
Impact of Nutrient Enrichment on Community Structure and Co-Occurrence Networks of Coral Symbiotic Microbiota in Duncanopsammia peltata: Zooxanthellae, Bacteria, and Archaea

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Area Description and Sample Collection
2.2. Environmental Parameters
2.3. DNA Extraction and Sequencing
2.4. Sequence Analysis
2.5. Data Analysis
3. Results
3.1. Physico-Chemical Properties in the Surrounding Sediment and Water
3.2. Characteristics of Microbial Communities in Surrounding Sediment and Water
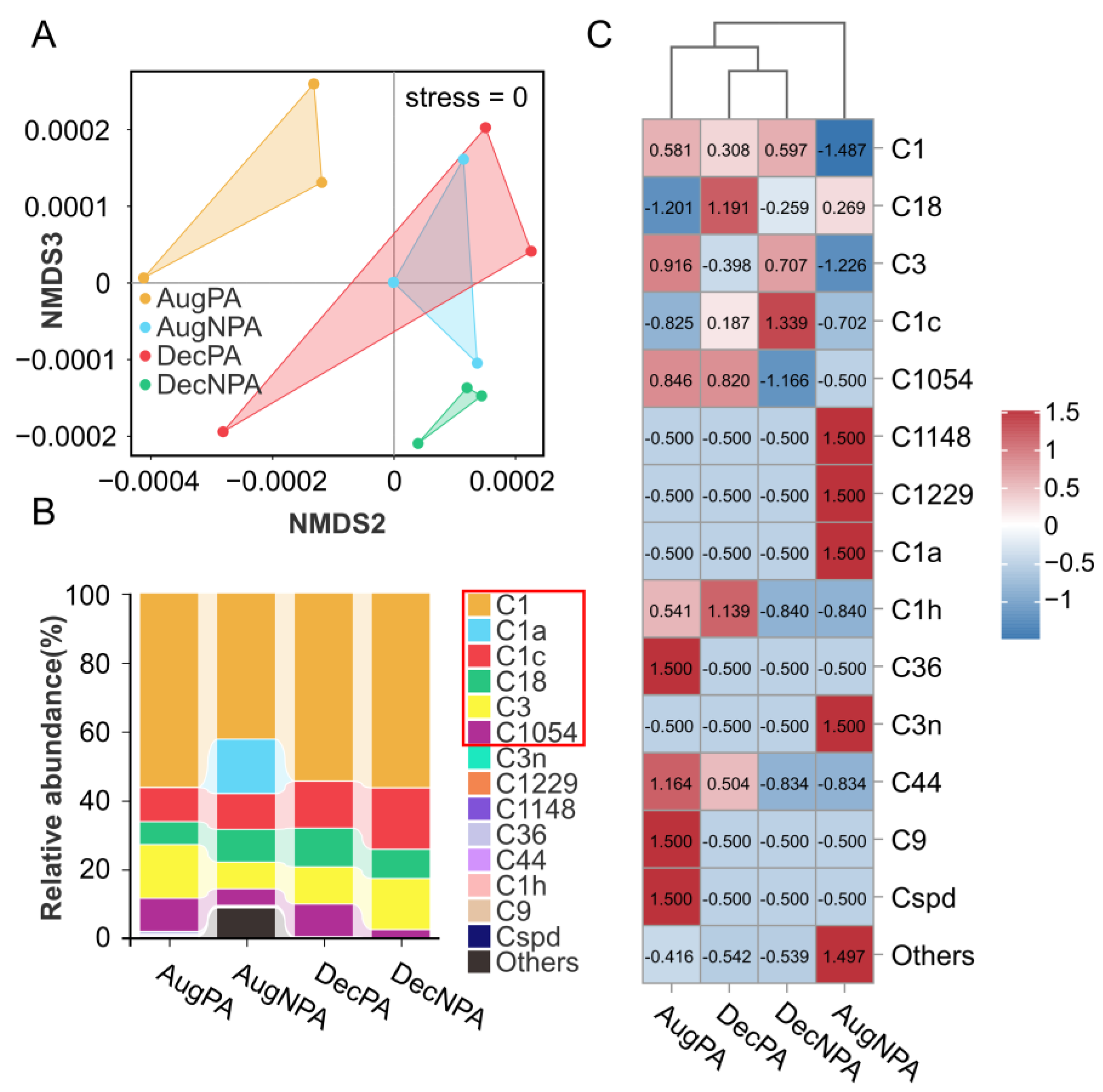
3.3. Diversity and Composition of Zooxanthellae Communities in Coral Symbiotic Microorganisms
3.4. Diversity and Composition of Bacterial Communities in Coral Symbiotic Microorganisms
3.5. Diversity and Composition of Archaeal Communities in Coral Symbiotic Microorganisms
3.6. Correlation Analysis between Microorganisms and Environmental Factors
3.7. Co-Occurrence Network Analysis
4. Discussion
4.1. Coral Symbiotic Microorganisms
4.2. Effects of Environmental Factors on Symbiotic Microorganisms
4.3. Co-Occurrence Network Features
4.4. Composition of Coral Keystone Taxa
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cheng, K.; Tong, M.; Cai, Z.; Jong, M.C.; Zhou, J.; Xiao, B. Prokaryotic and Eukaryotic Microbial Communities Associated with Coral Species Have High Host Specificity in the South China Sea. Sci. Total Environ. 2023, 867, 161185. [Google Scholar] [CrossRef]
- Hughes, T.P.; Barnes, M.L.; Bellwood, D.R.; Cinner, J.E.; Cumming, G.S.; Jackson, J.B.C.; Kleypas, J.; van de Leemput, I.A.; Lough, J.M.; Morrison, T.H.; et al. Coral Reefs in the Anthropocene. Nature 2017, 546, 82–90. [Google Scholar] [CrossRef]
- Ricci, F.; Leggat, W.; Page, C.E.; Ainsworth, T.D. Coral Growth Anomalies, Neoplasms, and Tumors in the Anthropocene. Trends Microbiol. 2022, 30, 1160–1173. [Google Scholar] [CrossRef]
- Zhu, W.; Liu, X.; Zhang, J.; Zhao, H.; Li, Z.; Wang, H.; Chen, R.; Wang, A.; Li, X. Response of Coral Bacterial Composition and Function to Water Quality Variations under Anthropogenic Influence. Sci. Total Environ. 2023, 884, 163837. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.; Bessell-Browne, P.; Jones, R. Synergistic and Antagonistic Impacts of Suspended Sediments and Thermal Stress on Corals. Nat. Commun. 2019, 10, 2346. [Google Scholar] [CrossRef] [PubMed]
- Shantz, A.A.; Burkepile, D.E. Context-Dependent Effects of Nutrient Loading on the Coral–Algal Mutualism. Ecology 2014, 95, 1995–2005. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, G.; Chen, G.; Ni, X.; Guo, L.; Yu, X.; Xiao, C.; Xu, Y.; Shi, X.; Huang, B. Elevated Ammonium Reduces the Negative Effect of Heat Stress on the Stony Coral Pocillopora Damicornis. Mar. Pollut. Bull. 2017, 118, 319–327. [Google Scholar] [CrossRef]
- Burkepile, D.E.; Shantz, A.A.; Adam, T.C.; Munsterman, K.S.; Speare, K.E.; Ladd, M.C.; Rice, M.M.; Ezzat, L.; McIlroy, S.; Wong, J.C.Y.; et al. Nitrogen Identity Drives Differential Impacts of Nutrients on Coral Bleaching and Mortality. Ecosystems 2020, 23, 798–811. [Google Scholar] [CrossRef]
