
Exosomal miR-7-25207 Increases Subgroup J Avian Leukosis Virus Titers by Targeting the Akt-CyclinQ1 and PRC1-YAF2 Dual Pathways
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Isolation and Purification of Exosome
2.3. Electron Microscopy and Nanoparticle Tracking Analysis (NTA)
2.4. Exosome Labeling and Uptake
2.5. Infection Assay and Immunofluorescence Assay
2.6. Total RNA Extraction, RT-PCR, and qRT-PCR Assay
2.7. Western Blot Analysis
2.8. miRNAomics Assay
2.9. miR-7-25207 Overexpression and Interference
2.10. ELISA Assay
2.11. Data Availability
2.12. Statistical Analysis
3. Results
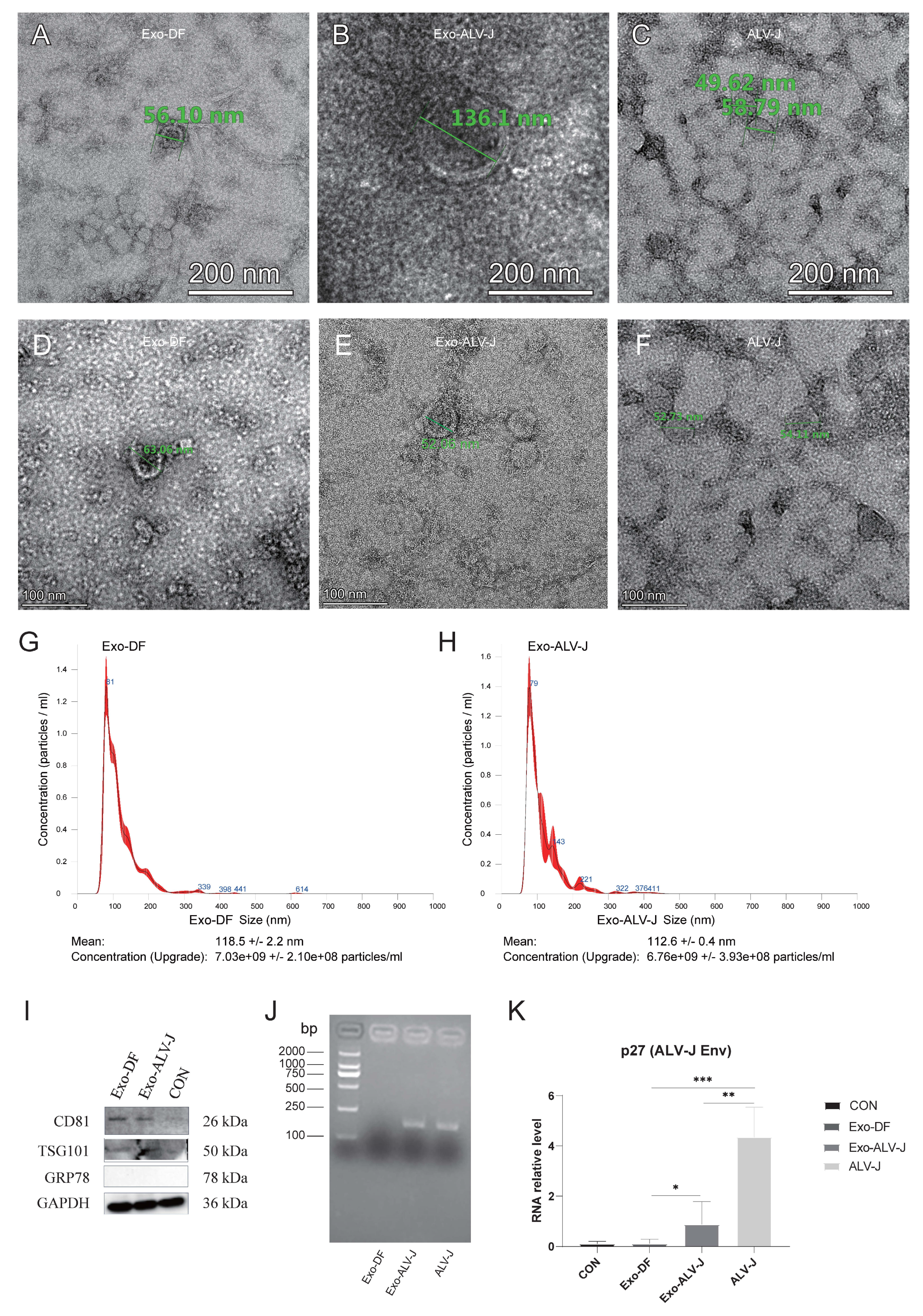
3.1. Identification of Exosome Morphology and Protein Expression
3.2. Exo-ALV-J Encapsulates ALV-J and Causes Productive Infection of ALV-J
3.3. ALV-J Activates the Expression of miR-7-25207
3.4. miR-7-25207 Increases the Titers of ALV-J
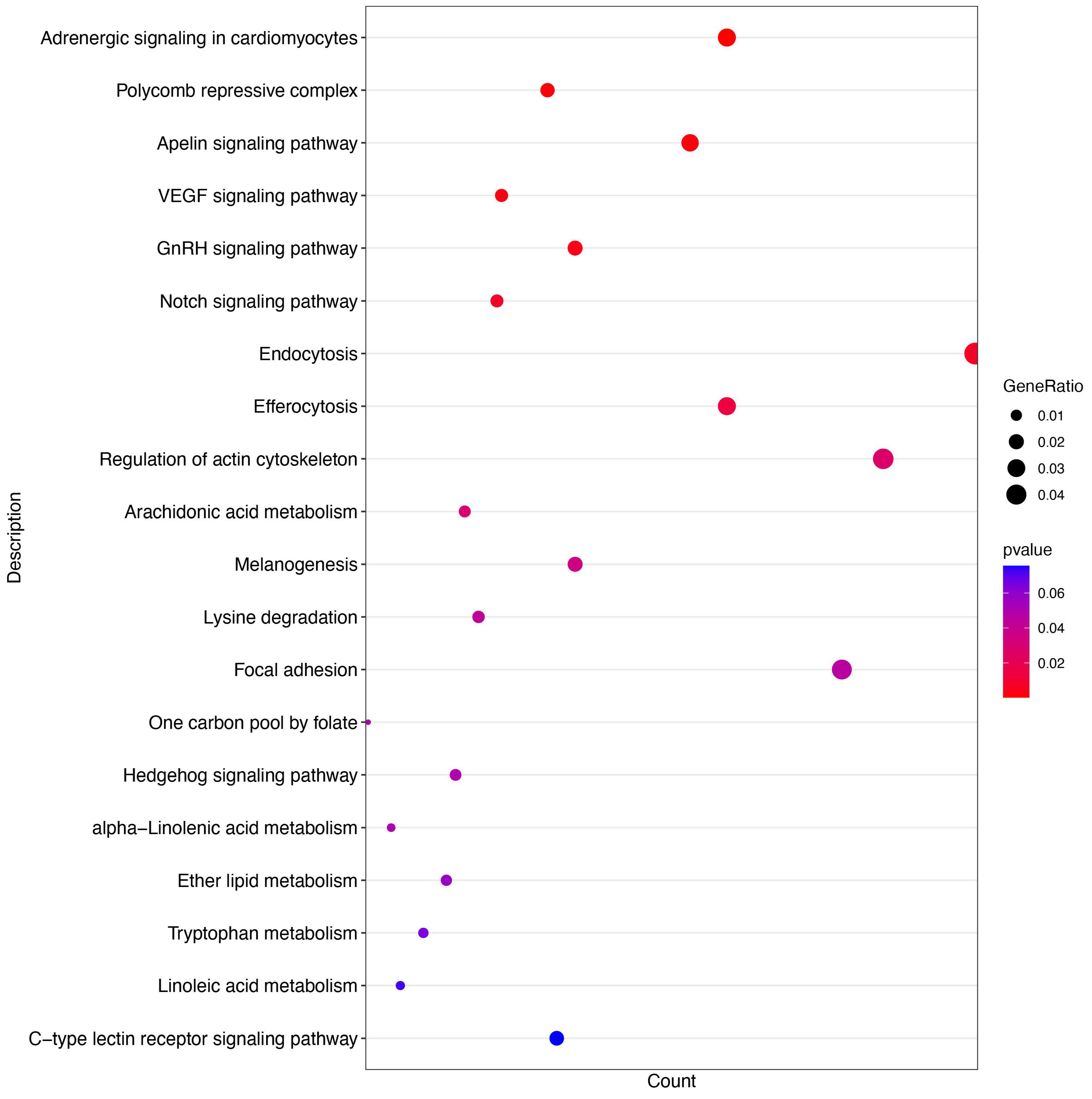
3.5. miR-7-25207 Targets the Akt-CyclinQ1 and PRC1-YAF2/CBX7 Dual Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Payne, L.N.; Venugopal, K. Neoplastic diseases: Marek’s disease, avian leukosis and reticuloendotheliosis. Rev. Sci. Technol. 2000, 19, 544–564. [Google Scholar] [CrossRef]
- Liao, L.; Chen, W.; Zhang, X.; Zhang, H.; Li, A.; Yan, Y.; Xie, Z.; Li, H.; Lin, W.; Ma, J.; et al. Semen extracellular vesicles mediate vertical transmission of subgroup J avian leukosis virus. Virol. Sin. 2022, 37, 284–2944. [Google Scholar] [CrossRef] [PubMed]
- Fandino, S.; Gomez-Lucia, E.; Benitez, L.; Domenech, A. Avian Leukosis: Will We Be Able to Get Rid of It? Animals 2023, 13, 2358. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, C.; Deng, Y.; Pan, M.; Mo, G.; Liao, Z.; Zhang, X.; Zhang, D.; Li, H. PMAIP1 promotes J subgroup avian leukosis virus replication by regulating mitochondrial function. Poult. Sci. 2024, 103, 103617. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Xue, J.; Zhang, Y.; Wang, G.; Feng, Y.; Hu, L.; Shang, Y.; Cheng, Z. Outbreak of myelocytomatosis caused by mutational avian leukosis virus subgroup J in China, 2018. Transbound. Emerg. Dis. 2019, 66, 622–626. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yao, X.; Li, C.; Zhang, J.; Xie, Q.; Wang, W.; Lu, H.; Fu, H.; Li, L.; Xie, J.; et al. Gp37 Regulates the Pathogenesis of Avian Leukosis Virus Subgroup J via Its C Terminus. J. Virol. 2020, 94, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Yang, J.; Bi, X.; Yang, Q.; Zhou, D.; Zhang, S.; Ding, L.; Wang, K.; Hua, S.; Cheng, Z. Molecular characteristics and pathogenicity of a Tibet-origin mutant avian leukosis virus subgroup J isolated from Tibetan chickens in China. Infect. Genet. Evol. 2023, 109, 105415. [Google Scholar] [CrossRef] [PubMed]
