
Oat Brewery Waste Decreased Methane Production and Alters Rumen Fermentation, Microbiota Composition, and CAZymes Profiles

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Material, Treatments and Composition
2.2. Rumen Fluid Donor
2.3. Total Gas and CH4 Production
2.4. In Vitro Dry Matter Digestibility (IVDMD)
2.5. Volatile Fatty Acid (VFA) and Ammonia-N
2.6. Protozoa Enumeration
2.7. DNA Extraction
2.8. Shotgun Metagenome Sequencing
2.9. Bioinformatics Analysis
2.10. Gene Prediction and CAZyme Annotation
2.11. Statistical Analysis
3. Results
3.1. Chemical Composition
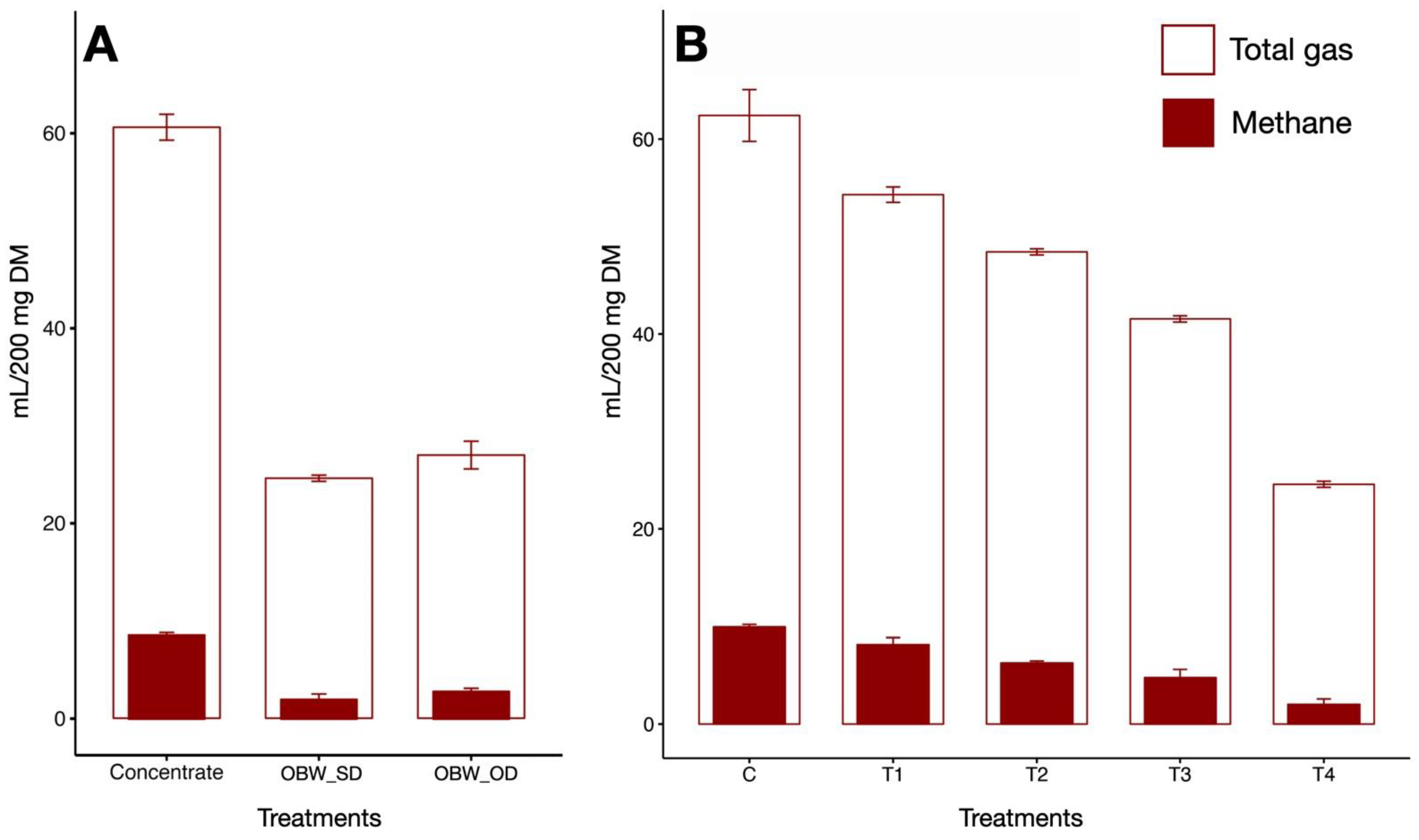
3.2. CH4 Production
3.3. Total Gas
3.4. In Vitro Dry Matter Digestibility (IVDMD) and Organic Matter Digestibility (OMD)
3.5. VFA, Ammonia-N and Protozoa
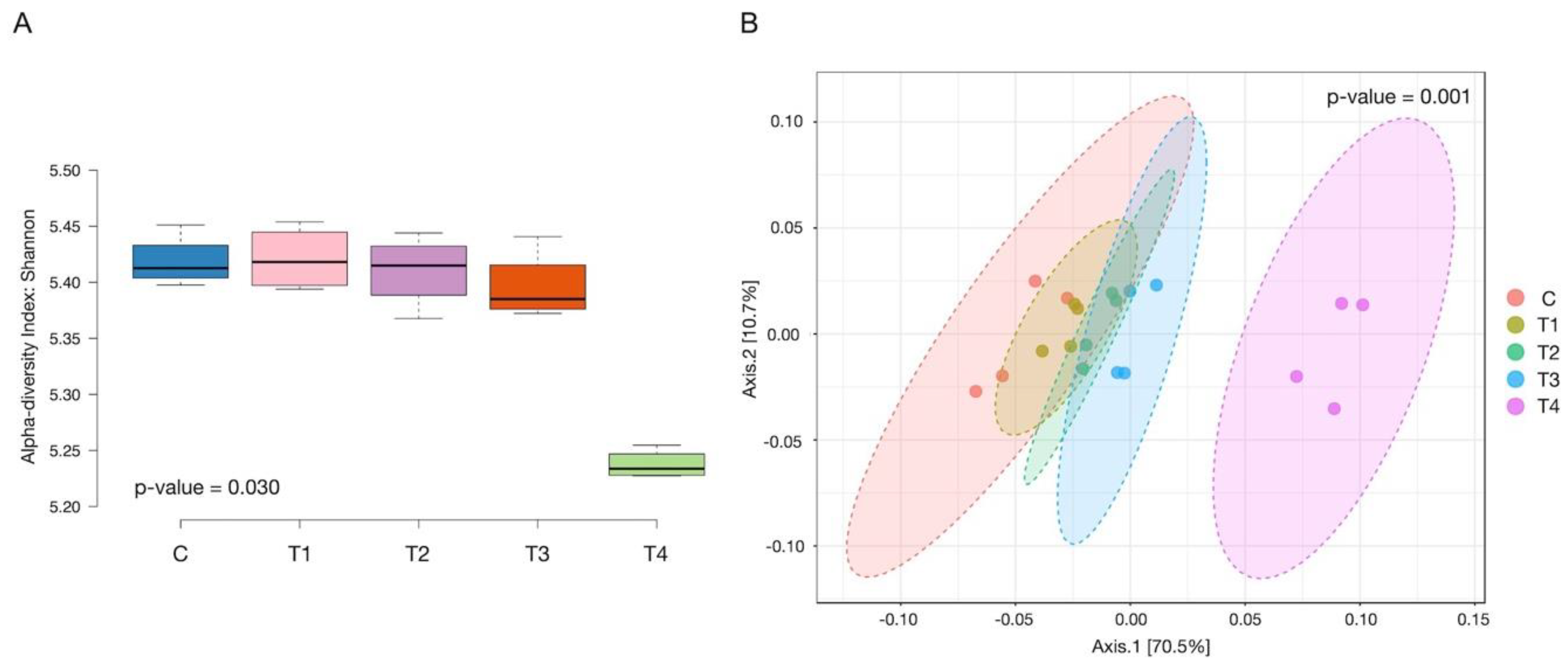
3.6. Alpha and Beta Diversity
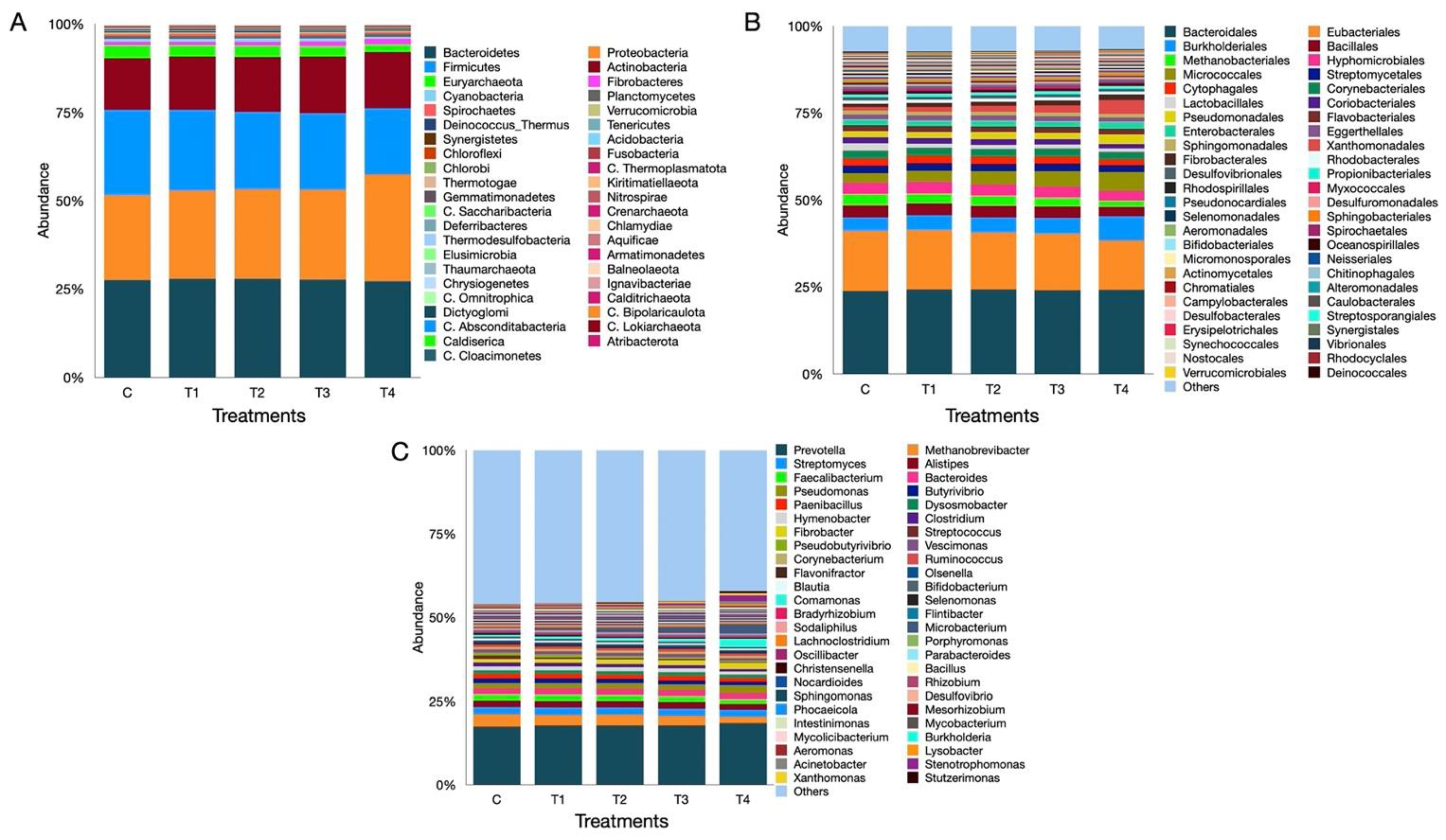
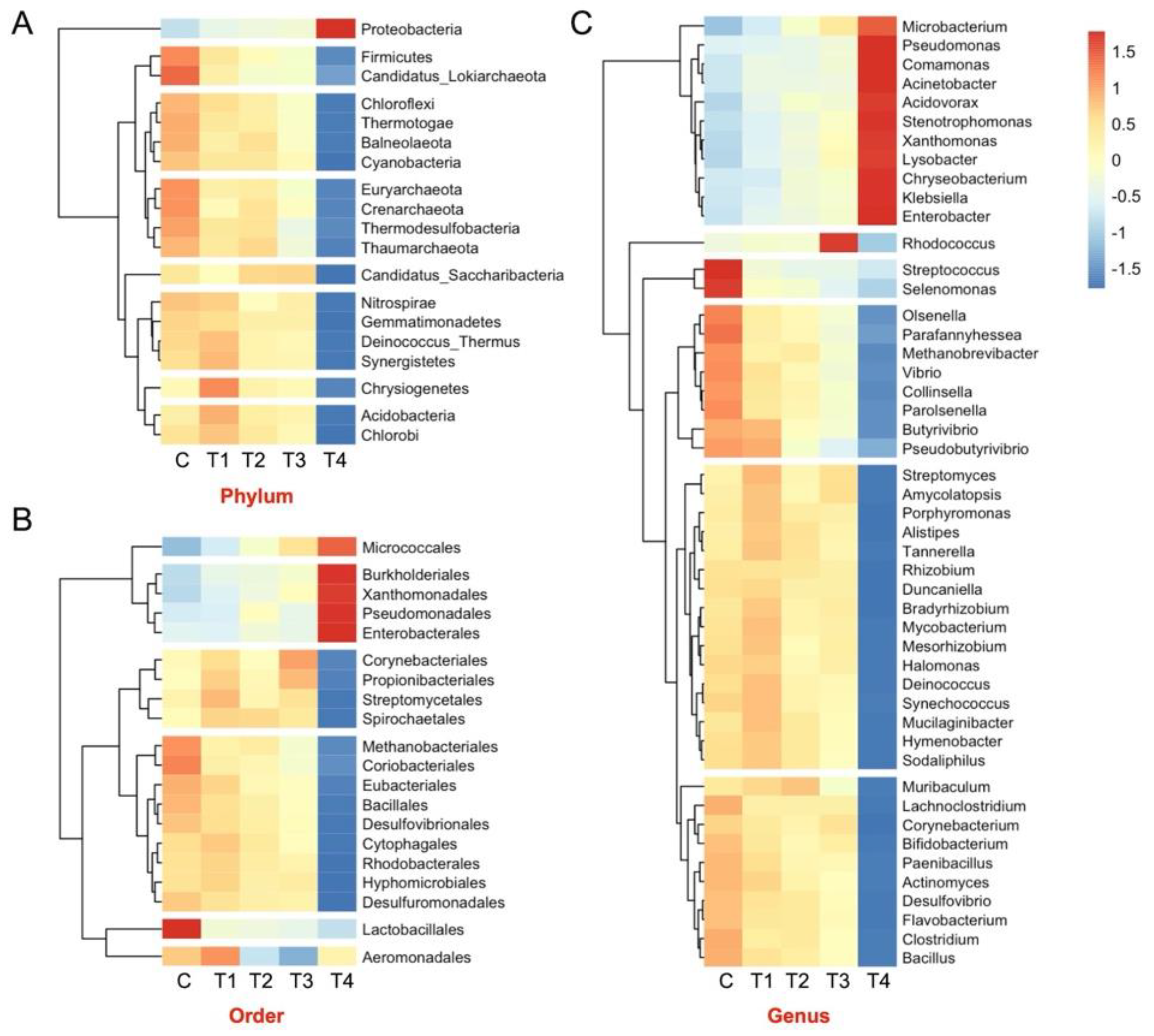
3.7. Metagenome Composition
