
The Improving Effects of Probiotic-Added Pollen Substitute Diets on the Gut Microbiota and Individual Health of Honey Bee (Apis mellifera L.)
 ,
,  ,
,
Abstract
:1. Introduction
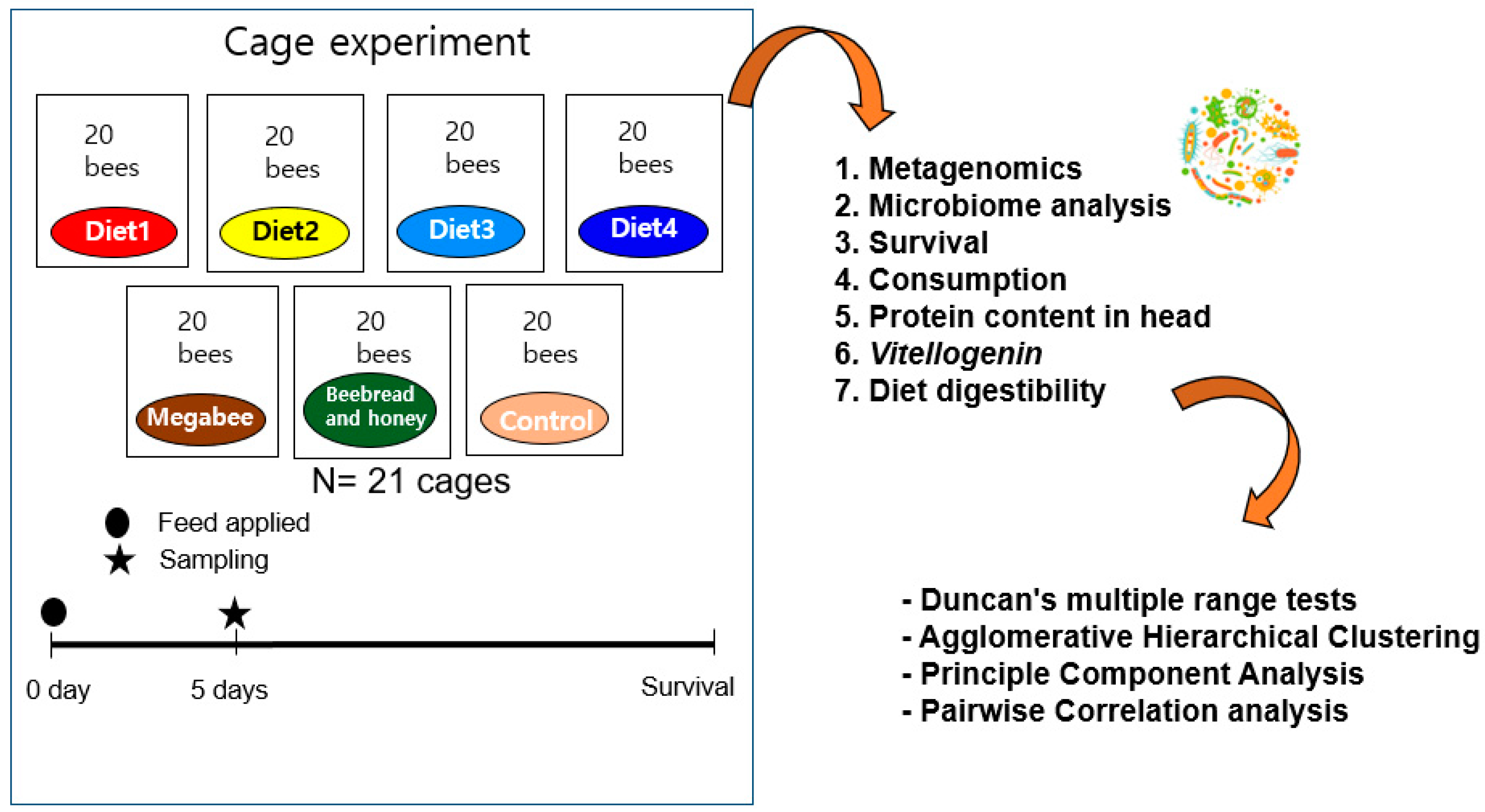
2. Materials and Methods
2.1. Diet Preparation
2.2. Cage Experiment and Sampling
2.3. DNA Extraction
2.4. RNA Extraction and Quantitative PCR (qPCR)
2.5. Sample Preparation, 16s Sequencing, and Taxonomic Analysis
2.6. Analysis in QIIME2
2.7. Bacterial Profiles
2.8. Statistical Analysis
3. Results
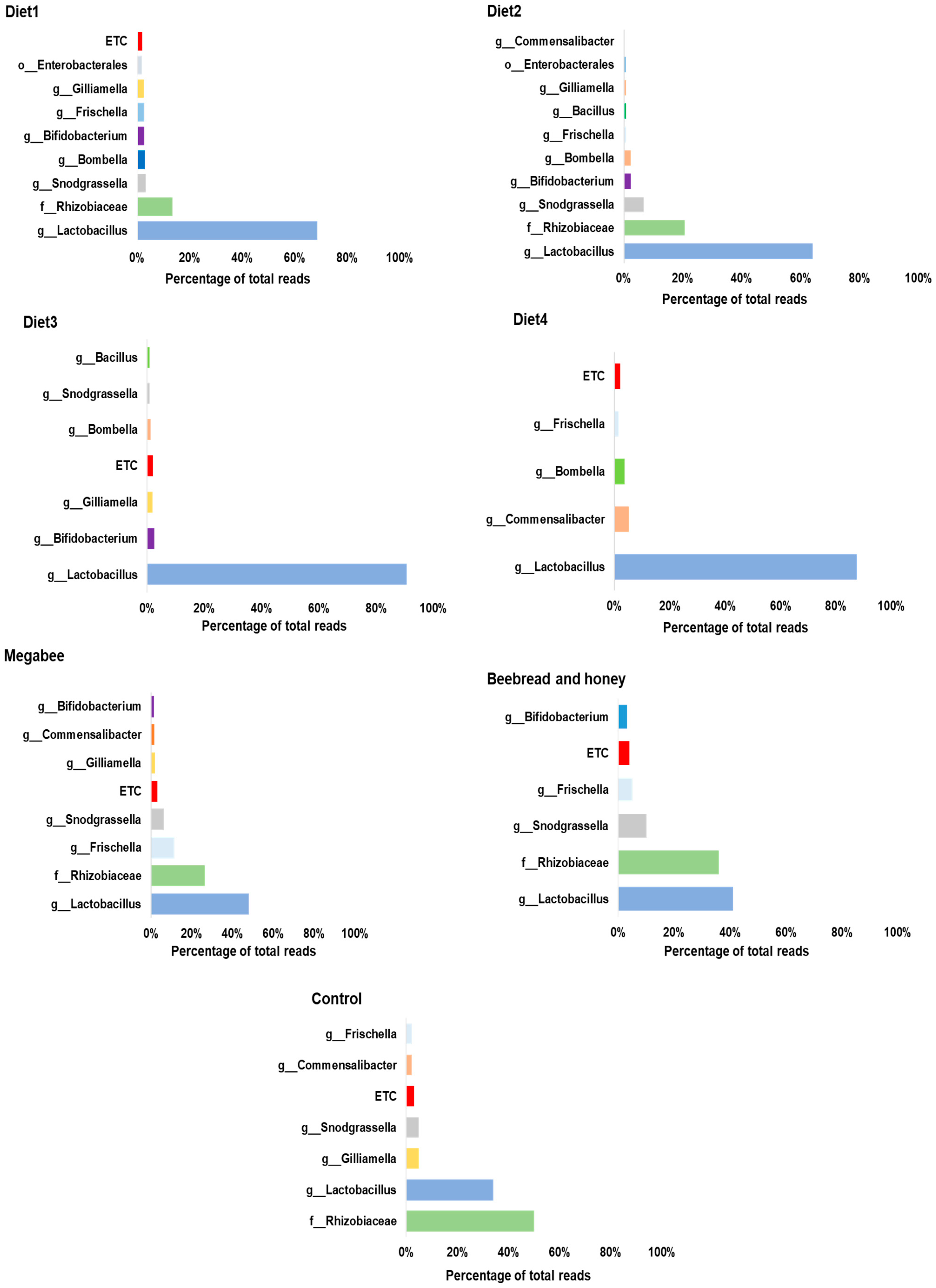
3.1. Reads Profiling for Microbial Community
3.2. Gut Microbiota Diversity and Richness between Different Diet Groups
3.2.1. Alpha Diversity
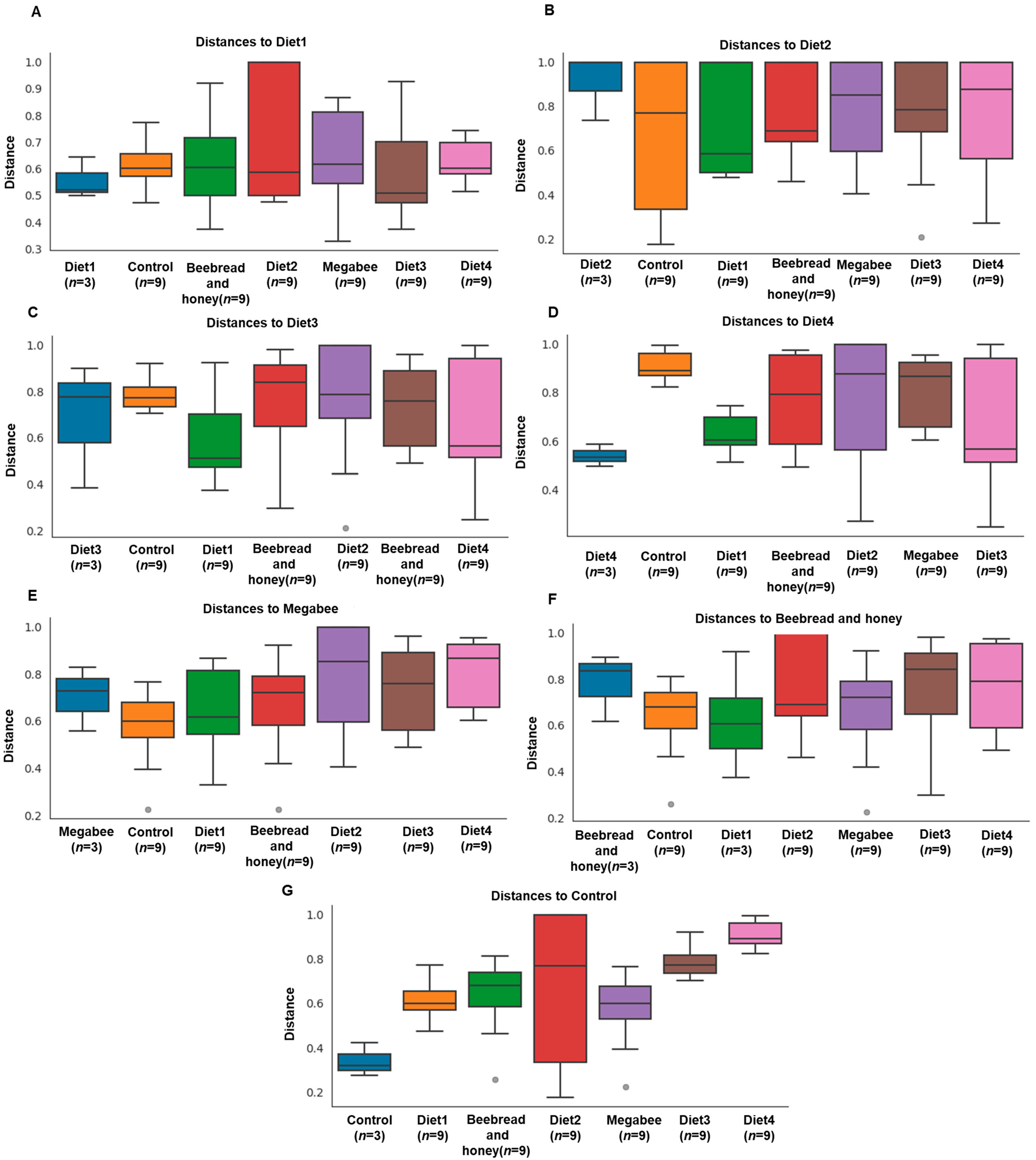
3.2.2. Beta Diversity
3.3. Gut Microbiota Taxonomy Diversity Based on Different Diets in Apis mellifera L.
3.4. Cage Experiments Depend on Different Diets
3.5. Diet Group Distribution and Correlation
3.6. Diet Group Correlation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.-P.; vanEngelsdorp, D. Drivers of colony losses. Curr. Opin. Insect Sci. 2018, 26, 142–148. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Amdam, G.V.; Omholt, S.W. The hive bee to forager transition in honeybee colonies: The double repressor hypothesis. J. Theor. Biol. 2003, 223, 451–464. [Google Scholar] [CrossRef]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Crailsheim, K.; Schneider, L.H.W.; Hrassnigg, N.; Bühlmann, G.; Brosch, U.; Gmeinbauer, R.; Schöffmann, B. Pollen consumption and utilization in worker honeybees (Apis mellifera carnica): Dependence on individual age and function. J. Insect Physiol. 1992, 38, 409–419. [Google Scholar] [CrossRef]
- Donkersley, P.; Rhodes, G.; Pickup, R.W.; Jones, K.C.; Wilson, K. Honeybee nutrition is linked to landscape composition. Ecol. Evol. 2014, 4, 4195–4206. [Google Scholar] [CrossRef]
- Morais, M.M.; Turcatto, A.P.; Pereira, R.A.; Francoy, T.M.; Guidugli-Lazzarini, K.R.; Goncalves, L.S.; De Almeida, J.; Ellis, J.; De Jong, D. Protein levels and colony development of africanized and european honey bees fed natural and artificial diets. Genet. Mol. Res. 2013, 12, 6915–6922. [Google Scholar] [CrossRef]
- Noordyke, E.R.; Ellis, J.D. Reviewing the Efficacy of Pollen Substitutes as a Management Tool for Improving the Health and Productivity of Western Honey Bee (Apis mellifera) Colonies. Front. Sustain. Food Syst. 2021, 5, 772897. [Google Scholar] [CrossRef]