- Fotouh, A.; Shosha, E.; Zanaty, A.; Darwesh, M. Immunopathological investigation and genetic evolution of Avian leukosis virus Subgroup-J associated with myelocytomatosis in broiler flocks in Egypt. Virol. J. 2024, 21, 83. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, Z.; Zhuang, P.; Zhao, X.; Cheng, Z. Exosomes carring gag/env of ALV-J possess negative effect on immunocytes. Microb. Pathog. 2017, 112, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Batagov, A.O.; Kuznetsov, V.A.; Kurochkin, I.V. Identification of nucleotide patterns enriched in secreted RNAs as putative cis-acting elements targeting them to exosome nano-vesicles. BMC Genom. 2011, 12 (Suppl. S3), S18. [Google Scholar] [CrossRef] [PubMed]
- Cha, D.; Franklin, J.; Dou, Y.; Liu, Q.; Higginbotham, J.; Beckler, M.; Weaver, A.; Vickers, K.; Prasad, N.; Levy, S.; et al. KRAS-dependent sorting of miRNA to exosomes. Elife 2015, 4, e07197. [Google Scholar] [CrossRef] [PubMed]
- Ostenfeld, M.; Jeppesen, D.; Laurberg, J.; Boysen, A.; Bramsen, J.; Primdal-Bengtson, B.; Hendrix, A.; Lamy, P.; Dagnaes-Hansen, F.; Rasmussen, M.; et al. Cellular disposal of miR23b by RAB27-dependent exosome release is linked to acquisition of metastatic properties. Cancer Res. 2014, 74, 5758–5771. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Malinowska, K.; Zöller, M. Exosomal tumor microRNA modulates premetastatic organ cells. Neoplasia 2013, 15, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Xu, Y.; Liu, N.; Lv, D.; Chen, Y.; Liu, Z.; Jin, X.; Xiao, M.; Lavillette, D.; Zhong, J.; et al. Extracellular vesicles from Zika virus-infected cells display viral E protein that binds ZIKV-neutralizing antibodies to prevent infection enhancement. EMBO J. 2023, 42, e112096. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, L.; Liang, W.; Liu, S.; Deng, W.; Liu, Y.; Liu, Y.; Song, M.; Guo, K.; Zhang, Y. Extracellular vesicles originating from autophagy mediate an antibody-resistant spread of classical swine fever virus in cell culture. Autophagy 2022, 18, 1433–1449. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Fang, L.; Zhao, F.; Wang, D.; Xiao, S. Exosomes Mediate Intercellular Transmission of Porcine Reproductive and Respiratory Syndrome Virus. J. Virol. 2018, 92, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.; Tran, N. miRNA interplay: Mechanisms and consequences in cancer. Dis. Model Mech. 2021, 14, dmm047662. [Google Scholar] [CrossRef] [PubMed]