3.8. CAZyme Profiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sonavale, K.P.; Shaikh, M.R.; Kadam, M.M.; Pokharkar, V.G. Livestock Sector in India: A Critical Analysis. Asian J. Agric. Ext. Econ. Sociol. 2020, 38, 51–62. [Google Scholar] [CrossRef]
- Gorti, R.K.; Sampath, K.T.; Suresh, K.P.; Giridhar, K.; Anandan, S. Modeling and Forecasting Livestock Fish Feed Resources: Requirement and Availability in India; National Institute of Animal Nutrition and Physiology: Bangalore, India, 2012. [Google Scholar]
- Makkar, H.P.S. Feed Demand Landscape and Implications of Food-Not Feed Strategy for Food Security and Climate Change. Animal 2018, 12, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.; Tiwari, M.R.; Acharaya, B. Effect of Inclusion of Dried Brewers Waste in Basal Diet of Dairy Animals on Milk Production and Its Parameter. Int. J. Res. Publ. Rev. 2023, 4, 265–269. [Google Scholar]
- Dlugokencky, E. NOAA/GML. Available online: https://Gml.Noaa.Gov/Ccgg/Trends_ch4/ (accessed on 11 June 2021).
- Yusuf, R.O.; Noor, Z.Z.; Abba, A.H.; Hassan, M.A.A.; Din, M.F.M. Methane Emission by Sectors: A Comprehensive Review of Emission Sources and Mitigation Methods. Renew. Sustain. Energy Rev. 2012, 16, 5059–5070. [Google Scholar] [CrossRef]
- EPA Global Methane Initiative. Importance of Methane. Available online: https://www.epa.gov/gmi/learn-about-global-methane-initiative (accessed on 19 September 2023).
- Nisbet, E.G.; Manning, M.R.; Dlugokencky, E.J.; Fisher, R.E.; Lowry, D.; Michel, S.E.; Myhre, C.L.; Platt, S.M.; Allen, G.; Bousquet, P.; et al. Very Strong Atmospheric Methane Growth in the 4 Years 2014–2017: Implications for the Paris Agreement. Glob. Biogeochem. Cycles 2019, 33, 318–342. [Google Scholar] [CrossRef]
- UNEP. United Nations Environment Programme/Climate and Clean Air Coalition. Global Methane Assessment: 2030 Baseline Report; UNEP: Nairobi, Kenya, 2022. [Google Scholar]
- UNEP. Pollution Action Note—Data You Need to Know; UNEP: Nairobi, Kenya, 2021; Available online: https://www.unep.org/interactives/air-pollution-note/ (accessed on 19 September 2023).