- Kim, H.; Frunze, O.; Maigoro, A.Y.; Lee, M.L.; Lee, J.H.; Kwon, H.W. Comparative Study of the Effect of Pollen Substitute Diets on Honey Bees during Early Spring. Insects 2024, 15, 101. [Google Scholar] [CrossRef]
- Fèvre, D.P.; Dearden, P.K. Influence of nutrition on honeybee queen egg-laying. Apidologie 2024, 55, 53. [Google Scholar] [CrossRef]
- Evans, J.D.; Armstrong, T.N. Antagonistic interactions between honey bee bacterial symbionts and implications for disease. BMC Ecol. 2006, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Sugimura, Y.; Takaya, N.; Takamatsu, D.; Kobayashi, M.; Taylor, D.; Yoshiyama, M. Characterization of bifidobacteria in the digestive tract of the Japanese honeybee, Apis cerana japonica. J. Invertebr. Pathol. 2013, 112, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Raymann, K.; Moran, N.A. The role of the gut microbiome in health and disease of adult honey bee workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.C.; Fruciano, C.; Hildebrand, F.; Al Toufalilia, H.; Balfour, N.J.; Bork, P.; Engel, P.; Ratnieks, F.L.; Hughes, W.O. Gut microbiota composition is associated with environmental landscape in honey bees. Ecol. Evol. 2018, 8, 441–451. [Google Scholar] [CrossRef]
- Huang, Y.H.; Chen, Y.H.; Chen, J.H.; Hsu, P.S.; Wu, T.H.; Lin, C.F.; Peng, C.C.; Wu, M.C. A potential probiotic Leuconostoc mesenteroides TBE-8 for honeybee. Sci. Rep. 2021, 11, 1846. [Google Scholar]
- Maes, P.W.; Rodrigues, P.A.; Oliver, R.; Mott, B.M.; Anderson, K.E. Diet--related gut bacterial dysbiosis correlates with impaired development, increased mortality and Nosema disease in the honeybee (Apis mellifera). Mol. Ecol. 2016, 25, 5439–5450. [Google Scholar] [CrossRef]
- Maigoro, A.Y.; Lee, J.-H.; Kim, H.; Frunze, O.; Kwon, H.-W. Gut Microbiota of Apis mellifera at Selected Ontogenetic Stages and Their Immunogenic Potential during Summer. Pathogens 2024, 13, 122. [Google Scholar] [CrossRef]
- Dillon, R.J.; Webster, G.; Weightman, A.J.; Keith Charnley, A. Diversity of gut microbiota increases with aging and starvation in the desert locust. Antonie Leeuwenhoek 2010, 97, 69–77. [Google Scholar] [CrossRef]
- Ma, W.; Zheng, X.; Li, L.; Shen, J.; Li, W.; Gao, Y. Changes in the gut microbiota of honey bees associated with jujube flower disease. Ecotoxicol. Environ. Saf. 2020, 198, 110616. [Google Scholar] [CrossRef]