- Wooldridge, S.A.; Done, T.J. Improved Water Quality Can Ameliorate Effects of Climate Change on Corals. Ecol. Appl. 2009, 19, 1492–1499. [Google Scholar] [CrossRef]
- Jin, Y.K.; Kininmonth, S.; Lundgren, P.B.; van Oppen, M.J.H.; Willis, B.L. Predicting the Spatial Distribution of Allele Frequencies for a Gene Associated with Tolerance to Eutrophication and High Temperature in the Reef-Building Coral, Acropora Millepora, on the Great Barrier Reef. Coral Reefs 2020, 39, 147–158. [Google Scholar] [CrossRef]
- Ziegler, M.; Seneca, F.O.; Yum, L.K.; Palumbi, S.R.; Voolstra, C.R. Bacterial Community Dynamics Are Linked to Patterns of Coral Heat Tolerance. Nat. Commun. 2017, 8, 14213. [Google Scholar] [CrossRef] [PubMed]
- Zaneveld, J.R.; Burkepile, D.E.; Shantz, A.A.; Pritchard, C.E.; McMinds, R.; Payet, J.P.; Welsh, R.; Correa, A.M.S.; Lemoine, N.P.; Rosales, S.; et al. Overfishing and Nutrient Pollution Interact with Temperature to Disrupt Coral Reefs down to Microbial Scales. Nat. Commun. 2016, 7, 11833. [Google Scholar] [CrossRef] [PubMed]
- Morris, L.A.; Voolstra, C.R.; Quigley, K.M.; Bourne, D.G.; Bay, L.K. Nutrient Availability and Metabolism Affect the Stability of Coral–Symbiodiniaceae Symbioses. Trends Microbiol. 2019, 27, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zheng, X.; Li, Y.; Sun, D.; Huang, W.; Shi, T. Symbiont Shuffling Dynamics Associated with Photodamage during Temperature Stress in Coral Symbiosis. Ecol. Indic. 2022, 145, 109706. [Google Scholar] [CrossRef]
- Berkelmans, R.; van Oppen, M.J.H. The Role of Zooxanthellae in the Thermal Tolerance of Corals: A ‘Nugget of Hope’ for Coral Reefs in an Era of Climate Change. Proc. R. Soc. B Biol. Sci. 2006, 273, 2305–2312. [Google Scholar] [CrossRef]
- Nalley, E.M.; Tuttle, L.J.; Conklin, E.E.; Barkman, A.L.; Wulstein, D.M.; Schmidbauer, M.C.; Donahue, M.J. A Systematic Review and Meta-Analysis of the Direct Effects of Nutrients on Corals. Sci. Total Environ. 2023, 856, 159093. [Google Scholar] [CrossRef]
- Bourne, D.G.; Morrow, K.M.; Webster, N.S. Insights into the Coral Microbiome: Underpinning the Health and Resilience of Reef Ecosystems. Annu. Rev. Microbiol. 2016, 70, 317–340. [Google Scholar] [CrossRef]
- Dunphy, C.M.; Gouhier, T.C.; Chu, N.D.; Vollmer, S.V. Structure and Stability of the Coral Microbiome in Space and Time. Sci. Rep. 2019, 9, 6785. [Google Scholar] [CrossRef]
- Osman, E.O.; Suggett, D.J.; Voolstra, C.R.; Pettay, D.T.; Clark, D.R.; Pogoreutz, C.; Sampayo, E.M.; Warner, M.E.; Smith, D.J. Coral Microbiome Composition along the Northern Red Sea Suggests High Plasticity of Bacterial and Specificity of Endosymbiotic Dinoflagellate Communities. Microbiome 2020, 8, 8. [Google Scholar] [CrossRef]
- Ziegler, M.; Grupstra, C.G.B.; Barreto, M.M.; Eaton, M.; BaOmar, J.; Zubier, K.; Al-Sofyani, A.; Turki, A.J.; Ormond, R.; Voolstra, C.R. Coral Bacterial Community Structure Responds to Environmental Change in a Host-Specific Manner. Nat. Commun. 2019, 10, 3092. [Google Scholar] [CrossRef]