- Chang, J.; Peng, S.; Ciais, P.; Saunois, M.; Dangal, S.R.S.; Herrero, M.; Havlík, P.; Tian, H.; Bousquet, P. Revisiting Enteric Methane Emissions from Domestic Ruminants and Their Δ13CCH4 Source Signature. Nat. Commun. 2019, 10, 3420. [Google Scholar] [CrossRef]
- World Nuclear Association Heat Values of Various Fuels. Available online: https://world-nuclear.org/information-library/facts-and-figures/heat-values-of-various-fuels (accessed on 3 June 2024).
- Oyedeji, A.B.; Wu, J. Food-Based Uses of Brewers Spent Grains: Current Applications and Future Possibilities. Food Biosci. 2023, 54, 102774. [Google Scholar] [CrossRef]
- Tan, Y.X.; Mok, W.K.; Chen, W.N. In Vitro Evaluation of Enriched Brewers’ Spent Grains Using Bacillus Subtilis WX-17 as Potential Functional Food Ingredients. Appl. Biochem. Biotechnol. 2021, 193, 349–362. [Google Scholar] [CrossRef]
- Westendorf, M.L.; Wohlt, J.E. Brewing By-Products: Their Use as Animal Feeds. Vet. Clin. Food Anim. 2002, 18, 233–252. [Google Scholar] [CrossRef]
- Tišma, M.; Jurić, A.; Bucić-Kojić, A.; Panjičko, M.; Planinić, M. Biovalorization of Brewers’ Spent Grain for the Production of Laccase and Polyphenols. J. Inst. Brew. 2018, 124, 182–186. [Google Scholar] [CrossRef]
- Arvanitoyannis, I.S.; Tserkezou, P. Wheat, Barley and Oat Waste: A Comparative and Critical Presentation of Methods and Potential Uses of Treated Waste. Int. J. Food Sci. Technol. 2008, 43, 694–725. [Google Scholar] [CrossRef]
- AOAC Official Method of Analysis; Association of Officiating Analytical Chemists: Washington DC, USA, 2012.
- AOAC Official Method of Analysis; Association of Officiating Analytical Chemists: Washington DC, USA, 2005.
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy. Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- ICAR. Nutrient Requirements of Cattle and Buffaloes; Indian Council of Agricultural Research: New Delhi, India, 2013. [Google Scholar]
- Menke, K.H.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The Estimation of the Digestibility and Metabolizable Energy Content of Ruminant Feeding Stuffs from the Gas Production When They Are Incubated with Rumen Liquor In Vitro. J. Agric. Sci. 1979, 93, 217–222. [Google Scholar] [CrossRef]
- Filípek, J.; Dvořák, R. Determination of the Volatile Fatty Acid Content in the Rumen Liquid: Comparison of Gas Chromatography and Capillary Isotachophoresis. Acta Vet. Brno 2009, 78, 627–633. [Google Scholar] [CrossRef]
- Malik, P.K.; Trivedi, S.; Mohapatra, A.; Kolte, A.P.; Sejian, V.; Bhatta, R.; Rahman, H. Comparison of Enteric Methane Yield and Diversity of Ruminal Methanogens in Cattle and Buffaloes Fed on the Same Diet. PLoS ONE 2021, 16, e0256048. [Google Scholar] [CrossRef] [PubMed]
- Conway, E.J. Microdiffusion Analysis and Volumetric Error, 4th ed.; Crosby Lockwood and Son Ltd.: London, UK, 1957. [Google Scholar]
- Kamra, D.N.; Agarwal, N. Rumen Protozoa. In Techniques in Rumen Microbiology; Centre of Advance Studies in Animal Nutrition, Indian Veterinary Research Institute, Izatnagar: Bareilly, India, 2003; pp. 35–45. [Google Scholar]
- Hungate, R.E. The Rumen and Its Microbes; Academic Press Inc: New York, NY, USA, 1966. [Google Scholar]
- Yu, Z.; Morrison, M. Improved Extraction of PCR-Quality Community DNA from Digesta and Fecal Samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 8 November 2023).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved Metagenomic Analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef]
- Breitwieser, F.P.; Salzberg, S.L. Pavian: Interactive Analysis of Metagenomics Data for Microbiome Studies and Pathogen Identification. Bioinformatics 2020, 36, 1303–1304. [Google Scholar] [CrossRef]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using Microbiome Analyst for Comprehensive Statistical, Functional, and Meta-Analysis of Microbiome Data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Paulson, J.N.; Colin Stine, O.; Bravo, H.C.; Pop, M. Differential Abundance Analysis for Microbial Marker-Gene Surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. MEGAHIT: An Ultra-Fast Single-Node Solution for Large and Complex Metagenomics Assembly via Succinct de Bruijn Graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed]
- Gemayel, K.; Lomsadze, A.; Borodovsky, M. MetaGeneMark-2: Improved Gene Prediction in Metagenomes. BioRxiv 2022, 2022–2027. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-Hit: A Fast Program for Clustering and Comparing Large Sets of Protein or Nucleotide Sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Ge, Q.; Yan, Y.; Zhang, X.; Huang, L.; Yin, Y. DbCAN3: Automated Carbohydrate-Active Enzyme and Substrate Annotation. Nucleic Acids Res. 2023, 51, W115–W121. [Google Scholar] [CrossRef] [PubMed]
- Kolde, R. Pheatmap: Pretty Heatmaps. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 12 December 2023).