- Endo, A.; Salminen, S. Honeybees and beehives are rich sources for fructophilic lactic acid bacteria. Syst. Appl. Microbiol. 2013, 36, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Hubert, J.; Bicianova, M.; Ledvinka, O.; Kamler, M.; Lester, P.J.; Nesvorna, M.; Kopecky, J.; Erban, T. Changes in the bacteriome of honey bees associated with the parasite Varroa destructor, and pathogens Nosema and Lotmaria passim. Microb. Ecol. 2017, 73, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Engel, P.; Moran, N.A. The gut microbiota of insects–diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [PubMed]
- Smriti; Rana, A.; Singh, G.; Gupta, G. Prospects of Probiotics in Beekeeping: A Review for Sustainable Approach to Boost Honeybee Health. Arch. Microbiol. 2024, 206, 205. [Google Scholar] [CrossRef] [PubMed]
- Besharati, M.; Bavand, R.; Paya, H.; Lackner, M. Comparative effect of probiotic and antibiotic on honeybee’s colony functional traits. EuroBiotech J. 2024, 8, 1–11. [Google Scholar] [CrossRef]
- Ricigliano, V.A.; Williams, S.T.; Oliver, R. Effects of different artificial diets on commercial honeybee colony performance, health biomarkers, and gut microbiota. BMC Vet. Res. 2022, 18, 52. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.; Wang, Y.; Gu, H.; Zhu, Q.; Liu, W.; Dai, Y.; Li, F.; Li, B. Effects of artificial diet breeding on intestinal microbial populations at the young stage of silkworm (Bombyx mori). Arch. Insect Biochem. Physiol. 2023, 113, e22019. [Google Scholar] [CrossRef]
- Trevelline, B.K.; Kohl, K.D. The gut microbiome influences host diet selection behavior. Proc. Natl Acad. Sci. USA 2022, 119, e2117537119. [Google Scholar] [CrossRef] [PubMed]
- Amdam, G.V.; Aase, A.L.T.; Seehuus, S.-C.; Fondrk, M.K.; Norberg, K.; Hartfelder, K. Social reversal of immunosenescence in honey bee workers. Exp. Gerontol. 2005, 40, 939–947. [Google Scholar] [CrossRef]
- Shoba, S.P.; Mary, J.V.J.; Anitha, C.; Punitha, A.M. Role of Vitellogenin as Immunocompetent Molecule. In Vitellogenin in Fishes—Diversification, Biological Properties, and Future Perspectives; Springer: Singapore, 2023; pp. 93–103. [Google Scholar]
- Seehuus, S.-C.; Norberg, K.; Gimsa, U.; Krekling, T.; Amdam, G.V. Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proc. Natl. Acad. Sci. USA 2006, 103, 962–967. [Google Scholar] [CrossRef]