- Siboni, N.; Ben-Dov, E.; Sivan, A.; Kushmaro, A. Global Distribution and Diversity of Coral-Associated Archaea and Their Possible Role in the Coral Holobiont Nitrogen Cycle. Environ. Microbiol. 2008, 10, 2979–2990. [Google Scholar] [CrossRef] [PubMed]
- Messyasz, A.; Maher, R.L.; Meiling, S.S.; Thurber, R.V. Nutrient Enrichment Predominantly Affects Low Diversity Microbiomes in a Marine Trophic Symbiosis between Algal Farming Fish and Corals. Microorganisms 2021, 9, 1873. [Google Scholar] [CrossRef]
- Cárdenas, A.; Rodriguez-R, L.M.; Pizarro, V.; Cadavid, L.F.; Arévalo-Ferro, C. Shifts in Bacterial Communities of Two Caribbean Reef-Building Coral Species Affected by White Plague Disease. ISME J. 2012, 6, 502–512. [Google Scholar] [CrossRef]
- Bourne, D.G.; Garren, M.; Work, T.M.; Rosenberg, E.; Smith, G.W.; Harvell, C.D. Microbial Disease and the Coral Holobiont. Trends Microbiol. 2009, 17, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Sweet, M.J.; Bulling, M.T. On the Importance of the Microbiome and Pathobiome in Coral Health and Disease. Front. Mar. Sci. 2017, 4, 9. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, Q.; Dong, X.; Wang, A.; Wang, J.; Chen, B. A New Perspective of Nutrient Management of Subtropical Coastal Stress-Tolerant Scleractinian Coral Communities. Cont. Shelf Res. 2021, 220, 104405. [Google Scholar] [CrossRef]
- Du, Y.; Meng, F.; Fu, W.; Wang, Z. Distribution, Speciation and Bioaccumulation of Hg and As in Mariculture Sediments from Dongshan Bay, China. Soil Sediment Contam. Int. J. 2016, 25, 489–504. [Google Scholar] [CrossRef]
- Wu, Q.; Huang, W.; Chen, B.; Yang, E.; Meng, L.; Chen, Y.; Li, J.; Huang, X.; Liang, J.; Yap, T.-K.; et al. Genetic Structure of Turbinaria Peltata in the Northern South China Sea Suggest Insufficient Genetic Adaptability of Relatively High-Latitude Scleractinian Corals to Environment Stress. Sci. Total Environ. 2021, 775, 145775. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Yu, K.; Liao, Z.; Yu, X.; Qin, Z.; Liang, J.; Wang, G.; Wu, Q.; Jiang, L. Microbiome Community and Complexity Indicate Environmental Gradient Acclimatisation and Potential Microbial Interaction of Endemic Coral Holobionts in the South China Sea. Sci. Total Environ. 2021, 765, 142690. [Google Scholar] [CrossRef] [PubMed]
- LY/T 1237-1999; Determination of Organic Matter in Forest Soil and Calculation Carbon-Nitrogen Ratio. Forest Soil Research Laboratory, Forestry Research Institute, Chinese Academy of Forestry: Beijing, China, 1999.
- HJ 717-2014; Soil Quality—Determination of Total Nitrogen—Modified Kjeldahl Method. Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China, 2014.
- NY/T 88-1988; Method for Determination of soil Total Phosphorus. Analysis and Testing Center of Chinese Academy of Agricultural Sciences: Beijing, China, 1998.
- Shi, T.; Niu, G.; Kvitt, H.; Zheng, X.; Qin, Q.; Sun, D.; Ji, Z.; Tchernov, D. Untangling ITS2 Genotypes of Algal Symbionts in Zooxanthellate Corals. Mol. Ecol. Resour. 2021, 21, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using Network Analysis to Explore Co-Occurrence Patterns in Soil Microbial Communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef]