- Alkan, A.H.; Akgul, B. Endogenous miRNA Sponges. Methods Mol. Biol. 2022, 2257, 91–104. [Google Scholar] [PubMed]
- Sharma, R.; Tiwari, A.; McGeachie, M.J. Recent miRNA Research in Asthma. Curr. Allergy Asthma Rep. 2022, 22, 231–258. [Google Scholar] [CrossRef] [PubMed]
- Muller, L. Exosomes: Nanodust? HNO 2020, 68 (Suppl. S1), 56–59. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Kang, M.; Jeyaraj, M.; Oasim, M.; Kim, J. Correction: Gurunathan, S. et al. Review of the Isolation, Characterization, Biological Function, and Multifarious Therapeutic Approaches of Exosomes. Cells 2019, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Tao, J.; Li, Y.; Feng, Y.; Ju, H.; Wang, Z.; Ding, L. Quantitative Localized Analysis Reveals Distinct Exosomal Protein-Specific Glycosignatures: Implications in Cancer Cell Subtyping, Exosome Biogenesis, and Function. J. Am. Chem. Soc. 2020, 142, 7404–7412. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.K.; Kim, S.; Kim, S. Shedding Light on the Cell Biology of Platelet-Derived Extracellular Vesicles and Their Biomedical Applications. Life 2023, 13, 1403. [Google Scholar] [CrossRef] [PubMed]
- Crenshaw, B.J.; Gu, L.; Sims, B.; Matthews, Q. Exosome Biogenesis and Biological Function in Response to Viral Infections. Open Virol. J. 2018, 12, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Nazarenko, I.; Rana, S.; Baumann, A.; McAlear, J.; Hellwig, A.; Trendelenburg, M.; Lochnit, G.; Preissner, K.; Zoller, M. Cell surface tetraspanin Tspan8 contributes to molecular pathways of exosome-induced endothelial cell activation. Cancer Res. 2010, 70, 1668–1678. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Yue, S.; Stadel, D.; Zoller, M. Toward tailored exosomes: The exosomal tetraspanin web contributes to target cell selection. Int. J. Biochem. Cell Biol. 2012, 44, 1574–1584. [Google Scholar] [CrossRef] [PubMed]
- Slonchak, A.; Clarke, B.; Mackenzie, J.; Amarilla, A.; Setoh, Y.; Khromykh, A. West Nile virus infection and interferon alpha treatment alter the spectrum and the levels of coding and noncoding host RNAs secreted in extracellular vesicles. BMC Genom. 2019, 20, 474. [Google Scholar] [CrossRef] [PubMed]