- Nelson, C.M.; Ihle, K.E.; Fondrk, M.K.; Page, R.E., Jr.; Amdam, G.V. The gene vitellogenin has multiple coordinating effects on social organization. PLoS Biol. 2007, 5, e62. [Google Scholar] [CrossRef]
- Crailsheim, K. The protein balance of the honey bee worker. Apidologie 1990, 21, 417–429. [Google Scholar] [CrossRef]
- Musila, J.; Přidal, A. Seasonal Changes in Hemolymph Protein Level and Hypopharyngeal Gland Size Depending on Age and In-Nest Location of Honeybee Workers. Animals 2024, 14, 512. [Google Scholar] [CrossRef]
- Lim, S.; Jung, J.; Yunusbaev, U.; Ilyasov, R.; Kwon, H.W. Characterization and its implication of a novel taste receptor detecting nutrients in the honey bee, Apis mellifera. Sci. Rep. 2019, 9, 11620. [Google Scholar] [CrossRef]
- Wenninger, E.J.; Landolt, P.J. Apple and sugar feeding in adult codling moths, Cydia pomonella: Effects on longevity, fecundity, and egg fertility. J. Insect Sci. 2011, 11, 161. [Google Scholar] [CrossRef]
- Cho, E.S.; Kim, H.J.; Lee, J.H.; Hwang, C.Y.; Siziya, I.N.; Kwon, H.W.; Seo, M.J. Intestinal microbial diversity in domestic honey bees and evaluation of their probiotic properties. J. Apic. Res. 2024, 1–12. [Google Scholar] [CrossRef]
- Noordyke, E.R.; van Santen, E.; Ellis, J.D. Tracing the Fate of Pollen Substitute Patties in Western Honey Bee (Hymenoptera: Apidae) Colonies. J. Econ. Entomol. 2021, 114, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Tawfik, A.I.; Ahmed, Z.H.; Abdel-Rahman, M.; Moustafa, A.M. Influence of winter feeding on colony development and the antioxidant system of the honey bee, Apis mellifera. J. Apic. Res. 2020, 59, 752–763. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Chen, Y.; Rivera, R.; Carroll, M.; Chambers, M.; Hidalgo, G.; de Jong, E.W. Honey bee colonies provided with natural forage have lower pathogen loads and higher overwinter survival than those fed protein supplements. Apidologie 2016, 47, 186–196. [Google Scholar] [CrossRef]
- Omar, E.M.; Darwish, H.Y.A.; Othman, A.A.; El-Seedi, H.R.; Al Naggar, Y. Crushing corn pollen grains increased diet digestibility and hemolymph protein content while decreasing honey bee consumption. Apidologie 2022, 53, 52. [Google Scholar] [CrossRef]
- Ketnawa, S.; Ogawa, Y. Evaluation of protein digestibility of fermented soybeans and changes in biochemical characteristics of digested fractions. J. Funct. Foods 2019, 52, 640–647. [Google Scholar] [CrossRef]
