
Fecal Microbiota Characteristics in Constipation-Predominant and Mixed-Type Irritable Bowel Syndrome
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Principles
2.2. Objects of Study
2.3. Ion Torrent PGM Sequencing
2.4. Bioinformatics and Statistical Analysis
3. Results
3.1. Gut Microbiome Composition of the Studied Groups, Defined by Phylum
3.2. Microbiome Composition of the Studied Groups, Defined by Species
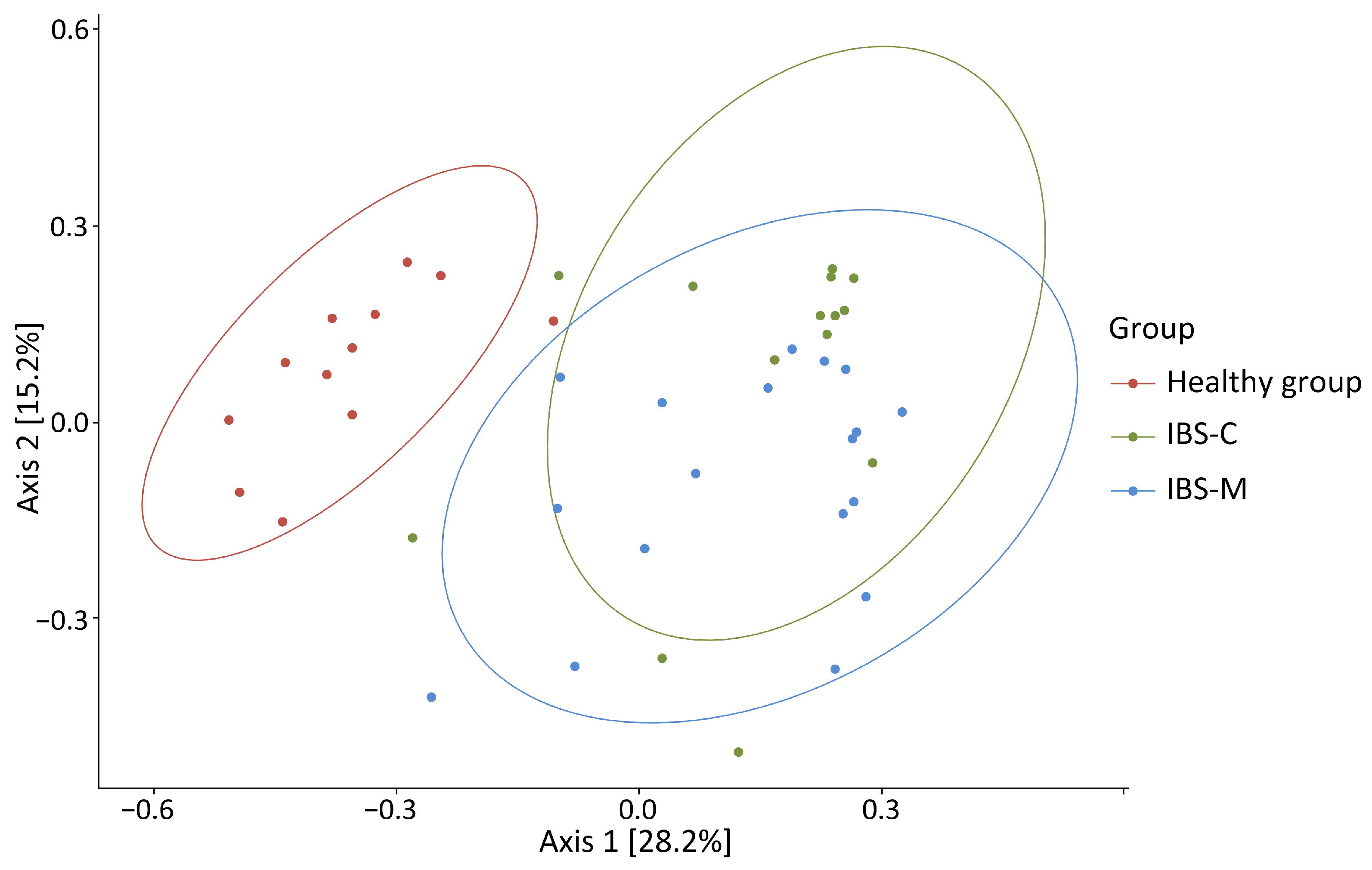
3.3. Alpha and Beta Diversity
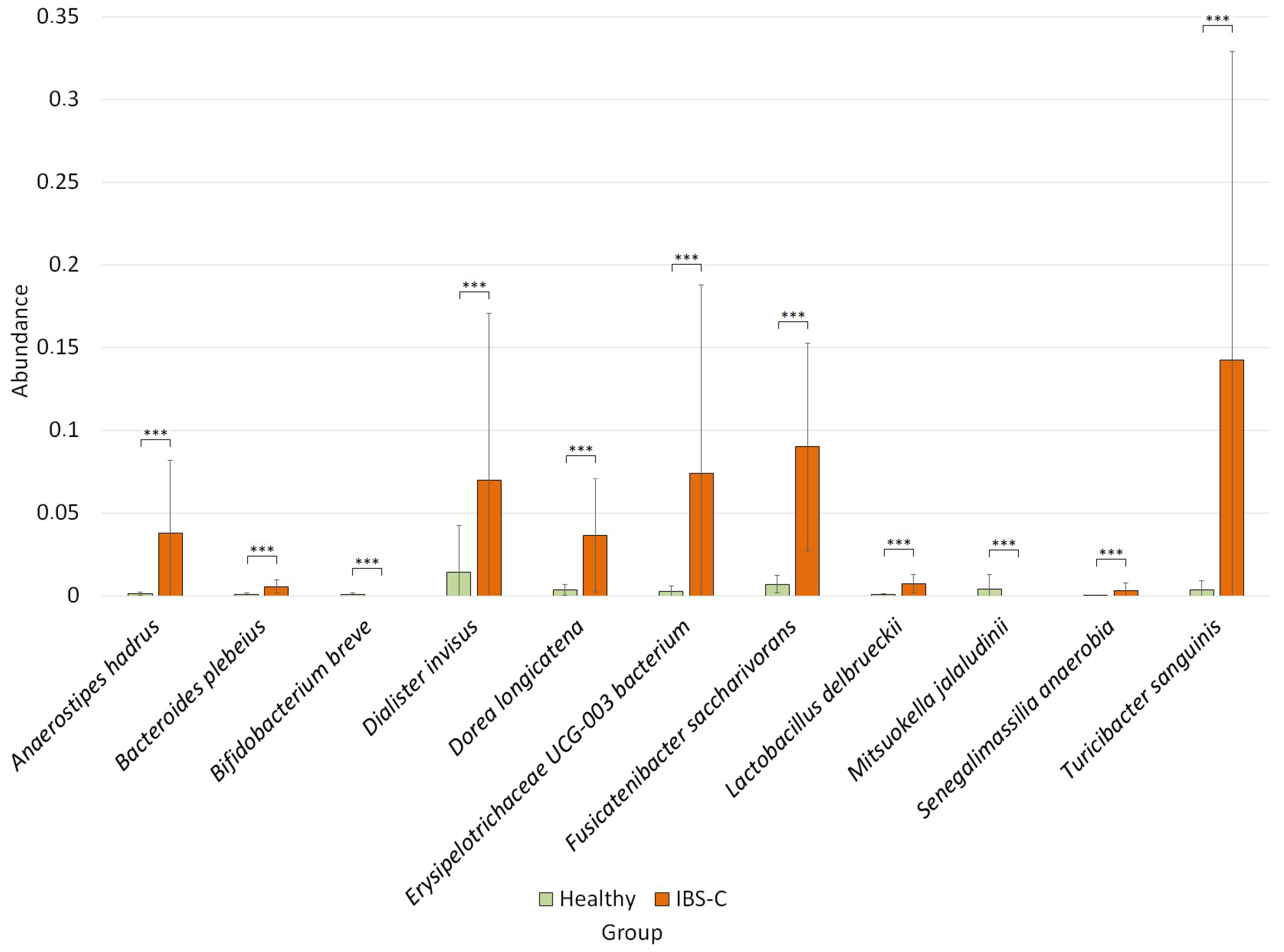
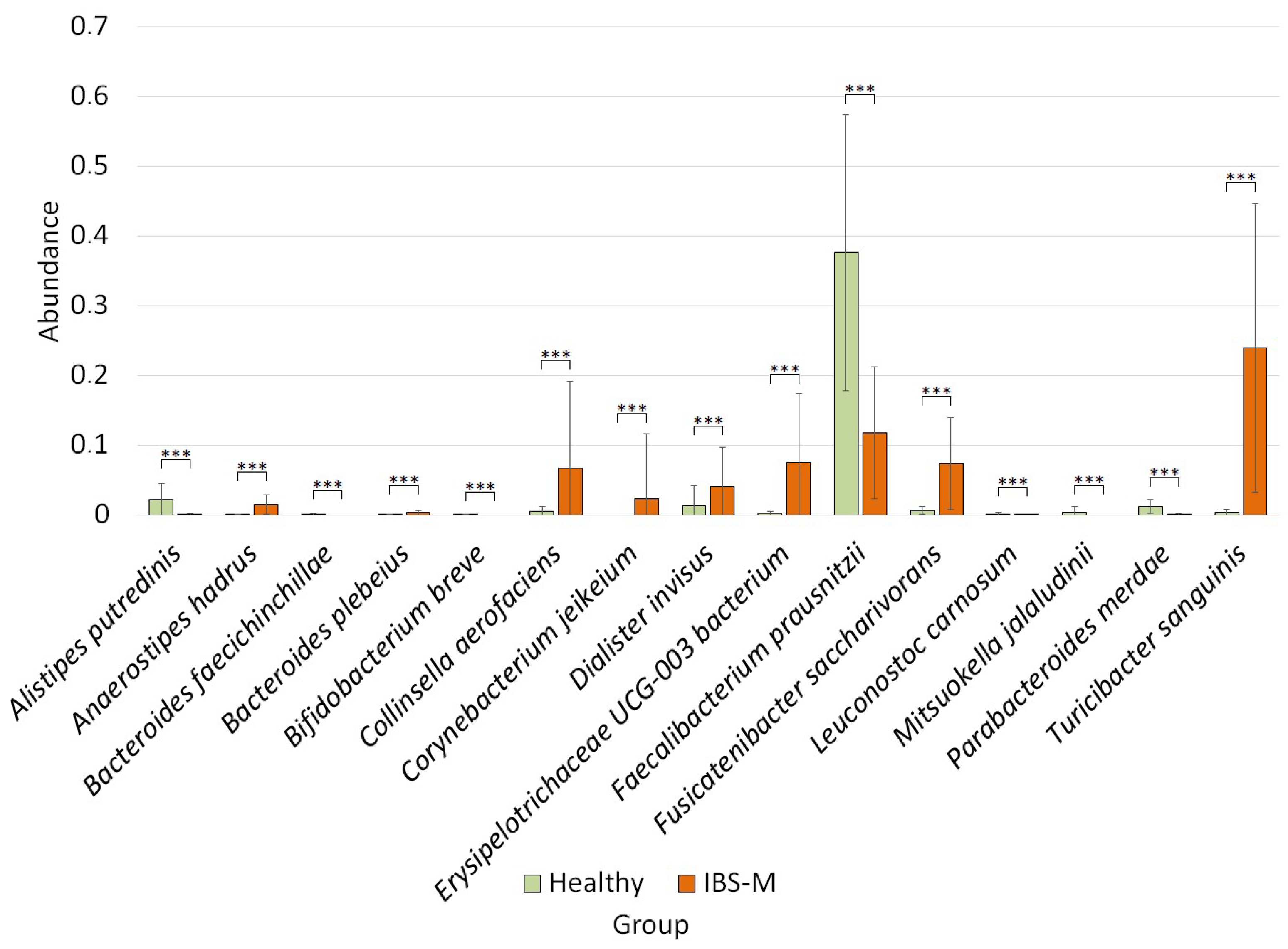
3.4. Differential Analysis of Bacterial Species Abundance between Studied Groups
4. Discussion
4.1. Changes in the Relative Abundance of Bacteria in the IBS-C, IBS-M, and Healthy Groups at the Phylum Level
4.2. Comparison of Relative Bacterial Abundance between IBS-C, IBS-M, and Healthy Groups at the Species Level
4.3. Specific Changes in the Composition of the Microbiome of the IBS-C Group
4.4. Changes in the Relative Abundance of Bacteria in the IBS-M Group
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 16S rRNA | 16S ribosomal RNA; |
| ASVs | amplicon sequence variants; |
| BSFS | Bristol Stool Form Scale; |
| dsDNA | double-stranded DNA; |
| FDR | false discovery rate; |
| IBD | Inflammatory bowel disease; |
| IBS | Irritable bowel syndrome; |
| IBS-C | constipation-predominant IBS; |
| IBS-C | constipation-predominant IBS; |
| IBS-D | diarrhoea-predominant IBS; |
| IBS-M | mixed fecal IBS; |
| NCBI | National Center for Biotechnology Information; |
| OTU | Operational taxonomic unit; |
| PCR | Polymerase chain reaction; |
| PGM | Personal Genome Machine; |
| SCFA | Short-chain fatty acid; |
| SD | standard deviation. |
References
- Shaikh, S.D.; Sun, N.; Canakis, A.; Park, W.Y.; Weber, H.C. Irritable Bowel Syndrome and the Gut Microbiome: A Comprehensive Review. J. Clin. Med. 2023, 12, 2558. [Google Scholar] [CrossRef] [PubMed]
- Canavan, C.; West, J.; Card, T. The Epidemiology of Irritable Bowel Syndrome. Clin. Epidemiol. 2014, 6, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Gravina, A.G.; Pellegrino, R.; Romeo, M.; Palladino, G.; Cipullo, M.; Iadanza, G.; Olivieri, S.; Zagaria, G.; Mazzarella, C.; Durante, T.; et al. The Burden of Irritable Bowel Syndrome in Medical and Nurse Italian University Student Population: The VANVITELLI-IBS Survey. Rev. Recent Clin. Trials 2023, 18, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Drossman, D.A. Functional Gastrointestinal Disorders: History, Pathophysiology, Clinical Features and Rome IV. Gastroenterology 2016, 150, 1262–1279. [Google Scholar] [CrossRef] [PubMed]
- Lacy, B.E.; Patel, N.K. Rome Criteria and a Diagnostic Approach to Irritable Bowel Syndrome. J. Clin. Med. 2017, 6, 99. [Google Scholar] [CrossRef] [PubMed]
- Ford, A.C.; Sperber, A.D.; Corsetti, M.; Camilleri, M. Irritable Bowel Syndrome. Lancet 2020, 396, 1675–1688. [Google Scholar] [CrossRef] [PubMed]
- Dean, B.B.; Aguilar, D.; Barghout, V.; Kahler, K.H.; Frech, F.; Groves, D.; Ofman, J.J. Impairment in Work Productivity and Health-Related Quality of Life in Patients with IBS. Am. J. Manag. Care 2005, 11, S17–S26. [Google Scholar] [PubMed]
- Longstreth, G.F.; Wilson, A.; Knight, K.; Wong, J.; Chiou, C.-F.; Barghout, V.; Frech, F.; Ofman, J.J. Irritable Bowel Syndrome, Health Care Use, and Costs: A U.S. Managed Care Perspective. Am. J. Gastroenterol. 2003, 98, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Rajilić-Stojanović, M.; Jonkers, D.M.; Salonen, A.; Hanevik, K.; Raes, J.; Jalanka, J.; de Vos, W.M.; Manichanh, C.; Golic, N.; Enck, P.; et al. Intestinal Microbiota and Diet in IBS: Causes, Consequences, or Epiphenomena? Am. J. Gastroenterol. 2015, 110, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Asha, M.Z.; Khalil, S.F.H. Efficacy and Safety of Probiotics, Prebiotics and Synbiotics in the Treatment of Irritable Bowel Syndrome: A Systematic Review and Meta-Analysis. Sultan Qaboos Univ. Med. J. 2020, 20, e13–e24. [Google Scholar] [CrossRef] [PubMed]