- Mussatto, S.I.; Dragone, G.; Roberto, I.C. Brewers’ Spent Grain: Generation, Characteristics and Potential Applications. J. Cereal Sci. 2006, 43, 1–14. [Google Scholar] [CrossRef]
- Agrawal, D.; Gopaliya, D.; Willoughby, N.; Khare, S.K.; Kumar, V. Recycling Potential of Brewer’s Spent Grains for Circular Biorefineries. Curr. Opin. Green. Sustain. Chem. 2023, 40, 100748. [Google Scholar] [CrossRef]
- Chetrariu, A.; Dabija, A. Brewer’s Spent Grains: Possibilities of Valorization, a Review. Appl. Sci. 2020, 10, 5619. [Google Scholar] [CrossRef]
- Lynch, K.M.; Steffen, E.J.; Arendt, E.K. Brewers’ Spent Grain: A Review with an Emphasis on Food and Health. J. Inst. Brew. 2016, 122, 553–568. [Google Scholar] [CrossRef]
- Bath, D.; Dunbar, J.; King, J.; Berry, S.; Leonard, R.; Olbrich, S. By-Products and Unusual Feedstuffs in Livestock Rations. Western Region Extension Publication-Cooperative Extension Service; USA, 1980. Available online: https://cesonoma.ucanr.edu/files/180221.pdf (accessed on 15 February 2024).
- Umunna, N.N.; Yusuf, A.A.; Aganga, A.A. Evaluation of Brewers’ Dried Grains and Palm Kernel Meal as Major Sources of Nitrogen for Growing Cattle. Trop. Anim. Prod. 1980, 5, 239–247. [Google Scholar]
- Kaur, V.I.; Saxena, P.K. Incorporation of Brewery Waste in Supplementary Feed and Its Impact on Growth in Some Carps. Bioresour. Technol. 2004, 91, 101–104. [Google Scholar] [CrossRef]
- Ogunjobi, A.A.; Mejeha, O.K.; Fagade, O.E. Protein enrichment of brewery spent grains using Aspergillus oryzae. AU J. Technol. 2011, 15, 53–56. [Google Scholar]
- Kebreab, E.; Strathe, A.; Fadel, J.; Moraes, L.; France, J. Impact of Dietary Manipulation on Nutrient Flows and Greenhouse Gas Emissions in Cattle. Rev. Bras. Zootec. 2010, 39, 458–464. [Google Scholar] [CrossRef]
- Malik, P.K.; Bhatta, R.; Soren, N.M.; Sejian, V.; Mech, A.; Prasad, K.S.; Prasad, C.S. Feed-Based Approaches in Enteric Methane Amelioration. In Livestock Production and Climate Change; Malik, P.K., Bhatta, R., Takahashi, J., Kohn, R.A., Prasad, C.S., Eds.; CABI: Wallingford, UK, 2015; pp. 336–359. [Google Scholar]
- Tavendale, M.H.; Meagher, L.P.; Pacheco, D.; Walker, N.; Attwood, G.T.; Sivakumaran, S. Methane Production from in Vitro Rumen Incubations with Lotus Pedunculatus and Medicago Sativa, and Effects of Extractable Condensed Tannin Fractions on Methanogenesis. Anim. Feed. Sci. Technol. 2005, 123–124, 403–419. [Google Scholar] [CrossRef]
- Bhatta, R.; Uyeno, Y.; Tajima, K.; Takenaka, A.; Yabumoto, Y.; Nonaka, I.; Enishi, O.; Kurihara, M. Difference in the Nature of Tannins on in Vitro Ruminal Methane and Volatile Fatty Acid Production and on Methanogenic Archaea and Protozoal Populations. J. Dairy. Sci. 2009, 92, 5512–5522. [Google Scholar] [CrossRef]
- Malik, P.K.; Kolte, A.P.; Baruah, L.; Saravanan, M.; Bakshi, B.; Bhatta, R. Enteric Methane Mitigation in Sheep through Leaves of Selected Tanniniferous Tropical Tree Species. Livest. Sci. 2017, 200, 29–34. [Google Scholar] [CrossRef]
- Malik, P.K.; Kolte, A.P.; Bakshi, B.; Baruah, L.; Dhali, A.; Bhatta, R. Effect of Tamarind Seed Husk Supplementation on Ruminal Methanogenesis, Methanogen Diversity and Fermentation Characteristics. Carbon. Manag. 2017, 8, 319–329. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Ungerfeld, E.M.; Eckard, R.J.; Wang, M. Review: Fifty Years of Research on Rumen Methanogenesis: Lessons Learned and Future Challenges for Mitigation. Animal 2020, 14, S2–S16. [Google Scholar] [CrossRef] [PubMed]
- Thirumalaisamy, G.; Malik, P.K.; Trivedi, S.; Kolte, A.P.; Bhatta, R. Effect of Long-Term Supplementation with Silkworm Pupae Oil on the Methane Yield, Ruminal Protozoa, and Archaea Community in Sheep. Front. Microbiol. 2022, 13, 780073. [Google Scholar] [CrossRef]
- Baruah, L.; Malik, P.K.; Kolte, A.P.; Goyal, P.; Dhali, A.; Bhatta, R. Rumen Methane Amelioration in Sheep Using Two Selected Tanniferous Phyto-Leaves. Carbon. Manag. 2019, 10, 299–308. [Google Scholar] [CrossRef]
- Poornachandra, K.T.; Malik, P.K.; Dhali, A.; Kolte, A.P.; Bhatta, R. Effect of Combined Supplementation of Tamarind Seed Husk and Soapnut on Enteric Methane Emission in Crossbred Cattle. Carbon. Manag. 2019, 10, 465–475. [Google Scholar] [CrossRef]
- Króliczewska, B.; Pecka-Kiełb, E.; Bujok, J. Strategies Used to Reduce Methane Emissions from Ruminants: Controversies and Issues. Agriculture 2023, 13, 602. [Google Scholar] [CrossRef]
- Hart, K.J.; Martin, P.G.; Foley, P.A.; Kenny, D.A.; Boland, T.M. Effect of Sward Dry Matter Digestibility on Methane Production, Ruminal Fermentation, and Microbial Populations of Zero-Grazed Beef Cattle. J. Anim. Sci. 2009, 87, 3342–3350. [Google Scholar] [CrossRef]