- Slansky, F.; Scriber, J.M. Food consumption and utilization. In Comparative Insect Physiology, Biochemistry, and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon: New York, NY, USA, 1985; Volume 4, pp. 87–163. [Google Scholar]
- Lee, J.H.; Kim, H.-W.; Mustafa, B.; Lee, H.I.; Kwon, H.W. The relationships between microbiome diversity and epidemiology in domestic species of malaria-mediated mosquitoes of Korea. Sci. Rep. 2023, 13, 9081. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Dada, S.H. High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Maigoro, A.; Lee, S. Gut microbiome-based analysis of lipid a biosynthesis in individuals with autism spectrum disorder: An in silico evaluation. Nutrients 2021, 13, 688. [Google Scholar] [CrossRef] [PubMed]
- Krams, I.A.; Kecko, S.; Jõers, P.; Trakimas, G.; Elferts, D.; Krams, R.; Luoto, S.; Rantala, M.J.; Inashkina, I.; Gudrā, D.; et al. Microbiome symbionts and diet diversity incur costs on the immune system of insect larvae. J. Exp. Biol. 2017, 220, 4204–4212. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.-H.; Roh, S.W.; Whon, T.W.; Jung, M.-J.; Kim, M.-S.; Park, D.-S.; Yoon, C.; Nam, Y.-D.; Kim, Y.-J.; Choi, J.-H.; et al. Insect gut bacterial diversity determined by environmental habitat, diet, developmental stage, and phylogeny of host. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef] [PubMed]
- Montagna, M.; Mereghetti, V.; Gargari, G.; Guglielmetti, S.; Faoro, F.; Lozzia, G.; Locatelli, D.; Limonta, L. Evidence of a bacterial core in the stored products pest Plodia interpunctella: The influence of different diets. Environ. Microbiol. 2016, 18, 4961–4973. [Google Scholar] [CrossRef]
- Chandler, J.A.; Morgan Lang, J.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial communities of diverse Drosophila species: Ecological context of a host–microbe model system. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef]
- Pérez-Cobas, A.E.; Maiques, E.; Angelova, A.; Carrasco, P.; Moya, A.; Latorre, A. Diet shapes the gut microbiota of the omnivorous cockroach Blattella germanica. FEMS Microbiol. Ecol. 2015, 91, fiv022. [Google Scholar] [CrossRef]