- Duboc, H.; Rainteau, D.; Rajca, S.; Humbert, L.; Farabos, D.; Maubert, M.; Grondin, V.; Jouet, P.; Bouhassira, D.; Seksik, P.; et al. Increase in Fecal Primary Bile Acids and Dysbiosis in Patients with Diarrhea-Predominant Irritable Bowel Syndrome. Neurogastroenterol. Motil. 2012, 24, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Xiong, L.; Li, L.; Li, M.; Chen, M. Alterations of Gut Microbiota in Patients with Irritable Bowel Syndrome: A Systematic Review and Meta-Analysis. J. Gastroenterol. Hepatol. 2017, 32, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Carroll, I.M.; Chang, Y.-H.; Park, J.; Sartor, R.B.; Ringel, Y. Luminal and Mucosal-Associated Intestinal Microbiota in Patients with Diarrhea-Predominant Irritable Bowel Syndrome. Gut Pathog. 2010, 2, 19. [Google Scholar] [CrossRef] [PubMed]
- Kerckhoffs, A.P.M.; Samsom, M.; van der Rest, M.E.; de Vogel, J.; Knol, J.; Ben-Amor, K.; Akkermans, L.M.A. Lower Bifidobacteria Counts in Both Duodenal Mucosa-Associated and Fecal Microbiota in Irritable Bowel Syndrome Patients. World J. Gastroenterol. 2009, 15, 2887–2892. [Google Scholar] [CrossRef] [PubMed]
- Rajilić-Stojanović, M.; Biagi, E.; Heilig, H.G.H.J.; Kajander, K.; Kekkonen, R.A.; Tims, S.; de Vos, W.M. Global and Deep Molecular Analysis of Microbiota Signatures in Fecal Samples from Patients with Irritable Bowel Syndrome. Gastroenterology 2011, 141, 1792–1801. [Google Scholar] [CrossRef] [PubMed]
- Malinen, E.; Rinttilä, T.; Kajander, K.; Mättö, J.; Kassinen, A.; Krogius, L.; Saarela, M.; Korpela, R.; Palva, A. Analysis of the Fecal Microbiota of Irritable Bowel Syndrome Patients and Healthy Controls with Real-Time PCR. Am. J. Gastroenterol. 2005, 100, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Gargari, G.; Mantegazza, G.; Cremon, C.; Taverniti, V.; Valenza, A.; Barbaro, M.R.; Marasco, G.; Duncan, R.; Fiore, W.; Ferrari, R.; et al. Collinsella Aerofaciens as a Predictive Marker of Response to Probiotic Treatment in Non-Constipated Irritable Bowel Syndrome. Gut Microbes 2024, 16, 2298246. [Google Scholar] [CrossRef] [PubMed]
- Scaldaferri, F.; Nardone, O.; Lopetuso, L.R.; Petito, V.; Bibbò, S.; Laterza, L.; Gerardi, V.; Bruno, G.; Scoleri, I.; Diroma, A.; et al. Intestinal Gas Production and Gastrointestinal Symptoms: From Pathogenesis to Clinical Implication. Eur. Rev. Med. Pharmacol. Sci. 2013, 17 (Suppl. S2), 2–10. [Google Scholar]
- Jahng, J.; Jung, I.S.; Choi, E.J.; Conklin, J.L.; Park, H. The Effects of Methane and Hydrogen Gases Produced by Enteric Bacteria on Ileal Motility and Colonic Transit Time. Neurogastroenterol. Motil. 2012, 24, 185–190. [Google Scholar] [CrossRef]
- Kumar, S.; Misra, A.; Ghoshal, U.C. Patients With Irritable Bowel Syndrome Exhale More Hydrogen Than Healthy Subjects in Fasting State. J. Neurogastroenterol. Motil. 2010, 16, 299–305. [Google Scholar] [CrossRef]
- Jalanka, J.; Salonen, A.; Fuentes, S.; de Vos, W.M. Microbial Signatures in Post-Infectious Irritable Bowel Syndrome--toward Patient Stratification for Improved Diagnostics and Treatment. Gut Microbes 2015, 6, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; Petito, V.; Graziani, C.; Schiavoni, E.; Paroni Sterbini, F.; Poscia, A.; Gaetani, E.; Franceschi, F.; Cammarota, G.; Sanguinetti, M.; et al. Gut Microbiota in Health, Diverticular Disease, Irritable Bowel Syndrome, and Inflammatory Bowel Diseases: Time for Microbial Marker of Gastrointestinal Disorders. Dig. Dis. 2018, 36, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Ermolenko, E.I.; Kudryavtsev, I.V.; Solovyova, O.I.; Karaseva, A.B.; Tsapieva, A.N.; Kondratenko, Y.D.; Kotyleva, M.P.; Serebryakova, M.K.; Shumikhina, I.A.; Suvorov, A.N. Profile of subpopulation composition of regulatory T lymphocytes and intestinal microbiota in patients with irritable bowel syndrome. Медицинская Иммунoлoгия 2020, 22, 335–346. [Google Scholar] [CrossRef]
- Chumpitazi, B.P.; Hollister, E.B.; Oezguen, N.; Tsai, C.M.; McMeans, A.R.; Luna, R.A.; Savidge, T.C.; Versalovic, J.; Shulman, R.J. Gut Microbiota Influences Low Fermentable Substrate Diet Efficacy in Children with Irritable Bowel Syndrome. Gut Microbes 2014, 5, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Hong, G.; Li, Y.; Yang, P.; Cheng, M.; Zhang, L.; Li, Y.; Ji, L.; Li, G.; Chen, C.; et al. Understanding of the Site-Specific Microbial Patterns Towards Accurate Identification for Patients with Diarrhea-Predominant Irritable Bowel Syndrome. Microbiol. Spectr. 2021, 9, e01255-21. [Google Scholar] [CrossRef] [PubMed]
