
The Repurposing of FDA-Approved Drugs as FtsZ Inhibitors against Mycobacterium tuberculosis: An In Silico and In Vitro Study
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Molecular Docking
2.2. Cell Culture
2.3. Microplate Alamar Blue Assay (MABA)
2.4. Evaluation of Growth Inhibition of Mtb
2.5. Cytotoxicity Assay
2.6. CFUs/mL Assay
2.7. Statistical Analysis
3. Results
3.1. Molecule Selection
3.2. Paroxetine and Nebivolol Show Anti-Mycobacterial Tuberculosis Activity
3.3. Paroxetine and Nebivolol Reduce CFU/mL Counts in Macrophages



4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2023; World Health Organization: Geneva, Switzerland, 2023. [Google Scholar]
- World Health Organization. The End TB Strategy. Available online: https://www.who.int/teams/global-tuberculosis-programme/the-end-tb-strategy (accessed on 4 June 2024).
- Liu, Y.; Tong, Z.; Shi, J.; Li, R.; Upton, M.; Wang, Z. Drug repurposing for next-generation combination therapies against multidrug-resistant bacteria. Theranostics 2021, 11, 4910–4928. [Google Scholar] [CrossRef]
- Sharma, K.; Ahmed, F.; Sharma, T.; Grover, A.; Agarwal, M.; Grover, S. Potential Repurposed Drug Candidates for Tuberculosis Treatment: Progress and Update of Drugs Identified in Over a Decade. ACS Omega 2023, 8, 17362–17380. [Google Scholar] [CrossRef]
- Conradie, F.; Bagdasaryan, T.R.; Borisov, S.; Howell, P.; Mikiashvili, L.; Ngubane, N.; Samoilova, A.; Skornykova, S.; Tudor, E.; Variava, E.; et al. Bedaquiline-Pretomanid-Linezolid Regimens for Drug-Resistant Tuberculosis. N. Engl. J. Med. 2022, 387, 810–823. [Google Scholar] [CrossRef]
- Stadler, J.A.M.; Maartens, G.; Meintjes, G.; Wasserman, S. Clofazimine for the treatment of tuberculosis. Front. Pharmacol. 2023, 14, 1100488. [Google Scholar] [CrossRef]
- Margolin, W. FtsZ and the division of prokaryotic cells and organelles. Nat. Rev. Mol. Cell Biol. 2005, 6, 862–871. [Google Scholar] [CrossRef]
- Nazir, A.; Harinarayanan, R. Inactivation of Cell Division Protein FtsZ by SulA Makes Lon Indispensable for the Viability of a ppGpp0 Strain of Escherichia coli. J. Bacteriol. 2015, 198, 688–700. [Google Scholar] [CrossRef] [PubMed]
- White, E.L.; Suling, W.J.; Ross, L.J.; Seitz, L.E.; Reynolds, R.C. 2-Alkoxycarbonylaminopyridines: Inhibitors of Mycobacterium tuberculosis FtsZ. J. Antimicrob. Chemother. 2002, 50, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, R.; Beuria, T.K.; Mohan, R.; Mahajan, S.K.; Panda, D. Totarol inhibits bacterial cytokinesis by perturbing the assembly dynamics of FtsZ. Biochemistry 2007, 46, 4211–4220. [Google Scholar] [CrossRef]
- Yamamoto, S.; Saito, R.; Nakamura, S.; Sogawa, H.; Karpov, P.; Shulga, S.; Blume, Y.; Kurita, N. Proposal of Potent Inhibitors for a Bacterial Cell Division Protein FtsZ: Molecular Simulations Based on Molecular Docking and ab Initio Molecular Orbital Calculations. Antibiotics 2020, 9, 846. [Google Scholar] [CrossRef] [PubMed]
- Akinpelu, O.I.; Kumalo, H.M.; Mhlongo, S.I.; Mhlongo, N.N. Identifying the analogues of berberine as promising antitubercular drugs targeting Mtb-FtsZ polymerisation through ligand-based virtual screening and molecular dynamics simulations. J. Mol. Recognit. 2022, 35, e2940. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, H.; Zhu, N.; Wang, X.; Han, Y.; Chen, M.; Jiang, J.; Si, S. Identification of TB-E12 as a novel FtsZ inhibitor with anti-tuberculosis activity. Tuberculosis 2018, 110, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Margalit, D.N.; Romberg, L.; Mets, R.B.; Hebert, A.M.; Mitchison, T.J.; Kirschner, M.W.; RayChaudhuri, D. Targeting cell division: Small-molecule inhibitors of FtsZ GTPase perturb cytokinetic ring assembly and induce bacterial lethality. Proc. Natl. Acad. Sci. USA 2004, 101, 11821–11826. [Google Scholar] [CrossRef]