- Faccenda, A.; Zambom, M.A.; Avila, A.S.; Fernandes, T.; Stum, M.M.; Garcias, J.; Tinini, R.C.R.; Dias, A.L.G. Dried Brewers’ Grain as a Replacement for Soybean Meal on Nutrient Digestibility and Rumen Parameters of Cattle. S. Afr. J. Anim. Sci. 2018, 48, 1093–1098. [Google Scholar] [CrossRef]
- Babatunde, G.; Fetuga, B.; Oyenuga, V.; Ayoade, A. The Effects of Graded Levels of Brewers’ Dried Grains and Maize Cobs in the Diets of Pigs on Their Performance Characteristics and Carcass Quality. Niger. J. Anim. Prod. 1975, 2, 119–133. [Google Scholar] [CrossRef]
- Gonzalez Pereyra, M.L.; Rosa, C.A.R.; Dalcero, A.M.; Cavaglieri, L.R. Mycobiota and Mycotoxins in Malted Barley and Brewer’s Spent Grain from Argentinean Breweries. Lett. Appl. Microbiol. 2011, 53, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Cenkowski, S.; Izydorczyk, M. Thin-Layer Drying of Spent Grains in Superheated Steam. J. Food Eng. 2005, 67, 457–465. [Google Scholar] [CrossRef]
- Benedeti, P.D.B.; de Castro Silva, B.; Pacheco, M.V.C.; Serão, N.V.L.; Filho, I.C.; Lopes, M.M.; Marcondes, M.I.; Mantovani, H.C.; de Campos Valadares Filho, S.; Detmann, E.; et al. Effects of Grain Processing Methods on the Expression of Genes Involved in Volatile Fatty Acid Transport and PH Regulation, and Keratinization in Rumen Epithelium of Beef Cattle. PLoS ONE 2018, 13, 0198963. [Google Scholar] [CrossRef]
- Yan, L.; Zhang, B.; Shen, Z. Dietary Modulation of the Expression of Genes Involved in Short-Chain Fatty Acid Absorption in the Rumen Epithelium Is Related to Short-Chain Fatty Acid Concentration and PH in the Rumen of Goats. J. Dairy. Sci. 2014, 97, 5668–5675. [Google Scholar] [CrossRef]
- Sheng, P.; Ribeiro, G.; Wang, Y.; McAllister, T. Humic Substances Reduce Ruminal Methane Production and Increase the Efficiency of Microbial Protein Synthesis in Vitro. J. Sci. Food Agric. 2019, 99, 2152–2157. [Google Scholar] [CrossRef] [PubMed]
- Benchaar, C.; Hassanat, F.; Gervais, R.; Chouinard, P.Y.; Petit, H.V.; Massé, D.I. Methane Production, Digestion, Ruminal Fermentation, Nitrogen Balance, and Milk Production of Cows Fed Corn Silage- or Barley Silage-Based Diets. J. Dairy. Sci. 2014, 97, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Newbold, C.J.; Lassalas, B.; Jouany, J.P. The Importance of Methanogens Associated with Ciliate Protozoa in Ruminal Methane Production in Vitro. Lett. Appl. Microbiol. 1995, 21, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Tymensen, L.D.; Beauchemin, K.A.; McAllister, T.A. Structures of Free-Living and Protozoa-Associated Methanogen Communities in the Bovine Rumen Differ According to Comparative Analysis of 16S RRNA and McrA Genes. Microbiology 2012, 158, 1808–1817. [Google Scholar] [CrossRef] [PubMed]
- Ushida, K.; Tokura, M.; Takenaka, A.; Itabashi, H. Ciliate Protozoa and Ruminal Methanogenesis. In Rumen Microbes and Digestive Physiology in Ruminants; Onodera, R., Itabashi, H., Ushida, K., Yano, H., Sasaki, Y., Eds.; Japan Scientific Societies Press: Tokyo, Japan, 1997; pp. 209–230. ISBN 4-7622-0864-7. [Google Scholar]
- Belanche, A.; De La Fuente, G.; Newbold, C.J. Effect of Progressive Inoculation of Fauna-Free Sheep with Holotrich Protozoa and Total-Fauna on Rumen Fermentation, Microbial Diversity and Methane Emissions. FEMS Microbiol. Ecol. 2015, 91, fiu026. [Google Scholar] [CrossRef] [PubMed]
- Finlay, B.J.; Esteban, G.; Clarke, K.J.; Williams, A.G.; Embley, T.M.; Hirt, R.P. Some Rumen Ciliates Have Endosymbiotic Methanogens. FEMS Microbiol. Lett. 1994, 117, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Nathani, N.M.; Patel, A.K.; Mootapally, C.S.; Reddy, B.; Shah, S.V.; Lunagaria, P.M.; Kothari, R.K.; Joshi, C.G. Effect of Roughage on Rumen Microbiota Composition in the Efficient Feed Converter and Sturdy Indian Jaffrabadi Buffalo (Bubalus bubalis). BMC Genom. 2015, 16, 1116. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Gao, X.; Yang, Y.; Zou, C.; Yang, Y.; Lin, B. A Comparative Study on Rumen Ecology of Water Buffalo and Cattle Calves under Similar Feeding Regime. Vet. Med. Sci. 2020, 6, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.K.; Zhang, X.X.; Li, F.D.; Li, C.; Li, G.Z.; Zhang, D.Y.; Song, Q.Z.; Li, X.L.; Zhao, Y.; Wang, W.M. Characterization of the Rumen Microbiota and Its Relationship with Residual Feed Intake in Sheep. Animal 2021, 15, 100161. [Google Scholar] [CrossRef]