- Rangel, I.; Sundin, J.; Fuentes, S.; Repsilber, D.; de Vos, W.M.; Brummer, R.J. The Relationship between Faecal-Associated and Mucosal-Associated Microbiota in Irritable Bowel Syndrome Patients and Healthy Subjects. Aliment. Pharmacol. Ther. 2015, 42, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Maharshak, N.; Ringel, Y.; Katibian, D.; Lundqvist, A.; Sartor, R.B.; Carroll, I.M.; Ringel-Kulka, T. Fecal and Mucosa-Associated Intestinal Microbiota in Patients with Diarrhea-Predominant Irritable Bowel Syndrome. Dig. Dis. Sci. 2018, 63, 1890–1899. [Google Scholar] [CrossRef] [PubMed]
- Pittayanon, R.; Lau, J.T.; Yuan, Y.; Leontiadis, G.I.; Tse, F.; Surette, M.; Moayyedi, P. Gut Microbiota in Patients With Irritable Bowel Syndrome—A Systematic Review. Gastroenterology 2019, 157, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Jalanka-Tuovinen, J.; Salojärvi, J.; Salonen, A.; Immonen, O.; Garsed, K.; Kelly, F.M.; Zaitoun, A.; Palva, A.; Spiller, R.C.; de Vos, W.M. Faecal Microbiota Composition and Host-Microbe Cross-Talk Following Gastroenteritis and in Postinfectious Irritable Bowel Syndrome. Gut 2014, 63, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, I.B.; O’Toole, P.W.; Öhman, L.; Claesson, M.J.; Deane, J.; Quigley, E.M.M.; Simrén, M. An Irritable Bowel Syndrome Subtype Defined by Species-Specific Alterations in Faecal Microbiota. Gut 2012, 61, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Tap, J.; Derrien, M.; Törnblom, H.; Brazeilles, R.; Cools-Portier, S.; Doré, J.; Störsrud, S.; Le Nevé, B.; Öhman, L.; Simrén, M. Identification of an Intestinal Microbiota Signature Associated With Severity of Irritable Bowel Syndrome. Gastroenterology 2017, 152, 111–123.e8. [Google Scholar] [CrossRef] [PubMed]
- Lluansí, A.; Llirós, M.; Carreras-Torres, R.; Bahí, A.; Capdevila, M.; Feliu, A.; Vilà-Quintana, L.; Elias-Masiques, N.; Cueva, E.; Peries, L.; et al. Impact of Bread Diet on Intestinal Dysbiosis and Irritable Bowel Syndrome Symptoms in Quiescent Ulcerative Colitis: A Pilot Study. PLoS ONE 2024, 19, e0297836. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Inoue, T.; Miyamoto, L.; Ono, Y.; Matsumoto, K.; Takeda, M.; Tsuchiya, K. Changes in Intestinal Microbiota and Biochemical Parameters in Patients with Inflammatory Bowel Disease and Irritable Bowel Syndrome Induced by the Prolonged Addition of Soluble Fibers to Usual Drug Therapy. J. Med. Investig. 2024, 71, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Vasapolli, R.; Krikonas, S.; Macke, L.; Gravdal, K.; Kirste, K.H.; Casén, C.; Storr, M.; Malfertheiner, P.; Schulz, C. Prolonged Intake of Luvos Healing Earth Does Not Alter the Composition of the Gut Microbiota in Patients with Diarrhea-Predominant Irritable Bowel Syndrome and Healthy Controls. J. Gastrointest. Liver Dis. 2024, 33, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple Statistical Identification and Removal of Contaminant Sequences in Marker-Gene and Metagenomics Data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [PubMed]
- Mallick, H.; Rahnavard, A.; McIver, L.J.; Ma, S.; Zhang, Y.; Nguyen, L.H.; Tickle, T.L.; Weingart, G.; Ren, B.; Schwager, E.H.; et al. Multivariable Association Discovery in Population-Scale Meta-Omics Studies. PLoS Comput. Biol. 2021, 17, e1009442. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Rudolph, P.E. Good, Ph.: Permutation Tests. A Practical Guide to Resampling Methods for Testing Hypotheses. Springer Series in Statistics, Springer-Verlag, Berlin—Heidelberg—New York: 1994, x, 228 pp., DM 74,00; ōS 577.20; sFr 74.–. ISBN 3-540-94097-9. Biom. J. 1995, 37, 150. [Google Scholar] [CrossRef]
- Duan, R.; Zhu, S.; Wang, B.; Duan, L. Alterations of Gut Microbiota in Patients With Irritable Bowel Syndrome Based on 16S rRNA-Targeted Sequencing: A Systematic Review. Clin. Transl. Gastroenterol. 2019, 10, e00012. [Google Scholar] [CrossRef] [PubMed]
- Distrutti, E.; Monaldi, L.; Ricci, P.; Fiorucci, S. Gut Microbiota Role in Irritable Bowel Syndrome: New Therapeutic Strategies. World J. Gastroenterol. 2016, 22, 2219–2241. [Google Scholar] [CrossRef] [PubMed]