- Alnami, A.; Norton, R.S.; Pena, H.P.; Haider, S.; Kozielski, F. Conformational Flexibility of A Highly Conserved Helix Controls Cryptic Pocket Formation in FtsZ. J. Mol. Biol. 2021, 433, 167061. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.J.; Tang, K.G.; Young, J.; Dandarchuluun, C.; Wong, B.R.; Khurelbaatar, M.; Moroz, Y.S.; Mayfield, J.; Sayle, R.A. ZINC20—A Free Ultralarge-Scale Chemical Database for Ligand Discovery. J. Chem. Inf. Model. 2020, 60, 6065–6073. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Carlos, A.; Jacobo-Delgado, Y.; Santos-Mena, A.O.; García-Hernández, M.H.; De Jesus-Gonzalez, L.A.; Lara-Ramirez, E.E.; Rivas-Santiago, B. Histone deacetylase (HDAC) inhibitors- based drugs are effective to control Mycobacterium tuberculosis infection and promote the sensibility for rifampicin in MDR strain. Mem. Inst. Oswaldo Cruz 2023, 118, e230143. [Google Scholar] [CrossRef] [PubMed]
- Henao Arias, D.C.; Toro, L.J.; Téllez Ramirez, G.A.; Osorio-Méndez, J.F.; Rodríguez-Carlos, A.; Valle, J.; Marín-Luevano, S.P.; Rivas-Santiago, B.; Andreu, D.; Castaño Osorio, J.C. Novel antimicrobial cecropins derived from O. curvicornis and D. satanas dung beetles. Peptides 2021, 145, 170626. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Santiago, B.; de Haro-Acosta, J.; Carlos, A.R.; Garcia-Hernandez, M.H.; Serrano, C.J.; Gonzalez-Curiel, I.; Rivas-Santiago, C. Nicotine promotes Mycobacterium tuberculosis H37Rv growth and overexpression of virulence genes. Microbiol. Immunol. 2023, 67, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J. Sitagliptin: A Review in Type 2 Diabetes. Drugs 2017, 77, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Foletto, V.S.; da Rosa, T.F.; Serafin, M.B.; Bottega, A.; Franco, L.N.; de Paula, B.R.; Hörner, R. Repositioning of antidepressant drugs and synergistic effect with ciprofloxacin against multidrug-resistant bacteria. World J. Microbiol. Biotechnol. 2021, 37, 53. [Google Scholar] [CrossRef]
- Uma Priya, K.; Venkataramaiah, C.; Sreedhar, N.Y.; Raju, C.N. Design, synthesis, characterization and in vitro, in vivo and in silico antimicrobial and antiinflammatory activities of a new series of sulphonamide and carbamate derivatives of a nebivolol intermediate. RSC Adv. 2021, 11, 3897–3916. [Google Scholar] [CrossRef]
- Spencer, C.M.; Goa, K.L. Atovaquone. A review of its pharmacological properties and therapeutic efficacy in opportunistic infections. Drugs 1995, 50, 176–196. [Google Scholar] [CrossRef] [PubMed]
- Capela, R.; Félix, R.; Clariano, M.; Nunes, D.; Perry, M.D.; Lopes, F. Target Identification in Anti-Tuberculosis Drug Discovery. Int. J. Mol. Sci. 2023, 24, 10482. [Google Scholar] [CrossRef] [PubMed]
- Barrows, J.M.; Goley, E.D. FtsZ dynamics in bacterial division: What, how, and why? Curr. Opin. Cell Biol. 2021, 68, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, M.; Mohanty, P.S. Molecular docking in organic, inorganic, and hybrid systems: A tutorial review. Monatshefte Chem. 2023, 154, 683–707. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Chaturvedi, V. Interactive effect of oral anti-hyperglycaemic or anti-hypertensive drugs on the inhibitory and bactericidal activity of first line anti-TB drugs against M. tuberculosis. PLoS ONE 2023, 18, e0292397. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Carlos, A.; Valdez-Miramontes, C.; Marin-Luevano, P.; González-Curiel, I.; Enciso-Moreno, J.A.; Rivas-Santiago, B. Metformin promotes Mycobacterium tuberculosis killing and increases the production of human β-defensins in lung epithelial cells and macrophages. Microbes Infect. 2020, 22, 111–118. [Google Scholar] [CrossRef]
- Erasmus, C.; Aucamp, J.; Smit, F.J.; Seldon, R.; Jordaan, A.; Warner, D.F.; N’Da, D.D. Synthesis and comparison of in vitro dual anti-infective activities of novel naphthoquinone hybrids and atovaquone. Bioorg. Chem. 2021, 114, 105118. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.C.; de Menezes, R.T.; de Oliveira, H.C.; de Oliveira, L.D.; Scorzoni, L. In vitro synergistic effects of fluoxetine and paroxetine in combination with amphotericin B against Cryptococcus neoformans. Pathog. Dis. 2021, 79, ftab001. [Google Scholar] [CrossRef] [PubMed]