- Kwong, W.K.; Moran, N.A. Cultivation and characterization of the gut symbionts of honey bees and bumble bees: Description of Snodgrassella alvi gen. nov., sp. nov., a member of the family Neisseriaceae of the Betaproteobacteria, and Gilliamella apicola gen. nov., sp. nov., a member of Orbaceae fam. nov., Orbales ord. nov., a sister taxon to the order ‘Enterobacteriales’ of the Gammaproteobacteria. Int. J. Syst. Evol. Microbiol. 2013, 63, 2008–2018. [Google Scholar] [PubMed]
- Ellegaard, K.M.; Engel, P. Genomic diversity landscape of the honey bee gut microbiota. Nat. Commun. 2019, 10, 446. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.-R.; Geng, L.-L.; Li, Y.-H.; Wang, Q.; Diao, Q.-Y.; Zhou, T.; Dai, P.-L. The effects of Bt Cry1Ie toxin on bacterial diversity in the midgut of Apis mellifera ligustica (Hymenoptera: Apidae). Sci. Rep. 2016, 6, 24664. [Google Scholar] [CrossRef]
- Gkantiragas, A.G.; Gabrielli, J.A. Meta-Analysis of the 16S-rRNA Gut Microbiome Data in Honeybees (Apis Mellifera). bioRxiv 2021. [Google Scholar] [CrossRef]
- Vásquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E.; Szekely, L.; Olofsson, T.C. Symbionts as major modulators of insect health: Lactic acid bacteria and honeybees. PLoS ONE 2012, 7, e33188. [Google Scholar] [CrossRef]
- Bauer, S.; Tholen, A.; Overmann, J.; Brune, A. Characterization of abundance and diversity of lactic acid bacteria in the hindgut of wood-and soil-feeding termites by molecular and culture-dependent techniques. Arch. Microbiol. 2000, 173, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, J.; Wang, X.; Jin, Z.; Zhang, P.; Su, H.; Sun, X. Effect of probiotics on respiratory tract allergic disease and gut microbiota. Front. Nutr. 2022, 9, 821900. [Google Scholar] [CrossRef] [PubMed]
- Daisley, B.A.; Pitek, A.P.; Chmiel, J.A.; Gibbons, S.; Chernyshova, A.M.; Al, K.F.; Faragalla, K.M.; Burton, J.P.; Thompson, G.J.; Reid, G. Lactobacillus spp. attenuate antibiotic-induced immune and microbiota dysregulation in honey bees. Commun. Biol. 2020, 3, 534. [Google Scholar] [CrossRef]
- Vásquez, A.; Olofsson, T.C.; Sammataro, D.J.A. A scientific note on the lactic acid bacterial flora in honeybees in the USA–A comparison with bees from Sweden. Apidologie 2009, 40, 26–28. [Google Scholar] [CrossRef]
- Li, L.; Solvi, C.; Zhang, F.; Qi, Z.; Chittka, L.; Zhao, W. Gut microbiome drives individual memory variation in bumblebees. Nat. Commun. 2021, 12, 6588. [Google Scholar] [CrossRef]