- Pascale, A.; Marchesi, N.; Marelli, C.; Coppola, A.; Luzi, L.; Govoni, S.; Giustina, A.; Gazzaruso, C. Microbiota and Metabolic Diseases. Endocrine 2018, 61, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimarăes, V.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J.-P. The Firmicutes/Bacteroidetes Ratio of the Human Microbiota Changes with Age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of Diet in Shaping Gut Microbiota Revealed by a Comparative Study in Children from Europe and Rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Defrees, D.N.; Bailey, J. Irritable Bowel Syndrome: Epidemiology, Pathophysiology, Diagnosis, and Treatment. Prim. Care 2017, 44, 655–671. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, L.; Wang, X.; Wang, Z.; Zhang, J.; Jiang, R.; Wang, X.; Wang, K.; Liu, Z.; Xia, Z.; et al. Similar Fecal Microbiota Signatures in Patients With Diarrhea-Predominant Irritable Bowel Syndrome and Patients With Depression. Clin. Gastroenterol. Hepatol. 2016, 14, 1602–1611.e5. [Google Scholar] [CrossRef] [PubMed]
- Pozuelo, M.; Panda, S.; Santiago, A.; Mendez, S.; Accarino, A.; Santos, J.; Guarner, F.; Azpiroz, F.; Manichanh, C. Reduction of Butyrate- and Methane-Producing Microorganisms in Patients with Irritable Bowel Syndrome. Sci. Rep. 2015, 5, 12693. [Google Scholar] [CrossRef] [PubMed]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A Relevant Minority for the Maintenance of Gut Homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Lepage, P.; Nolte, A.; Hellmig, S.; Schreiber, S.; Ott, S.J. Transcriptional Activity of the Dominant Gut Mucosal Microbiota in Chronic Inflammatory Bowel Disease Patients. J. Med. Microbiol. 2010, 59, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Zheng, Z.; Shao, T.; Liu, L.; Xie, Z.; Le Chatelier, E.; He, Z.; Zhong, W.; Fan, Y.; Zhang, L.; et al. Quantitative Metagenomics Reveals Unique Gut Microbiome Biomarkers in Ankylosing Spondylitis. Genome Biol. 2017, 18, 142. [Google Scholar] [CrossRef]
- Sroka-Oleksiak, A.; Młodzińska, A.; Bulanda, M.; Salamon, D.; Major, P.; Stanek, M.; Gosiewski, T. Metagenomic Analysis of Duodenal Microbiota Reveals a Potential Biomarker of Dysbiosis in the Course of Obesity and Type 2 Diabetes: A Pilot Study. J. Clin. Med. 2020, 9, 369. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Ye, D.; Yang, H.; Song, J.; Sun, X.; He, Z.; Mao, Y.; Hao, G. Assessing the Relationship between Gut Microbiota and Irritable Bowel Syndrome: A Two-Sample Mendelian Randomization Analysis. BMC Gastroenterol. 2023, 23, 150. [Google Scholar] [CrossRef] [PubMed]
- Nagel, R.; Traub, R.J.; Allcock, R.J.N.; Kwan, M.M.S.; Bielefeldt-Ohmann, H. Comparison of Faecal Microbiota in Blastocystis-Positive and Blastocystis-Negative Irritable Bowel Syndrome Patients. Microbiome 2016, 4, 47. [Google Scholar] [CrossRef] [PubMed]
- Ringel-Kulka, T.; Benson, A.K.; Carroll, I.M.; Kim, J.; Legge, R.M.; Ringel, Y. Molecular Characterization of the Intestinal Microbiota in Patients with and without Abdominal Bloating. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G417–G426. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.-S.; Chang, P.-F.; Liao, C.-H.; Lee, T.-H.; Chen, Y.; Lee, Y.-C.; Wu, M.-S.; Wang, H.-P.; Ni, Y.-H. Differences of Microbiota in Small Bowel and Faeces between Irritable Bowel Syndrome Patients and Healthy Subjects. Scand. J. Gastroenterol. 2016, 51, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Zhang, C.; Ren, X.; Tai, B.; Xing, F. Metagenome Analysis Identifies Microbial Shifts upon Deoxynivalenol Exposure and Post-Exposure Recovery in the Mouse Gut. Toxins 2023, 15, 243. [Google Scholar] [CrossRef] [PubMed]
- Dubourg, G.; Lagier, J.-C.; Armougom, F.; Robert, C.; Audoly, G.; Papazian, L.; Raoult, D. High-Level Colonisation of the Human Gut by Verrucomicrobia Following Broad-Spectrum Antibiotic Treatment. Int. J. Antimicrob. Agents 2013, 41, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Barandouzi, Z.A.; Lee, J.; Maas, K.; Starkweather, A.R.; Cong, X.S. Altered Gut Microbiota in Irritable Bowel Syndrome and Its Association with Food Components. J. Pers. Med. 2021, 11, 35. [Google Scholar] [CrossRef]
- Barandouzi, Z.A.; Lee, J.; Del Carmen Rosas, M.; Chen, J.; Henderson, W.A.; Starkweather, A.R.; Cong, X.S. Associations of Neurotransmitters and the Gut Microbiome with Emotional Distress in Mixed Type of Irritable Bowel Syndrome. Sci. Rep. 2022, 12, 1648. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, A.; Coppola, G.; Santopaolo, F.; Gasbarrini, A.; Ponziani, F.R. Role of Akkermansia in Human Diseases: From Causation to Therapeutic Properties. Nutrients 2023, 15, 1815. [Google Scholar] [CrossRef]