- Nahand, J.; Bokharaei-Salim, F.; Karimzadeh, M.; Moghoofei, M.; Karampoor, S.; Mirzaei, H.; Tabibzadeh, A.; Jafari, A.; Ghaderi, A.; Asemi, Z.; et al. MicroRNAs and exosomes: Key players in HIV pathogenesis. HIV Med. 2020, 21, 246–278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yu, D. Exosomes in cancer development, metastasis, and immunity. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Beckler, M.; Higginbotham, J.; Franklin, J.; Ham, A.; Halvey, P.; Imasuen, I.; Whitwell, C.; Li, M.; Liebler, D.; Coffey, R. Proteomic analysis of exosomes from mutant KRAS colon cancer cells identifies intercellular transfer of mutant KRAS. Mol. Cell Proteom. 2013, 12, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Pavlyukov, M.; Yu, H.; Bastola, S.; Minata, M.; Shender, V.; Lee, Y.; Zhang, S.; Wang, J.; Komarova, S.; Wang, J.; et al. Apoptotic Cell-Derived Extracellular Vesicles Promote Malignancy of Glioblastoma via Intercellular Transfer of Splicing Factors. Cancer Cell 2018, 34, 119–135.e10. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, C.; Chan, L.; Wei, Y.; Hsu, J.; Xia, W.; Cha, J.; Hou, J.; Hsu, J.; Sun, L.; et al. Exosomal PD-L1 harbors active defense function to suppress T cell killing of breast cancer cells and promote tumor growth. Cell Res. 2018, 28, 862–864. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Zhang, X.; Xue, J.; Yao, Y.; Zhou, D.; Cheng, Z. TMT-based proteomic analysis reveals integrins involved in the synergistic infection of reticuloendotheliosis virus and avian leukosis virus subgroup J. BMC Vet. Res. 2022, 18, 131. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Zhou, D.; Zhou, J.; Xue, J.; Cheng, Z. Marek’s Disease Virus and Reticuloendotheliosis Virus Coinfection Enhances Viral Replication and Alters Cellular Protein Profiles. Front. Vet. Sci. 2022, 9, 854007. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Zhou, D.; Zhou, J.; Du, X.; Zhang, X.; Liu, X.; Ding, L.; Cheng, Z. miR-155 facilitates the synergistic replication between avian leukosis virus subgroup J and reticuloendotheliosis virus by targeting a dual pathway. J. Virol. 2023, 97, e0093723. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Feng, Y.; Wang, J.; Liang, Y.; Zou, W. Long non-coding RNA SNHG15 in various cancers: A meta and bioinformatic analysis. BMC Cancer 2020, 20, 1156. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Yin, Y.; Yang, H.; Rui, W.; Ri, K.; Rui, D.; Liu, J.; Yu, Z. Comprehensive analysis of lncRNA-mRNA co-expression networks in HPV-driven cervical cancer reveals the pivotal function of LINC00511-PGK1 in tumorigenesis. Comput. Biol. Med. 2023, 159, 106943. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Jike, Y.; Liu, K.; Gan, F.; Zhang, K.; Xie, M.; Zhang, J.; Chen, C.; Zou, X.; Jiang, X.; et al. Exosome-mediated miR-144-3p promotes ferroptosis to inhibit osteosarcoma proliferation, migration, and invasion through regulating ZEB1. Mol. Cancer 2023, 22, 113. [Google Scholar] [CrossRef] [PubMed]