- Chen, B.; Wang, K.; Peng, C.; Dong, X.; Wang, S.; Lin, H. Changes of Eutrophication Degree in Dongshan Bay in China Affected by the COVID-19 Outbreak. Reg. Stud. Mar. Sci. 2023, 66, 103119. [Google Scholar] [CrossRef]
- Banerjee, S.; Schlaeppi, K.; van der Heijden, M.G.A. Keystone Taxa as Drivers of Microbiome Structure and Functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef] [PubMed]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 2018, 28, 2570–2580.e6. [Google Scholar] [CrossRef]
- Ziegler, M.; Arif, C.; Burt, J.A.; Dobretsov, S.; Roder, C.; LaJeunesse, T.C.; Voolstra, C.R. Biogeography and Molecular Diversity of Coral Symbionts in the Genus Symbiodinium around the Arabian Peninsula. J. Biogeogr. 2017, 44, 674–686. [Google Scholar] [CrossRef]
- Stat, M.; Yost, D.M.; Gates, R.D. Geographic Structure and Host Specificity Shape the Community Composition of Symbiotic Dinoflagellates in Corals from the Northwestern Hawaiian Islands. Coral Reefs 2015, 34, 1075–1086. [Google Scholar] [CrossRef]
- Robbins, S.J.; Singleton, C.M.; Chan, C.X.; Messer, L.F.; Geers, A.U.; Ying, H.; Baker, A.; Bell, S.C.; Morrow, K.M.; Ragan, M.A.; et al. A Genomic View of the Reef-Building Coral Porites Lutea and Its Microbial Symbionts. Nat. Microbiol. 2019, 4, 2090–2100. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Yu, K.; Qin, Z.; Liang, J.; Wang, G.; Huang, X.; Wu, Q.; Jiang, L. Dispersal, Genetic Variation, and Symbiont Interaction Network of Heat-Tolerant Endosymbiont Durusdinium trenchii: Insights into the Adaptive Potential of Coral to Climate Change. Sci. Total Environ. 2020, 723, 138026. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.Y.; Ang, P. Low Symbiont Diversity as a Potential Adaptive Strategy in a Marginal Non-Reefal Environment: A Case Study of Corals in Hong Kong. Coral Reefs 2016, 35, 941–957. [Google Scholar] [CrossRef]
- Reimer, J.D.; Takishita, K.; Maruyama, T. Molecular Identification of Symbiotic Dinoflagellates (Symbiodinium spp.) from Palythoa Spp. (Anthozoa: Hexacorallia) in Japan. Coral Reefs 2006, 25, 521–527. [Google Scholar] [CrossRef]
- Hughes, T.P.; Baird, A.H.; Bellwood, D.R.; Card, M.; Connolly, S.R.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.B.C.; Kleypas, J.; et al. Climate Change, Human Impacts, and the Resilience of Coral Reefs. Science 2003, 301, 929–933. [Google Scholar] [CrossRef]
- Qin, Z.; Yu, K.; Chen, B.; Wang, Y.; Liang, J.; Luo, W.; Xu, L.; Huang, X. Diversity of Symbiodiniaceae in 15 Coral Species From the Southern South China Sea: Potential Relationship With Coral Thermal Adaptability. Front. Microbiol. 2019, 10, 2343. [Google Scholar] [CrossRef] [PubMed]
- Jandang, S.; Viyakarn, V.; Yoshioka, Y.; Shinzato, C.; Chavanich, S. The Seasonal Investigation of Symbiodiniaceae in Broadcast Spawning, Acropora Humilis and Brooding, Pocillopora Cf. Damicornis Corals. PeerJ 2022, 10, e13114. [Google Scholar] [CrossRef] [PubMed]