- Aggarwal, V.; Sunder, S.; Verma, S.R. Disease-Associated Dysbiosis and Potential Therapeutic Role of Akkermansia muciniphila, a Mucus Degrading Bacteria of Gut Microbiome. Folia Microbiol. 2022, 67, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Han, R.; Yuan, Y.; Xing, Y.; Zhang, W.; Sun, Z.; Liu, Y.; Li, J.; Mao, T. The Role of Akkermansia muciniphila in Inflammatory Bowel Disease: Current Knowledge and Perspectives. Front. Immunol. 2022, 13, 1089600. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Ji, X.; Lu, G.; Zhang, F. The Potential of Akkermansia muciniphila in Inflammatory Bowel Disease. Appl. Microbiol. Biotechnol. 2021, 105, 5785–5794. [Google Scholar] [CrossRef] [PubMed]
- Mruk-Mazurkiewicz, H.; Kulaszyńska, M.; Czarnecka, W.; Podkówka, A.; Ekstedt, N.; Zawodny, P.; Wierzbicka-Woś, A.; Marlicz, W.; Skupin, B.; Stachowska, E.; et al. Insights into the Mechanisms of Action of Akkermansia muciniphila in the Treatment of Non-Communicable Diseases. Nutrients 2024, 16, 1695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wu, Y.; Wang, J.; Wu, G.; Long, W.; Xue, Z.; Wang, L.; Zhang, X.; Pang, X.; Zhao, Y.; et al. Accelerated Dysbiosis of Gut Microbiota during Aggravation of DSS-Induced Colitis by a Butyrate-Producing Bacterium. Sci. Rep. 2016, 6, 27572. [Google Scholar] [CrossRef]
- Abdugheni, R.; Wang, W.-Z.; Wang, Y.-J.; Du, M.-X.; Liu, F.-L.; Zhou, N.; Jiang, C.-Y.; Wang, C.-Y.; Wu, L.; Ma, J.; et al. Metabolite Profiling of Human-Originated Lachnospiraceae at the Strain Level. iMeta 2022, 1, e58. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Duncan, S.H.; McCrae, S.I.; Millar, J.; Jackson, M.S.; Flint, H.J. Restricted Distribution of the Butyrate Kinase Pathway among Butyrate-Producing Bacteria from the Human Colon. J. Bacteriol. 2004, 186, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Endo, A.; Tanno, H.; Kadowaki, R.; Fujii, T.; Tochio, T. Extracellular Fructooligosaccharide Degradation in Anaerostipes hadrus for Co-Metabolism with Non-Fructooligosaccharide Utilizers. Biochem. Biophys. Res. Commun. 2022, 613, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, H.; Zhao, X.; Zhou, R.; Liu, H.; Sun, Y.; Fan, Y.; Shi, Y.; Qiao, S.; Liu, S.; et al. Multi-Omics Study Reveals That Statin Therapy Is Associated with Restoration of Gut Microbiota Homeostasis and Improvement in Outcomes in Patients with Acute Coronary Syndrome. Theranostics 2021, 11, 5778–5793. [Google Scholar] [CrossRef] [PubMed]
- Ali, R.O.; Quinn, G.M.; Umarova, R.; Haddad, J.A.; Zhang, G.Y.; Townsend, E.C.; Scheuing, L.; Hill, K.L.; Gewirtz, M.; Rampertaap, S.; et al. Longitudinal Multi-Omics Analyses of the Gut-Liver Axis Reveals Metabolic Dysregulation in Hepatitis C Infection and Cirrhosis. Nat. Microbiol. 2023, 8, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Meydan, C.; Afshinnekoo, E.; Rickard, N.; Daniels, G.; Kunces, L.; Hardy, T.; Lili, L.; Pesce, S.; Jacobson, P.; Mason, C.E.; et al. Improved Gastrointestinal Health for Irritable Bowel Syndrome with Metagenome-Guided Interventions. Precis. Clin. Med. 2020, 3, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Agnello, M.; Carroll, L.N.; Imam, N.; Pino, R.; Palmer, C.; Varas, I.; Greene, C.; Hitschfeld, M.; Gupta, S.; Almonacid, D.E.; et al. Gut Microbiome Composition and Risk Factors in a Large Cross-Sectional IBS Cohort. BMJ Open Gastroenterol. 2020, 7, e000345. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Xie, L.-S.; Lian, S.; Li, K.; Yang, Y.; Wang, W.-Z.; Hu, S.; Liu, S.-J.; Liu, C.; He, Z. Anaerostipes Hadrus, a Butyrate-Producing Bacterium Capable of Metabolizing 5-Fluorouracil. mSphere 2024, 9, e0081623. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Han, L.; Zhao, L.; Zhou, Y.; Yang, C.; Xiong, T.; Luo, W.; Chen, Y.; Qiu, Q.; Shang, X.; et al. Integrated Multi-Omics Data Analysis Reveals Altered Metabolome Activity and Microbiome Composition in Irritable Bowel Syndrome. 2020. Available online: https://www.researchsquare.com/article/rs-51066/v1 (accessed on 29 April 2024).
