| Alkaloids | Synechococcus elongatus UTEX 2973 | 0.75–3 mg/L (30 h) | [93] | |
| Acetone | Synechococcus elongatus PCC 7942 | 0.41 g/L/d | [94] | |
| 1-Butanol | Synechococcus elongatus PCC 7942 | 600 mg/L/d | [95] | |
| 1,3-Propanediol | Synechococcus elongatus PCC 7942 | 0.338 g/L (20 d) | [96] | |
| Glycerin glucoside (GG) | Synechocystis sp. PCC 6803 | 1.64 g/L (32 d) | [97] | |
| Succinic acid | Synechocystis sp. PCC 6803 | 1.8 g/L (3 d) | [98] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Tang, K.; Hu, J. Application of Cyanobacteria as Chassis Cells in Synthetic Biology. Microorganisms 2024, 12, 1375. https://doi.org/10.3390/microorganisms12071375
Liu X, Tang K, Hu J. Application of Cyanobacteria as Chassis Cells in Synthetic Biology. Microorganisms. 2024; 12(7):1375. https://doi.org/10.3390/microorganisms12071375
Chicago/Turabian StyleLiu, Xueli, Kaixin Tang, and Jinlu Hu. 2024. "Application of Cyanobacteria as Chassis Cells in Synthetic Biology" Microorganisms 12, no. 7: 1375. https://doi.org/10.3390/microorganisms12071375
APA StyleLiu, X., Tang, K., & Hu, J. (2024). Application of Cyanobacteria as Chassis Cells in Synthetic Biology. Microorganisms, 12(7), 1375. https://doi.org/10.3390/microorganisms12071375