- Sampayo, E.M.; Ridgway, T.; Bongaerts, P.; Hoegh-Guldberg, O. Bleaching Susceptibility and Mortality of Corals Are Determined by Fine-Scale Differences in Symbiont Type. Proc. Natl. Acad. Sci. USA 2008, 105, 10444–10449. [Google Scholar] [CrossRef] [PubMed]
- Ong, J.H.; Wainwright, B.J.; Jain, S.S.; Afiq-Rosli, L.; Lee, J.N.; Huang, D. Species and Spatio-Environmental Effects on Coral Endosymbiont Communities in Southeast Asia. Coral Reefs 2022, 41, 1131–1145. [Google Scholar] [CrossRef]
- Sweet, M.; Villela, H.; Keller-Costa, T.; Costa, R.; Romano, S.; Bourne, D.G.; Cárdenas, A.; Huggett, M.J.; Kerwin, A.H.; Kuek, F.; et al. Insights into the Cultured Bacterial Fraction of Corals. mSystems 2021, 6, e01249-20. [Google Scholar] [CrossRef]
- Zubia, M.; Vieira, C.; Palinska, K.A.; Roué, M.; Gaertner, J.-C.; Zloch, I.; Grellier, M.; Golubic, S. Benthic Cyanobacteria on Coral Reefs of Moorea Island (French Polynesia): Diversity Response to Habitat Quality. Hydrobiologia 2019, 843, 61–78. [Google Scholar] [CrossRef]
- Boeuf, D.; Eppley, J.M.; Mende, D.R.; Malmstrom, R.R.; Woyke, T.; DeLong, E.F. Metapangenomics Reveals Depth-Dependent Shifts in Metabolic Potential for the Ubiquitous Marine Bacterial SAR324 Lineage. Microbiome 2021, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wu, Z.; Tang, S. Extracellular Polymeric Substances (EPS) Properties and Their Effects on Membrane Fouling in a Submerged Membrane Bioreactor. Water Res. 2009, 43, 2504–2512. [Google Scholar] [CrossRef]
- McDevitt-Irwin, J.M.; Baum, J.K.; Garren, M.; Vega Thurber, R.L. Responses of Coral-Associated Bacterial Communities to Local and Global Stressors. Front. Mar. Sci. 2017, 4, 262. [Google Scholar] [CrossRef]
- Mera, H.; Bourne, D.G. Disentangling Causation: Complex Roles of Coral-associated Microorganisms in Disease. Environ. Microbiol. 2018, 20, 431–449. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.R.; Ochsenkühn, M.A.; Kazlak, A.M.; Moustafa, A.; Amin, S.A. The Coral Microbiome: Towards an Understanding of the Molecular Mechanisms of Coral–Microbiota Interactions. FEMS Microbiol. Rev. 2023, 47, fuad005. [Google Scholar] [CrossRef]
- Santoro, E.P.; Borges, R.M.; Espinoza, J.L.; Freire, M.; Messias, C.S.M.A.; Villela, H.D.M.; Pereira, L.M.; Vilela, C.L.S.; Rosado, J.G.; Cardoso, P.M.; et al. Coral Microbiome Manipulation Elicits Metabolic and Genetic Restructuring to Mitigate Heat Stress and Evade Mortality. Sci. Adv. 2021, 7, eabg3088. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Cheng, Y.; Guo, C.; Xie, F.; Jung, D.; Zhang, W.; He, S. Nisaea sediminum Sp. Nov., a Heavy Metal Resistant Bacterium Isolated from Marine Sediment in the East China Sea. Antonie Van Leeuwenhoek 2021, 114, 2113–2121. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, V.G.; Mani, K.; Senthil Kumar, P.; Rangasamy, G.; Sridharan, R.; Rethnaraj, C.; Amirtha Ganesh, S.S.; Kalidas, S.; Palanisamy, V.; Chellama, N.J.; et al. Prevalence of Differential Microbiome in Healthy, Diseased and Nipped Colonies of Corals, Porites Lutea in the Gulf of Kachchh, North-West Coast of India. Environ. Res. 2023, 216, 114622. [Google Scholar] [CrossRef]