- Huang, Y.; Li, N.; Yang, C.; Lin, Y.; Wen, Y.; Zheng, L.; Zhao, C. Honeybee as a food nutrition analysis model of neural development and gut microbiota. Neurosci. Biobehav. Rev. 2023, 153, 105372. [Google Scholar] [CrossRef]
- Coenye, T.; Vandamme, P. Extracting phylogenetic information from whole-genome sequencing projects: The lactic acid bacteria as a test case. Microbiology 2003, 149, 3507–3517. [Google Scholar] [CrossRef]
- Anderson, K.E.; Johansson, A.; Sheehan, T.H.; Mott, B.M.; Corby-Harris, V.; Johnstone, L.; Sprissler, R.; Fitz, W. Draft genome sequences of two Bifidobacterium sp. from the honey bee (Apis mellifera). Gut Pathog. 2013, 5, 42. [Google Scholar] [CrossRef]
- Butler, È.; Alsterfjord, M.; Olofsson, T.C.; Karlsson, C.; Malmström, J.; Vásquez, A. Proteins of novel lactic acid bacteria from Apis mellifera mellifera: An insight into the production of known extra-cellular proteins during microbial stress. BMC Microbiol. 2013, 13, 235. [Google Scholar] [CrossRef]
- Hamdi, C.; Balloi, A.; Essanaa, J.; Crotti, E.; Gonella, E.; Raddadi, N.; Ricci, I.; Boudabous, A.; Borin, S.; Manino, A.; et al. Gut microbiome dysbiosis and honeybee health. J. Appl. Entomol. 2011, 135, 524–533. [Google Scholar] [CrossRef]
- Olofsson, T.C.; Vásquez, A. Detection and identification of a novel lactic acid bacterial flora within the honey stomach of the honeybee Apis mellifera. Curr. Microbiol. 2008, 57, 356–363. [Google Scholar] [CrossRef]
- Fu, J.; Wang, J.; Huang, X.; Guan, B.; Feng, Q.; Deng, H. Composition and diversity of gut microbiota across developmental stages of Spodoptera frugiperda and its effect on the reproduction. Front. Microbiol. 2023, 14, 1237684. [Google Scholar] [CrossRef]
- Shamjana, U.; Vasu, D.A.; Hembrom, P.S.; Nayak, K.; Grace, T. The role of insect gut microbiota in host fitness, detoxification and nutrient supplementation. Antonie Leeuwenhoek 2024, 117, 71. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, S.-W.; Zeng, Z.-J.; Yan, W.-Y. Nutrition affects longevity and gene expression in honey bee (Apis mellifera) workers. Apidologie 2014, 45, 618–625. [Google Scholar] [CrossRef]
- Raikhel, A.S.; Dhadialla, T. Accumulation of yolk proteins in insect oocytes. Annu. Rev. Entomol. 1992, 37, 217–251. [Google Scholar]
- Aurori, C.M.; Buttstedt, A.; Dezmirean, D.S.; Mărghita¸s, L.A.; Moritz, R.F.; Erler, S. What is the main driver of aging in long-lived winter honeybees: Antioxidant enzymes, innate immunity, or vitellogenin? J. Gerontol. 2014, 69, 633–639. [Google Scholar] [CrossRef]