- Gargari, G.; Mantegazza, G.; Taverniti, V.; Gardana, C.; Valenza, A.; Rossignoli, F.; Barbaro, M.R.; Marasco, G.; Cremon, C.; Barbara, G.; et al. Fecal Short-Chain Fatty Acids in Non-Constipated Irritable Bowel Syndrome: A Potential Clinically Relevant Stratification Factor Based on Catabotyping Analysis. Gut Microbes 2023, 15, 2274128. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Everard, A. Talking Microbes: When Gut Bacteria Interact with Diet and Host Organs. Mol. Nutr. Food Res. 2016, 60, 58–66. [Google Scholar] [CrossRef]
- Pimentel, M.; Mayer, A.G.; Park, S.; Chow, E.J.; Hasan, A.; Kong, Y. Methane Production during Lactulose Breath Test Is Associated with Gastrointestinal Disease Presentation. Dig. Dis. Sci. 2003, 48, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.P.; Lagishetty, V.; Hauer, M.C.; Labus, J.S.; Dong, T.S.; Toma, R.; Vuyisich, M.; Naliboff, B.D.; Lackner, J.M.; Gupta, A.; et al. Multi-Omics Profiles of the Intestinal Microbiome in Irritable Bowel Syndrome and Its Bowel Habit Subtypes. Microbiome 2023, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Welham, Z.; Li, J.; Engel, A.F.; Molloy, M.P. Mucosal Microbiome in Patients with Early Bowel Polyps: Inferences from Short-Read and Long-Read 16S rRNA Sequencing. Cancers 2023, 15, 5045. [Google Scholar] [CrossRef]
- Nagao-Kitamoto, H.; Kamada, N. Host-Microbial Cross-Talk in Inflammatory Bowel Disease. Immune Netw. 2017, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Joossens, M.; Huys, G.; Cnockaert, M.; De Preter, V.; Verbeke, K.; Rutgeerts, P.; Vandamme, P.; Vermeire, S. Dysbiosis of the Faecal Microbiota in Patients with Crohn’s Disease and Their Unaffected Relatives. Gut 2011, 60, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Rigsbee, L.; Agans, R.; Shankar, V.; Kenche, H.; Khamis, H.J.; Michail, S.; Paliy, O. Quantitative Profiling of Gut Microbiota of Children with Diarrhea-Predominant Irritable Bowel Syndrome. Am. J. Gastroenterol. 2012, 107, 1740–1751. [Google Scholar] [CrossRef] [PubMed]
- Fourie, N.H.; Wang, D.; Abey, S.K.; Sherwin, L.B.; Joseph, P.V.; Rahim-Williams, B.; Ferguson, E.G.; Henderson, W.A. The Microbiome of the Oral Mucosa in Irritable Bowel Syndrome. Gut Microbes 2016, 7, 286–301. [Google Scholar] [CrossRef] [PubMed]
- Vervier, K.; Moss, S.; Kumar, N.; Adoum, A.; Barne, M.; Browne, H.; Kaser, A.; Kiely, C.J.; Neville, B.A.; Powell, N.; et al. Two Microbiota Subtypes Identified in Irritable Bowel Syndrome with Distinct Responses to the Low FODMAP Diet. Gut 2022, 71, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Fukui, H.; Oshima, T.; Tanaka, Y.; Oikawa, Y.; Makizaki, Y.; Ohno, H.; Tomita, T.; Watari, J.; Miwa, H. Effect of Probiotic Bifidobacterium Bifidum G9-1 on the Relationship between Gut Microbiota Profile and Stress Sensitivity in Maternally Separated Rats. Sci. Rep. 2018, 8, 12384. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Zhao, X.; Cui, X.; Mao, X.; Tang, N.; Jiao, C.; Wang, D.; Zhang, Y.; Ye, Z.; Zhang, H. Characterization of Fungal and Bacterial Dysbiosis in Young Adult Chinese Patients with Crohn’s Disease. Ther. Adv. Gastroenterol. 2020, 13, 1756284820971202. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Lee, M.; Yang, S.; Kim, K.; Yong, D.; Kim, H.R.; Lee, I. Human Reference Gut Microbiome Catalog Including Newly Assembled Genomes from Under-Represented Asian Metagenomes. Genome Med. 2021, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.M.; Margolis, K.G. Building Community in the Gut: A Role for Mucosal Serotonin. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Cotozzolo, E.; Cremonesi, P.; Curone, G.; Menchetti, L.; Riva, F.; Biscarini, F.; Marongiu, M.L.; Castrica, M.; Castiglioni, B.; Miraglia, D.; et al. Characterization of Bacterial Microbiota Composition along the Gastrointestinal Tract in Rabbits. Animals 2020, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and Their Role as Members of the Human Gut Microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [PubMed]
- Cronin, M.; Ventura, M.; Fitzgerald, G.F.; van Sinderen, D. Progress in Genomics, Metabolism and Biotechnology of Bifidobacteria. Int. J. Food Microbiol. 2011, 149, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Torres-Maravilla, E.; Holowacz, S.; Delannoy, J.; Lenoir, L.; Jacouton, E.; Gervason, S.; Meynier, M.; Boucard, A.-S.; Carvalho, F.A.; Barbut, F.; et al. Serpin-Positive Bifidobacterium Breve CNCM I-5644 Improves Intestinal Permeability in Two Models of Irritable Bowel Syndrome. Sci. Rep. 2022, 12, 19776. [Google Scholar] [CrossRef] [PubMed]
- Bozzi Cionci, N.; Baffoni, L.; Gaggìa, F.; Di Gioia, D. Therapeutic Microbiology: The Role of Bifidobacterium Breve as Food Supplement for the Prevention/Treatment of Paediatric Diseases. Nutrients 2018, 10, 1723. [Google Scholar] [CrossRef] [PubMed]
- Lan, G.Q.; Ho, Y.W.; Abdullah, N. Mitsuokella jalaludinii sp. Nov., from the Rumens of Cattle in Malaysia. Int. J. Syst. Evol. Microbiol. 2002, 52, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Savin, K.W.; Moate, P.J.; Williams, S.R.O.; Bath, C.; Hemsworth, J.; Wang, J.; Ram, D.; Zawadzki, J.; Rochfort, S.; Cocks, B.G. Dietary Wheat and Reduced Methane Yield Are Linked to Rumen Microbiome Changes in Dairy Cows. PLoS ONE 2022, 17, e0268157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, H.; Wang, S.; Zhang, W.; Wang, J.; Tian, H.; Wang, Y.; Ji, H. Fecal Microbiota and Its Correlation With Fatty Acids and Free Amino Acids Metabolism in Piglets After a Lactobacillus Strain Oral Administration. Front. Microbiol. 2019, 10, 785. [Google Scholar] [CrossRef] [PubMed]