- Thompson, F.L.; Barash, Y.; Sawabe, T.; Sharon, G.; Swings, J.; Rosenberg, E. Thalassomonas loyana sp. nov., a Causative Agent of the White Plague-like Disease of Corals on the Eilat Coral Reef. Int. J. Syst. Evol. Microbiol. 2006, 56, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, B.; Smith, A.; Aeby, G.S.; Callahan, S.M. Vibrio Owensii Induces the Tissue Loss Disease Montipora White Syndrome in the Hawaiian Reef Coral Montipora Capitata. PLoS ONE 2012, 7, e46717. [Google Scholar] [CrossRef] [PubMed]
- Littman, R.; Willis, B.L.; Bourne, D.G. Metagenomic Analysis of the Coral Holobiont during a Natural Bleaching Event on the Great Barrier Reef. Environ. Microbiol. Rep. 2011, 3, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Cardini, U.; van Hoytema, N.; Bednarz, V.N.; Rix, L.; Foster, R.A.; Al-Rshaidat, M.M.D.; Wild, C. Microbial Dinitrogen Fixation in Coral Holobionts Exposed to Thermal Stress and Bleaching. Environ. Microbiol. 2016, 18, 2620–2633. [Google Scholar] [CrossRef]
- Frade, P.R.; Roll, K.; Bergauer, K.; Herndl, G.J. Archaeal and Bacterial Communities Associated with the Surface Mucus of Caribbean Corals Differ in Their Degree of Host Specificity and Community Turnover Over Reefs. PLoS ONE 2016, 11, e0144702. [Google Scholar] [CrossRef]
- Megyes, M.; Móga, J.; Strat, D.; Borsodi, A.K. Bacterial and Archaeal Taxonomic Diversity of Mud Volcanoes (Beciu, Romania) via Metagenomic Approach. Geomicrobiol. J. 2021, 38, 532–539. [Google Scholar] [CrossRef]
- Jameson, E.; Stephenson, J.; Jones, H.; Millard, A.; Kaster, A.-K.; Purdy, K.J.; Airs, R.; Murrell, J.C.; Chen, Y. Deltaproteobacteria (Pelobacter) and Methanococcoides Are Responsible for Choline-Dependent Methanogenesis in a Coastal Saltmarsh Sediment. ISME J. 2019, 13, 277–289. [Google Scholar] [CrossRef] [PubMed]
- D Ainsworth, T.; Krause, L.; Bridge, T.; Torda, G.; Raina, J.-B.; Zakrzewski, M.; Gates, R.D.; Padilla-Gamiño, J.L.; Spalding, H.L.; Smith, C.; et al. The Coral Core Microbiome Identifies Rare Bacterial Taxa as Ubiquitous Endosymbionts. ISME J. 2015, 9, 2261–2274. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Agreda, A.; Leggat, W.; Bongaerts, P.; Herrera, C.; Ainsworth, T.D. Rethinking the Coral Microbiome: Simplicity Exists within a Diverse Microbial Biosphere. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Agreda, A.; Leggat, W.; Bongaerts, P.; Ainsworth, T.D. The Microbial Signature Provides Insight into the Mechanistic Basis of Coral Success across Reef Habitats. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- van Oppen, M.J.H.; Blackall, L.L. Coral Microbiome Dynamics, Functions and Design in a Changing World. Nat. Rev. Microbiol. 2019, 17, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, J.A. Microbial Community Structure and Its Functional Implications. Nature 2009, 459, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Matchado, M.S.; Lauber, M.; Reitmeier, S.; Kacprowski, T.; Baumbach, J.; Haller, D.; List, M. Network Analysis Methods for Studying Microbial Communities: A Mini Review. Comput. Struct. Biotechnol. J. 2021, 19, 2687–2698. [Google Scholar] [CrossRef]