- Murayama, N.; Okamoto, K.; Nakagawa, T.; Miyoshi, J.; Nishida, K.; Kawaguchi, T.; Kagemoto, K.; Kitamura, S.; Ma, B.; Miyamoto, H.; et al. miR-144-3p/miR-451a promotes lymphovascular invasion through repression of PTEN/p19 in rectal neuroendocrine tumors. J. Gastroenterol. Hepatol. 2022, 37, 919–927. [Google Scholar] [CrossRef]
- Cao, H.; Gu, M.; Sun, Z.; Chen, Z. miR-144-3p Contributes to the Development of Thyroid Tumors Through the PTEN/PI3K/AKT Pathway. Cancer Manag. Res. 2020, 12, 9845–9855. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, Y.; Jiang, X.; Hang, Y.; Wang, H.; Liu, H.; Chen, Z.; Xiao, Y. Inhibition of miR-144-3p exacerbates non-small cell lung cancer progression by targeting CEP55. Acta Biochim. Biophys. Sin. 2021, 53, 1398–1407. [Google Scholar] [CrossRef] [PubMed]
- Fang, G.; Zhang, C.; Liu, Z.; Peng, Z.; Tang, M.; Xue, Q. MiR-144-3p inhibits the proliferation and metastasis of lung cancer A549 cells via targeting HGF. J. Cardiothorac. Surg. 2022, 17, 117. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.Y.; Liu, H.; Zhang, Z. miR-144-3p increases radiosensibility of gastric cancer cells by targeting inhibition of ZEB1. Clin. Transl. Oncol. 2021, 23, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Luo, D.; Gu, X.; Pervez, A.; Huang, X.; Li, S. MiR-144-3p targets STC1 to activate PI3K/AKT pathway to induce cell apoptosis and cell cycle arrest in selenium deficiency broilers. J. Inorg. Biochem. 2022, 226, 111665. [Google Scholar]
- Flora, P.; Dalal, G.; Cohen, I.; Ezhkova, E. Polycomb Repressive Complex(es) and Their Role in Adult Stem Cells. Genes 2021, 12, 1485. [Google Scholar] [CrossRef]
- Barrasa, J.; Kahn, T.; Lundkvist, M.; Schwartz, Y. DNA elements tether canonical Polycomb Repressive Complex 1 to human genes. Nucleic Acids Res. 2023, 51, 11613–11633. [Google Scholar] [CrossRef]
- Shimura, T.; Kakuda, S.; Ochiai, Y.; Kuwahara, Y.; Takai, Y.; Fukumoto, M. Targeting the AKT/GSK3beta/cyclin D1/Cdk4 survival signaling pathway for eradication of tumor radioresistance acquired by fractionated radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2011, 80, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Shimura, T. Targeting the AKT/cyclin D1 pathway to overcome intrinsic and acquired radioresistance of tumors for effective radiotherapy. Int. J. Radiat. Biol. 2017, 93, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Perez-Nunez, R.; Chamorro, A.; Gonzalez, M.; Contreras, P.; Artigas, R.; Corvalan, A.; Zundert, B.; Reyes, C.; Moya, P.; Avalos, A.; et al. Protein kinase B (AKT) upregulation and Thy-1-alpha(v)beta(3) integrin-induced phosphorylation of Connexin43 by activated AKT in astrogliosis. J. Neuroinflamm. 