- Taras, D.; Simmering, R.; Collins, M.D.; Lawson, P.A.; Blaut, M. Reclassification of Eubacterium Formicigenerans Holdeman and Moore 1974 as Dorea formicigenerans gen. nov., comb. nov., and Description of Dorea longicatena sp. nov., Isolated from Human Faeces. Int. J. Syst. Evol. Microbiol. 2002, 52, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Altomare, A.; Di Rosa, C.; Imperia, E.; Emerenziani, S.; Cicala, M.; Guarino, M.P.L. Diarrhea Predominant-Irritable Bowel Syndrome (IBS-D): Effects of Different Nutritional Patterns on Intestinal Dysbiosis and Symptoms. Nutrients 2021, 13, 1506. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Shen, X.; Shi, X.; Sakandar, H.A.; Quan, K.; Li, Y.; Jin, H.; Kwok, L.-Y.; Zhang, H.; Sun, Z. Targeting Gut Microbiota and Metabolism as the Major Probiotic Mechanism—An Evidence-Based Review. Trends Food Sci. Technol. 2023, 138, 178–198. [Google Scholar] [CrossRef]
- Tarique, M.; Ali, A.H.; Kizhakkayil, J.; Gan, R.-Y.; Liu, S.-Q.; Kamal-Eldin, A.; Ayyash, M. Investigating the Biological Activities and Prebiotic Potential of Exopolysaccharides Produced by Lactobacillus delbrueckii and Lacticaseibacillus rhamnosus: Implications for Gut Microbiota Modulation and Rheological Properties in Fermented Milk. Food Hydrocoll. Health 2023, 4, 100162. [Google Scholar] [CrossRef]
- Wang, L.; Alammar, N.; Singh, R.; Nanavati, J.; Song, Y.; Chaudhary, R.; Mullin, G.E. Gut Microbial Dysbiosis in the Irritable Bowel Syndrome: A Systematic Review and Meta-Analysis of Case-Control Studies. J. Acad. Nutr. Diet. 2020, 120, 565–586. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Lu, X.; Shi, H.; Zhao, G.; Song, Y.; Wang, Y.; Zhang, J.; Jin, Y.; Wang, S. Distinct Microbial Populations Exist in the Mucosa-Associated Microbiota of Diarrhea Predominant Irritable Bowel Syndrome and Ulcerative Colitis. J. Clin. Gastroenterol. 2019, 53, 660–672. [Google Scholar] [CrossRef] [PubMed]
- Lagier, J.-C.; Elkarkouri, K.; Rivet, R.; Couderc, C.; Raoult, D.; Fournier, P.-E. Non Contiguous-Finished Genome Sequence and Description of Senegalemassilia anaerobia Gen. Nov., sp. Nov. Stand. Genomic Sci. 2013, 7, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Adamberg, K.; Adamberg, S.; Ernits, K.; Larionova, A.; Voor, T.; Jaagura, M.; Visnapuu, T.; Alamäe, T. Composition and Metabolism of Fecal Microbiota from Normal and Overweight Children Are Differentially Affected by Melibiose, Raffinose and Raffinose-Derived Fructans. Anaerobe 2018, 52, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, A.; Benno, Y.; Nakase, T. Phylogenetic and Phenotypic Evidence for the Transfer of Eubacterium Aerofaciens to the Genus Collinsella as Collinsella aerofaciens Gen. Nov., Comb. Nov. Int. J. Syst. Bacteriol. 1999, 49 Pt 2, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Bag, S.; Ghosh, T.S.; Das, B. Complete Genome Sequence of Collinsella Aerofaciens Isolated from the Gut of a Healthy Indian Subject. Genome Announc. 2017, 5, e01361-17. [Google Scholar] [CrossRef] [PubMed]
- Masoodi, I.; Alshanqeeti, A.S.; Alyamani, E.J.; AlLehibi, A.A.; Alqutub, A.N.; Alsayari, K.N.; Alomair, A.O. Microbial Dysbiosis in Irritable Bowel Syndrome: A Single-Center Metagenomic Study in Saudi Arabia. JGH Open 2020, 4, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Gargari, G.; Taverniti, V.; Gardana, C.; Cremon, C.; Canducci, F.; Pagano, I.; Barbaro, M.R.; Bellacosa, L.; Castellazzi, A.M.; Valsecchi, C.; et al. Fecal Clostridiales Distribution and Short-Chain Fatty Acids Reflect Bowel Habits in Irritable Bowel Syndrome. Environ. Microbiol. 2018, 20, 3201–3213. [Google Scholar] [CrossRef] [PubMed]
- Tauch, A.; Burkovski, A. Molecular Armory or Niche Factors: Virulence Determinants of Corynebacterium Species. FEMS Microbiol. Lett. 2015, 362, fnv185. [Google Scholar] [CrossRef]
- Moore Pardo, S.M.; Patel, R.H.; Ramsakal, A.; Greene, J. Disseminated Corynebacterium jeikeium Infection in Cancer Patients. Cureus 2020, 12, e8764. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zeng, S.; Wang, X.; Liu, H.; Sun, H.; Zhou, X.; Yang, Q. Patterns of Circulating Microbiota during the Acute Phase Following ST-Segment Elevation Myocardial Infarction Predict Long-Term Cardiovascular Events. Int. Heart J. 2023, 64, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Barbara, G.; Cremon, C.; Stanghellini, V. Inflammatory Bowel Disease and Irritable Bowel Syndrome: Similarities and Differences. Curr. Opin. Gastroenterol. 2014, 30, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, M.; Lasch, K.; Zhou, W. Irritable Bowel Syndrome: Methods, Mechanisms, and Pathophysiology. The Confluence of Increased Permeability, Inflammation, and Pain in Irritable Bowel Syndrome. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G775–G785. [Google Scholar] [CrossRef] [PubMed]
- El-Salhy, M. Intestinal Bacteria Associated with Irritable Bowel Syndrome and Chronic Fatigue. Neurogastroenterol. Motil. 2023, 35, e14621. [Google Scholar] [CrossRef] [PubMed]
- Saulnier, D.M.; Riehle, K.; Mistretta, T.-A.; Diaz, M.-A.; Mandal, D.; Raza, S.; Weidler, E.M.; Qin, X.; Coarfa, C.; Milosavljevic, A.; et al. Gastrointestinal Microbiome Signatures of Pediatric Patients with Irritable Bowel Syndrome. Gastroenterology 2011, 141, 1782–1791. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel Disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Brusaferro, A.; Cozzali, R.; Orabona, C.; Biscarini, A.; Farinelli, E.; Cavalli, E.; Grohmann, U.; Principi, N.; Esposito, S. Is It Time to Use Probiotics to Prevent or Treat Obesity? Nutrients 2018, 10, 1613. [Google Scholar] [CrossRef] [PubMed]