- Faust, K.; Raes, J. Microbial Interactions: From Networks to Models. Nat. Rev. Microbiol. 2012, 10, 538–550. [Google Scholar] [CrossRef]
- Deng, Y.; Jiang, Y.-H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular Ecological Network Analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The Modularity of Pollination Networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef]
- Newman, M.E.J. Modularity and Community Structure in Networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef] [PubMed]
- Kitano, H. Biological Robustness. Nat. Rev. Genet. 2004, 5, 826–837. [Google Scholar] [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; et al. Soil Bacterial Networks Are Less Stable under Drought than Fungal Networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Xiao, G.; Zhou, J.; Wang, Y.; Wang, Z.; Kurths, J.; Schellnhuber, H.J. System Crash as Dynamics of Complex Networks. Proc. Natl. Acad. Sci. USA 2016, 113, 11726–11731. [Google Scholar] [CrossRef] [PubMed]
- Karimi, B.; Maron, P.A.; Chemidlin-Prevost Boure, N.; Bernard, N.; Gilbert, D.; Ranjard, L. Microbial Diversity and Ecological Networks as Indicators of Environmental Quality. Environ. Chem. Lett. 2017, 15, 265–281. [Google Scholar] [CrossRef]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The Ecology of the Microbiome: Networks, Competition, and Stability. Science 2015, 350, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Yang, H.; Shi, Q.; Wang, G. Changes in Coral Bacterial Communities during a Natural Bleaching Event Linked to El Niño in the South China Sea. Reg. Stud. Mar. Sci. 2022, 53, 102383. [Google Scholar] [CrossRef]
- Bascompte, J.; Stouffer, D.B. The Assembly and Disassembly of Ecological Networks. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1781–1787. [Google Scholar] [CrossRef]
- Stat, M.; Pochon, X.; Cowie, R.O.M.; Gates, R.D. Specificity in Communities of Symbiodinium in Corals from Johnston Atoll. Mar. Ecol. Prog. Ser. 2009, 386, 83–96. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic Identification and in Situ Detection of Individual Microbial Cells without Cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef] [PubMed]
- Rohwer, F.; Seguritan, V.; Azam, F.; Knowlton, N. Diversity and Distribution of Coral-Associated Bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar] [CrossRef]
- Guo, M.; Wu, F.; Hao, G.; Qi, Q.; Li, R.; Li, N.; Wei, L.; Chai, T. Bacillus Subtilis Improves Immunity and Disease Resistance in Rabbits. Front. Immunol. 2017, 8, 354. [Google Scholar] [CrossRef] [PubMed]
- Pires, A.C.C.; Cleary, D.F.R.; Almeida, A.; Cunha, Â.; Dealtry, S.; Mendonça-Hagler, L.C.S.; Smalla, K.; Gomes, N.C.M. Denaturing Gradient Gel Electrophoresis and Barcoded Pyrosequencing Reveal Unprecedented Archaeal Diversity in Mangrove Sediment and Rhizosphere Samples. Appl. Environ. Microbiol. 2012, 78, 5520–5528. [Google Scholar] [CrossRef] [PubMed]