- Malik, P.K.; Trivedi, S.; Kolte, A.P.; Mohapatra, A.; Biswas, S.; Bhattar, A.V.K.; Bhatta, R.; Rahman, H. Comparative Analysis of Rumen Metagenome, Metatranscriptome, Fermentation and Methane Yield in Cattle and Buffaloes Fed on the Same Diet. Front. Microbiol. 2023, 14, 1266025. [Google Scholar] [CrossRef]
- Liu, J.H.; Bian, G.R.; Zhu, W.Y.; Mao, S.Y. High-Grain Feeding Causes Strong Shifts in Ruminal Epithelial Bacterial Community and Expression of Toll-like Receptor Genes in Goats. Front. Microbiol. 2015, 6, 167. [Google Scholar] [CrossRef] [PubMed]
- Jami, E.; Mizrahi, I. Composition and Similarity of Bovine Rumen Microbiota across Individual Animals. PLoS ONE 2012, 7, e33306. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Zeng, S.; Zhang, R.; Diao, Q.; Tu, Y. Effects of Dietary Energy Levels on Rumen Bacterial Community Composition in Holstein Heifers under the Same Forage to Concentrate Ratio Condition. BMC Microbiol. 2018, 18, 69. [Google Scholar] [CrossRef]
- Liu, C.; Meng, Q.; Chen, Y.; Xu, M.; Shen, M.; Gao, R.; Gan, S. Role of Age-Related Shifts in Rumen Bacteria and Methanogens in Methane Production in Cattle. Front. Microbiol. 2017, 8, 1563. [Google Scholar] [CrossRef]
- Malik, P.K.; Trivedi, S.; Kolte, A.P.; Mohapatra, A.; Bhatta, R.; Rahman, H. Effect of an Anti-Methanogenic Supplement on Enteric Methane Emission, Fermentation, and Whole Rumen Metagenome in Sheep. Front. Microbiol. 2022, 13, 1048288. [Google Scholar] [CrossRef]
- Malik, P.K.; Trivedi, S.; Kolte, A.P.; Mohapatra, A.; Biswas, S.; Bhattar, A.V.K.; Bhatta, R.; Rahman, H. Comparative Rumen Metagenome and CAZyme Profiles in Cattle and Buffaloes: Implications for Methane Yield and Rumen Fermentation on a Common Diet. Microorganisms 2023, 12, 47. [Google Scholar] [CrossRef]
- Moissl-Eichinger, C.; Probst, A.J.; Birarda, G.; Auerbach, A.; Koskinen, K.; Wolf, P.; Holman, H.Y.N. Human Age and Skin Physiology Shape Diversity and Abundance of Archaea on Skin. Sci. Rep. 2017, 7, 4039. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Forano, E.; Martin, C.; Newbold, C.J. Erratum: Microbial Ecosystem and Methanogenesis in Ruminants. Animal 2010, 4, 1024–1036. [Google Scholar] [CrossRef] [PubMed]
- Hungate, R.E. Hydrogen as an Intermediate in the Rumen Fermentation. Arch. Mikrobiol. 1967, 59, 158–164. [Google Scholar] [CrossRef]
- Kelly, W.J.; Mackie, R.I.; Attwood, G.T.; Janssen, P.H.; McAllister, T.A.; Leahy, S.C. Hydrogen and Formate Production and Utilisation in the Rumen and the Human Colon. Anim. Microbiome 2022, 4, 22. [Google Scholar] [CrossRef]
- Li, Y.; Lv, J.; Wang, J.; Zhou, S.; Zhang, G.; Wei, B.; Sun, Y.; Lan, Y.; Dou, X.; Zhang, Y. Changes in Carbohydrate Composition in Fermented Total Mixed Ration and Its Effects on in Vitro Methane Production and Microbiome. Front. Microbiol. 2021, 12, 738334. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Wang, Z.; He, X. Hydrogen Formation from Methane Rich Combustion under High Pressure and High Temperature Conditions. Int. J. Hydrog. Energy 2017, 42, 14301–14311. [Google Scholar] [CrossRef]
- Zhou, M.; McAllister, T.A.; Guan, L.L. Molecular Identification of Rumen Methanogens: Technologies, Advances and Prospects. Anim. Feed. Sci. Technol. 2011, 166–167, 76–86. [Google Scholar] [CrossRef]
- Danielsson, R.; Dicksved, J.; Sun, L.; Gonda, H.; Müller, B.; Schnürer, A.; Bertilsson, J. Methane Production in Dairy Cows Correlates with Rumen Methanogenic and Bacterial Community Structure. Front. Microbiol. 2017, 8, 226. [Google Scholar] [CrossRef] [PubMed]
- Danielsson, R.; Schnürer, A.; Arthurson, V.; Bertilsson, J. Methanogenic Population and CH4 Production in Swedish Dairy Cows Fed Different Levels of Forage. Appl. Environ. Microbiol. 2012, 78, 6172–6179. [Google Scholar] [CrossRef] [PubMed]
- Pitta, D.W.; Melgar, A.; Hristov, A.N.; Indugu, N.; Narayan, K.S.; Pappalardo, C.; Hennessy, M.L.; Vecchiarelli, B.; Kaplan-Shabtai, V.; Kindermann, M.; et al. Temporal Changes in Total and Metabolically Active Ruminal Methanogens in Dairy Cows Supplemented with 3-Nitrooxypropanol. J. Dairy. Sci. 2021, 104, 8721–8735. [Google Scholar] [CrossRef] [PubMed]
- Thauer, R.K.; Kaster, A.K.; Seedorf, H.; Buckel, W.; Hedderich, R. Methanogenic Archaea: Ecologically Relevant Differences in Energy Conservation. Nat. Rev. Microbiol. 2008, 6, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Hinsu, A.T.; Tulsani, N.J.; Panchal, K.J.; Pandit, R.J.; Jyotsana, B.; Dafale, N.A.; Patil, N.V.; Purohit, H.J.; Joshi, C.G.; Jakhesara, S.J. Characterizing Rumen Microbiota and CAZyme Profile of Indian Dromedary Camel (Camelus Dromedarius) in Response to Different Roughages. Sci. Rep. 2021, 11, 9400. [Google Scholar] [CrossRef]