2023, 20, 5. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name a | Primer Sequence (5′-3′) |
|---|---|
| ALV-J-p27 | F: ACAAGACTGGCTGATACGGT |
| R: AAGCGTCCATCCATAAGGCA | |
| GAPDH | F: CCCCCATGTTTGTGATGGGT |
| R: TGATGGCATGGACAGTGGTC | |
| RBBP7 | F: CCTTCTTGTACGACCTGGTGATGAC |
| R: GCCAGTGTAGAGCGTAATCCTTTCC | |
| YAF2 | F: GGACGAGGGCTACTGGGACTG |
| R: ACGAATTGCTGAGGAACCTGCTG | |
| Egr1 | F: GACCACTTGACCACGCACATCC |
| R: GAGGAAGAAGTTGCTGAGACCGAAG | |
| CyclinD1 | F: TCCAGTCTACGCCAGGCACAG |
| R: TGTTCACATCTCGCACATCAGTGG | |
| EZH | F: GGGAGGGCTGAACAACGAAACC |
| R: CGGCTGTGCTGCTGCTTAGG | |
| CBX7 | F: AGGTTTGGAGGAGGAGACAGTAGC |
| R: CTTCTTCCTGGACAGTCGCAAGTAC | |
| Akt | F: GTAGCGATAGTGAAGGAAGGATGGC |
| R: CGTCTTGCGGTCGTTCCTTGTAG | |
| PRC1 | F: CCGCCAGATTGAGACAGAGATGATG |
| R: TGGAGAGGGAAGTGCCGTTGAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, X.; Liu, T.; Tang, S.; Dong, X.; Li, Y.; Liao, L.; Chen, S.; Chen, L.; Kong, J.; Dai, Z.; et al. Exosomal miR-7-25207 Increases Subgroup J Avian Leukosis Virus Titers by Targeting the Akt-CyclinQ1 and PRC1-YAF2 Dual Pathways. Microorganisms 2024, 12, 1495. https://doi.org/10.3390/microorganisms12071495
Zeng X, Liu T, Tang S, Dong X, Li Y, Liao L, Chen S, Chen L, Kong J, Dai Z, et al. Exosomal miR-7-25207 Increases Subgroup J Avian Leukosis Virus Titers by Targeting the Akt-CyclinQ1 and PRC1-YAF2 Dual Pathways. Microorganisms. 2024; 12(7):1495. https://doi.org/10.3390/microorganisms12071495
Chicago/Turabian StyleZeng, Xiaona, Tongfei Liu, Shengqiu Tang, Xiaoying Dong, Yajuan Li, Liqin Liao, Sheng Chen, Liyi Chen, Jie Kong, Zhenkai Dai, and et al. 2024. "Exosomal miR-7-25207 Increases Subgroup J Avian Leukosis Virus Titers by Targeting the Akt-CyclinQ1 and PRC1-YAF2 Dual Pathways" Microorganisms 12, no. 7: 1495. https://doi.org/10.3390/microorganisms12071495
APA StyleZeng, X., Liu, T., Tang, S., Dong, X., Li, Y., Liao, L., Chen, S., Chen, L., Kong, J., Dai, Z., Feng, K., Wong, Y.-H., & Xie, Q. (2024). Exosomal miR-7-25207 Increases Subgroup J Avian Leukosis Virus Titers by Targeting the Akt-CyclinQ1 and PRC1-YAF2 Dual Pathways. Microorganisms, 12(7), 1495. https://doi.org/10.3390/microorganisms12071495