- Bobadilla Fazzini, R.A.; Levican, G.; Parada, P. Acidithiobacillus Thiooxidans Secretome Containing a Newly Described Lipoprotein Licanantase Enhances Chalcopyrite Bioleaching Rate. Appl. Microbiol. Biotechnol. 2011, 89, 771–780. [Google Scholar] [CrossRef]
- Yu, Y.; Lee, C.; Kim, J.; Hwang, S. Group-Specific Primer and Probe Sets to Detect Methanogenic Communities Using Quantitative Real-Time Polymerase Chain Reaction. Biotechnol. Bioeng. 2005, 89, 670–679. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| August Protected Area | August Non-Protected Area | December Protected Area | December Non-Protected Area | |
|---|---|---|---|---|
| TOC (g/kg) 1 | 2.83 | 5.11 | 4.37 | 14.52 |
| TP (g/kg) 1 | 0.39 | 0.48 | 0.36 | 1.09 |
| TN (g/kg) 1 | 0.66 | 0.86 | 0.47 | 1.38 |
| Turbidity (NTU) 2 | 3.55 ± 1.35 | 6.10 ± 2.25 | 4.84 ± 2.26 | 9.63 ± 2.94 |
| DO (mg/L) 2 | 7.17 ± 0.72 | 4.36 ± 0.60 | 7.28 ± 0.37 | 6.47 ± 0.35 |
| Chl a (μg/L) 2 | 5.26 ± 2.26 | 2.04 ± 1.10 | 4.12 ± 2.75 | 1.73 ± 0.40 |
| Salinity (‰) 2 | 33.58 ± 0.39 | 33.27 ± 0.86 | 32.36 ± 0.15 | 32.46 ± 0.08 |
| pH 2 | 8.10 ± 0.04 | 7.89 ± 0.05 | 8.20 ± 0.03 | 8.08 ± 0.03 |
| Temperature (°C) 2 | 25.92 ± 0.69 | 27.08 ± 0.72 | 21.97 ± 0.52 | 21.79 ± 0.96 |
| DIN (μM) 3 | 6.38 | 11.21 | 23.51 | 23.02 |
| PO43−-P (μM) 3 | 0.54 | 0.74 | 1.35 | 1.39 |
| N/P 3 | 11.81 | 15.15 | 17.41 | 16.56 |
| NO3−-N (μM) 3 | 3.55 | 5.66 | 22.64 | 21.46 |
| NO2−-N (μM) 3 | 0.55 | 0.51 | 0.26 | 0.42 |
| NH4+-N (μM) 3 | 2.28 | 5.04 | 0.61 | 1.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, C.; Wang, Q.; Xu, J.; Zhang, H.; Huang, Y.; Cai, L.; Zheng, X.; Yang, M. Impact of Nutrient Enrichment on Community Structure and Co-Occurrence Networks of Coral Symbiotic Microbiota in Duncanopsammia peltata: Zooxanthellae, Bacteria, and Archaea. Microorganisms 2024, 12, 1540. https://doi.org/10.3390/microorganisms12081540
Bai C, Wang Q, Xu J, Zhang H, Huang Y, Cai L, Zheng X, Yang M. Impact of Nutrient Enrichment on Community Structure and Co-Occurrence Networks of Coral Symbiotic Microbiota in Duncanopsammia peltata: Zooxanthellae, Bacteria, and Archaea. Microorganisms. 2024; 12(8):1540. https://doi.org/10.3390/microorganisms12081540
Chicago/Turabian StyleBai, Chuanzhu, Qifang Wang, Jinyan Xu, Han Zhang, Yuxin Huang, Ling Cai, Xinqing Zheng, and Ming Yang. 2024. "Impact of Nutrient Enrichment on Community Structure and Co-Occurrence Networks of Coral Symbiotic Microbiota in Duncanopsammia peltata: Zooxanthellae, Bacteria, and Archaea" Microorganisms 12, no. 8: 1540. https://doi.org/10.3390/microorganisms12081540
APA StyleBai, C., Wang, Q., Xu, J., Zhang, H., Huang, Y., Cai, L., Zheng, X., & Yang, M. (2024). Impact of Nutrient Enrichment on Community Structure and Co-Occurrence Networks of Coral Symbiotic Microbiota in Duncanopsammia peltata: Zooxanthellae, Bacteria, and Archaea. Microorganisms, 12(8), 1540. https://doi.org/10.3390/microorganisms12081540