- Kataeva, I.A.; Seidel, R.D.; Shah, A.; West, L.T.; Li, X.L.; Ljungdahl, L.G. The Fibronectin Type 3-like Repeat from the Clostridium Thermocellum Cellobiohydrolase CbHa Promotes Hydrolysis of Cellulose by Modifying Its Surface. Appl. Environ. Microbiol. 2002, 68, 4292–4300. [Google Scholar] [CrossRef]
- Stewart, R.D.; Auffret, M.D.; Warr, A.; Wiser, A.H.; Press, M.O.; Langford, K.W.; Liachko, I.; Snelling, T.J.; Dewhurst, R.J.; Walker, A.W.; et al. Assembly of 913 Microbial Genomes from Metagenomic Sequencing of the Cow Rumen. Nat. Commun. 2018, 9, 870. [Google Scholar] [CrossRef]
- Lehninger, A.L. Carbohydrates and Glycobiology. In Lehninger Principles of Biochemistry; Lehninger, A., Nelson, D., Cox, M., Eds.; Macmillan: New York, NY, USA, 2005; pp. 305–325. [Google Scholar]
- Lairson, L.; Henrissat, B.; Davies, G.; Withers, S. Glycosyltransferases: Structures, Functions, and Mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, G.; Xu, H.; Xin, H.; Zhang, Y. Metagenomic Analyses of Microbial and Carbohydrate-Active Enzymes in the Rumen of Holstein Cows Fed Different Forage-to-Concentrate Ratios. Front. Microbiol. 2019, 10, 649. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, R.; Malik, P.K.; Kolte, A.P.; Suresh, K.P. Assessment of Enteric Methane Emission from Indian Livestock: A New Approach. In Proceedings of the 7th Pan Commonwealth Veterinary Conference, Bangalore, India, 3–7 March 2019; Sejian, V., Isloor, S., Rahman, S.A., Bhatta, R., Eds.; Commonwealth Veterinary Association(Asia): Bengaluru, Indian, 2019; pp. 101–103. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attributes | Treatment | SEM | p | ||||
|---|---|---|---|---|---|---|---|
| C | T1 | T2 | T3 | T4 | |||
| Total gas (mL/g DM) | 312 a | 271 b | 242 c | 208 d | 123 e | 11.05 | <0.0001 |
| CH4 (mL/g DM) | 49.8 a | 40.8 a | 31.4 b | 23.9 b | 10.1 c | 4.92 | <0.0001 |
| CH4 (mL/g OM) | 52.7 a | 43.0 a | 32.9 b | 25.0 b | 10.5 c | 7.29 | <0.0001 |
| IVDMD (%) | 83.7 a | 79.3 a | 72.9 b | 64.7 c | 47.3 d | 2.56 | <0.0001 |
| OMD (%) | 75.3 a | 71.6 b | 66.4 c | 58.5 d | 41.6 e | 0.855 | <0.0001 |
| Total gas (mL/g dig. DM) | 373 a | 343 b | 333 b | 321 b | 260 c | 15.28 | <0.0001 |
| CH4 (mL/g dig. DM) | 59.5 a | 51.5 ab | 43.1 b | 36.8 b | 21.2 c | 7.83 | <0.0001 |
| CH4 (mL/g dig. OM) | 70.1 a | 60.1 ab | 49.6 bc | 42.6 c | 25.2 d | 7.67 | <0.0001 |
| Attributes | Treatment | SEM | p | ||||
|---|---|---|---|---|---|---|---|
| C | T1 | T2 | T3 | T4 | |||
| VFA | |||||||
| Acetate | 67.6 | 67.9 | 66.4 | 67.9 | 67.1 | 2.01 | 0.829 |
| Propionate | 23.2 | 23.1 | 20.5 | 22.1 | 18.9 | 3.84 | 0.502 |
| Butyrate | 4.57 | 5.49 | 6.46 | 5.04 | 6.08 | 1.00 | 0.104 |
| Valerate | 2.07 a | 2.29 ab | 2.44 ab | 3.02 c | 3.29 c | 0.18 | <0.0001 |
| Iso-valerate | 2.67 | 1.67 | 2.43 | 0.18 | 0.49 | 1.47 | 0.102 |
| A/P ratio | 2.93 | 2.95 | 3.33 | 3.31 | 3.55 | 0.62 | 0.565 |
| Ammonia-N | 16.1 a | 18.9 ab | 19.2 ab | 21.0 b | 22.4 b | 1.06 | 0.005 |
| Protozoa | |||||||
| Total (×107 cells/mL fluid) | 8.27 | 8.24 | 8.21 | 8.07 | 8.01 | 0.050 | 0.604 |
| Entodiniomorphs (×107 cells/mL fluid) | 8.25 | 8.22 | 8.19 | 8.06 | 8.00 | 0.048 | 0.647 |
| Holotrichs (×106 cells/mL fluid) | 0.213 a | 0.214 a | 0.213 a | 0.153 b | 0.137 b | 0.016 | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malik, P.K.; Trivedi, S.; Mohapatra, A.; Kolte, A.P.; Mech, A.; Victor, T.; Ahasic, E.; Bhatta, R. Oat Brewery Waste Decreased Methane Production and Alters Rumen Fermentation, Microbiota Composition, and CAZymes Profiles. Microorganisms 2024, 12, 1475. https://doi.org/10.3390/microorganisms12071475
Malik PK, Trivedi S, Mohapatra A, Kolte AP, Mech A, Victor T, Ahasic E, Bhatta R. Oat Brewery Waste Decreased Methane Production and Alters Rumen Fermentation, Microbiota Composition, and CAZymes Profiles. Microorganisms. 2024; 12(7):1475. https://doi.org/10.3390/microorganisms12071475
Chicago/Turabian StyleMalik, Pradeep Kumar, Shraddha Trivedi, Archit Mohapatra, Atul Purshottam Kolte, Anjumoni Mech, Tsuma Victor, Elena Ahasic, and Raghavendra Bhatta. 2024. "Oat Brewery Waste Decreased Methane Production and Alters Rumen Fermentation, Microbiota Composition, and CAZymes Profiles" Microorganisms 12, no. 7: 1475. https://doi.org/10.3390/microorganisms12071475
APA StyleMalik, P. K., Trivedi, S., Mohapatra, A., Kolte, A. P., Mech, A., Victor, T., Ahasic, E., & Bhatta, R. (2024). Oat Brewery Waste Decreased Methane Production and Alters Rumen Fermentation, Microbiota Composition, and CAZymes Profiles. Microorganisms, 12(7), 1475. https://doi.org/10.3390/microorganisms12071475