- Duboc, H.; Rajca, S.; Rainteau, D.; Benarous, D.; Maubert, M.-A.; Quervain, E.; Thomas, G.; Barbu, V.; Humbert, L.; Despras, G.; et al. Connecting Dysbiosis, Bile-Acid Dysmetabolism and Gut Inflammation in Inflammatory Bowel Diseases. Gut 2013, 62, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S. Fermentation in the Human Large Intestine: Its Physiologic Consequences and the Potential Contribution of Prebiotics. J. Clin. Gastroenterol. 2011, 45, S120–S127. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Shen, J.; Ran, Z.H. Association between Faecalibacterium Prausnitzii Reduction and Inflammatory Bowel Disease: A Meta-Analysis and Systematic Review of the Literature. Gastroenterol. Res. Pract. 2014, 2014, 872725. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhao, S.; Li, Y. Faecalibacterium Prausnitzii: A Next-Generation Probiotic in Gut Disease Improvement. Can. J. Infect. Dis. Med. Microbiol. 2021, 2021, e6666114. [Google Scholar] [CrossRef]
- Martín, R.; Bermúdez-Humarán, L.G.; Langella, P. Searching for the Bacterial Effector: The Example of the Multi-Skilled Commensal Bacterium Faecalibacterium prausnitzii. Front. Microbiol. 2018, 9, 346. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.; Saad, R.J.; Long, M.D.; Rao, S.S.C. ACG Clinical Guideline: Small Intestinal Bacterial Overgrowth. Am. J. Gastroenterol. 2020, 115, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, S.; Spampinato, G.; Candeliere, F.; Amaretti, A.; Brun, P.; Castagliuolo, I.; Rossi, M. Phenotypic Traits and Immunomodulatory Properties of Leuconostoc carnosum Isolated From Meat Products. Front. Microbiol. 2021, 12, 730827. [Google Scholar] [CrossRef] [PubMed]
- Sanz, Y.; Sánchez, E.; Marzotto, M.; Calabuig, M.; Torriani, S.; Dellaglio, F. Differences in Faecal Bacterial Communities in Coeliac and Healthy Children as Detected by PCR and Denaturing Gradient Gel Electrophoresis. FEMS Immunol. Med. Microbiol. 2007, 51, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, A.C.; Salminen, S.J. The Health Effects of Cultured Milk Products with Viable and Non-Viable Bacteria. Int. Dairy J. 1998, 8, 749–758. [Google Scholar] [CrossRef]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.-Y. Probiotics and Their Fermented Food Products Are Beneficial for Health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-Level Analysis of Gut Microbiome Variation. Science 2016, 352, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhang, L.; Wang, X.; Yi, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Roles of Intestinal Parabacteroides in Human Health and Diseases. FEMS Microbiol. Lett. 2022, 369, fnac072. [Google Scholar] [CrossRef] [PubMed]
- Zitomersky, N.L.; Atkinson, B.J.; Franklin, S.W.; Mitchell, P.D.; Snapper, S.B.; Comstock, L.E.; Bousvaros, A. Characterization of Adherent Bacteroidales from Intestinal Biopsies of Children and Young Adults with Inflammatory Bowel Disease. PLoS ONE 2013, 8, e63686. [Google Scholar] [CrossRef] [PubMed]
- Haro, C.; Garcia-Carpintero, S.; Alcala-Diaz, J.F.; Gomez-Delgado, F.; Delgado-Lista, J.; Perez-Martinez, P.; Rangel Zuñiga, O.A.; Quintana-Navarro, G.M.; Landa, B.B.; Clemente, J.C.; et al. The Gut Microbial Community in Metabolic Syndrome Patients Is Modified by Diet. J. Nutr. Biochem. 2016, 27, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Becerra, L.; Cornejo-Granados, F.; García-López, R.; Valdez-Lara, A.; Bikel, S.; Canizales-Quinteros, S.; López-Contreras, B.E.; Mendoza-Vargas, A.; Nielsen, H.; Ochoa-Leyva, A. Metatranscriptomic Analysis to Define the Secrebiome, and 16S rRNA Profiling of the Gut Microbiome in Obesity and Metabolic Syndrome of Mexican Children. Microb. Cell Factories 2020, 19, 61. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y.-S.; Lu, J.-H.; Li, S.-H.; Li, J.-H.; Yuan, M.-Y.; He, J.-R.; Chen, N.-N.; Xiao, W.-Q.; Shen, S.-Y.; Qiu, L.; et al. Connections between the Human Gut Microbiome and Gestational Diabetes Mellitus. Gigascience 2017, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, Y.D.; Gryaznova, M.V.; Burakova, I.Y.; Syromyatnikov, M.Y.; Sviridova, T.N.; Lebedeva, O.P.; Maslov, A.Y.; Popov, V.N. Study of Microbiome Aberrations in Patients with Irritable Bowel Syndrome with Diarrhea by Next-Generation Sequencing. Res. Results Biomed. 2023, 9, 446–460. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Height ± SD, cm | Weight ± SD, kg | BMI ± SD | Age ± SD | Duration of the Disease |
|---|---|---|---|---|---|
| Control | 180.92 ± 8.04 | 77.83 ± 7.64 | 23.79 ± 2.19 | 30.41 ± 6.42 | — |
| IBS-C | 178.43 ± 8.62 | 74.14 ± 5.88 | 23.96 ± 2.61 | 32.67 ± 6.67 | 5.36 ± 4.43 |
| IBS-M | 170.80 ± 10.27 | 65.11 ± 12.12 | 22.59 ± 2.02 | 30.14 ± 6.79 | 5.50 ± 3.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gryaznova, M.; Smirnova, Y.; Burakova, I.; Morozova, P.; Lagutina, S.; Chizhkov, P.; Korneeva, O.; Syromyatnikov, M. Fecal Microbiota Characteristics in Constipation-Predominant and Mixed-Type Irritable Bowel Syndrome. Microorganisms 2024, 12, 1414. https://doi.org/10.3390/microorganisms12071414
Gryaznova M, Smirnova Y, Burakova I, Morozova P, Lagutina S, Chizhkov P, Korneeva O, Syromyatnikov M. Fecal Microbiota Characteristics in Constipation-Predominant and Mixed-Type Irritable Bowel Syndrome. Microorganisms. 2024; 12(7):1414. https://doi.org/10.3390/microorganisms12071414
Chicago/Turabian StyleGryaznova, Mariya, Yuliya Smirnova, Inna Burakova, Polina Morozova, Svetlana Lagutina, Pavel Chizhkov, Olga Korneeva, and Mikhail Syromyatnikov. 2024. "Fecal Microbiota Characteristics in Constipation-Predominant and Mixed-Type Irritable Bowel Syndrome" Microorganisms 12, no. 7: 1414. https://doi.org/10.3390/microorganisms12071414