- Cabral, V.P.; Rodrigues, D.S.; Barbosa, A.D.; Moreira, L.E.; Sá, L.G.; Silva, C.R.; Neto, J.B.; Silva, J.; Marinho, E.S.; Santos, H.S.; et al. Antibacterial activity of paroxetine against Staphylococcus aureus and possible mechanisms of action. Future Microbiol. 2023, 18, 415–426. [Google Scholar] [CrossRef]
- Caldara, M.; Marmiroli, N. Antimicrobial Properties of Antidepressants and Antipsychotics-Possibilities and Implications. Pharmaceuticals 2021, 14, 915. [Google Scholar] [CrossRef]
- Shen, R.; Zong, K.; Liu, J.; Zhang, L. Risk Factors for Depression in Tuberculosis Patients: A Meta-Analysis. Neuropsychiatr. Dis. Treat. 2022, 18, 847–866. [Google Scholar] [CrossRef] [PubMed]
- Trenton, A.J.; Currier, G.W. Treatment of Comorbid Tuberculosis and Depression. Prim. Care Companion J. Clin. Psychiatry 2001, 3, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Caccia, S. Metabolism of the newer antidepressants. An overview of the pharmacological and pharmacokinetic implications. Clin. Pharmacokinet. 1998, 34, 281–302. [Google Scholar] [CrossRef] [PubMed]
- Prisant, L.M. Nebivolol: Pharmacologic Profile of an Ultraselective, Vasodilatory β1-Blocker. J. Clin. Pharmacol. 2008, 48, 225–239. [Google Scholar] [CrossRef]
- Diego, L.M.; Jazmin, F.M.; Diana, R.H.; German-Isauro, G.F.; Salvador, F.C.; Maria-Elena, H.C. Modulation of TNF-α, interleukin-6, and interleukin-10 by nebivolol-valsartan and nebivolol-lisinopril polytherapy in SHR rats. Pharmacol. Res. Perspect. 2024, 12, e1189. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ZINC ID | Vina Score (kCal/mol) | FDA- Approved Drug | Structure | Reported Activity | Amino Acid Residues | Citation |
|---|---|---|---|---|---|---|
| 1489478 | −9.4 | Sitagliptin | 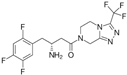 | Inhibition of dipeptidyl peptidase-4 (DPP-4) | GLU-136 ARG-140 | [20] |
| 527386 | −9 | Paroxetine |  | Selective serotonin reuptake inhibitor (SSRI) | THR-106 GLY-105 GLY-19 GLU-102 | [21] |
| 4213946 | −8.6 | Nebivolol |  | Beta-blocker | ASN-22 THR-106 GLY-19 ARG-140 GLU-136 | [22] |
| 12504271 | −8.5 | Atovaquone |  | Antimicrobial, antifungal | ARG-140 | [23] |
| −8.5 | GDP guanosine diphosphate |  | ASP-184 GLU-136 ARG-140 GLY-19 GLY-18 ANS-22 THR-106 GLY-105 GLY-107 |
| FDA-Approved Drug | H37Rv (µg/mL) | MDR (µg/mL) |
|---|---|---|
| Rifampicin | 0.5 | >1 |
| Streptomycin | 0.5 | 0.5 |
| Sitagliptin | >150 | >150 |
| Atovaquone | >10 | >10 |
| Paroxetine | 25 | 25 |
| Nebivolol | 25 | 25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tovar-Nieto, A.M.; Flores-Padilla, L.E.; Rivas-Santiago, B.; Trujillo-Paez, J.V.; Lara-Ramirez, E.E.; Jacobo-Delgado, Y.M.; López-Ramos, J.E.; Rodríguez-Carlos, A. The Repurposing of FDA-Approved Drugs as FtsZ Inhibitors against Mycobacterium tuberculosis: An In Silico and In Vitro Study. Microorganisms 2024, 12, 1505. https://doi.org/10.3390/microorganisms12081505
Tovar-Nieto AM, Flores-Padilla LE, Rivas-Santiago B, Trujillo-Paez JV, Lara-Ramirez EE, Jacobo-Delgado YM, López-Ramos JE, Rodríguez-Carlos A. The Repurposing of FDA-Approved Drugs as FtsZ Inhibitors against Mycobacterium tuberculosis: An In Silico and In Vitro Study. Microorganisms. 2024; 12(8):1505. https://doi.org/10.3390/microorganisms12081505
Chicago/Turabian StyleTovar-Nieto, Andrea Michel, Luis Enrique Flores-Padilla, Bruno Rivas-Santiago, Juan Valentin Trujillo-Paez, Edgar Eduardo Lara-Ramirez, Yolanda M. Jacobo-Delgado, Juan Ernesto López-Ramos, and Adrián Rodríguez-Carlos. 2024. "The Repurposing of FDA-Approved Drugs as FtsZ Inhibitors against Mycobacterium tuberculosis: An In Silico and In Vitro Study" Microorganisms 12, no. 8: 1505. https://doi.org/10.3390/microorganisms12081505