- Corona, M.; Velarde, R.A.; Remolina, S.; Moran-Lauter, A.; Wang, Y.; Hughes, K.A.; Robinson, G.E. Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. USA 2007, 104, 7128–7133. [Google Scholar] [CrossRef]
- Gemeda, T.K. Testing the effect of dearth period supplementary feeding of honeybee (Apis mellifera) on brood development and honey production. Int. J. Adv. Res. 2014, 2, 319–324. [Google Scholar]
- Horton, M.A.; Oliver, R.; Newton, I.L. No apparent correlation between honey bee forager gut microbiota and honey production. PeerJ 2015, 3, e1329. [Google Scholar] [CrossRef]
- D’Alvise, P.; Bohme, F.; Codrea, M.C.; Seitz, A.; Nahnsen, S.; Binzer, M.; Rosenkranz, P.; Hasselmann, M. The impact of winter feed type on intestinal microbiota and parasites in honey bees. Apidologie 2018, 49, 252–264. [Google Scholar] [CrossRef]
- Bleau, N.; Bouslama, S.; Giovenazzo, P.; Derome, N. Dynamics of the honeybee (Apis mellifera) gut microbiota throughout the overwintering period in Canada. Microorganisms 2020, 8, 1146. [Google Scholar] [CrossRef]
- Taylor, M. Microbiota in the Honey Bee Gut and Their Association with Bee Health. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 2020. [Google Scholar]
- Cornet, L.; Cleenwerck, I.; Praet, J.; Leonard, R.R.; Vereecken, N.J.; Michez, D.; Smagghe, G.; Baurain, D.; Vandamme, P. Phylogenomic analyses of Snodgrassella isolates from honeybees and bumblebees reveal taxonomic and functional diversity. mSystems 2022, 7, e01500-21. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Diet1 | Diet2 | Diet3 | Diet4 |
|---|---|---|---|---|
| Brewer’s yeast | 39.69 | 39.69 | 39.69 | 39.69 |
| Egg yolk | 2.21 | 2.21 | 2.21 | 2.21 |
| Defatted soybean powder | 2.21 | - | - | - |
| Sugar | 35.36 | 35.36 | 35.36 | 35.36 |
| Boiled water | 7.16 | 7.16 | 7.16 | 5.16 |
| Canola oil | 1.01 | 1.01 | 1.01 | 1.01 |
| Cellulose | 0.88 | 0.88 | 0.88 | 0.88 |
| Wheat bran powder | 0.88 | 0.88 | 0.88 | 0.88 |
| Multiple vitamins | 0.44 | 0.44 | 0.44 | 0.44 |
| L-methionine | 0.10 | 0.10 | 0.10 | 0.10 |
| L-lysine | 0.24 | 0.24 | 0.24 | 0.24 |
| Citric acid | 1.85 | 1.85 | 1.85 | 1.85 |
| IMP | 0.00 | 0.00 | 0.00 | 0.00 |
| GMP | 0.00 | 0.00 | 0.00 | 0.00 |
| Tangerine juice | 4.00 | 4.00 | 4.00 | 10.00 |
| Soytide powder | - | - | 2.21 | 2.21 |
| Apple juice | 4.00 | 4.00 | 4.00 | - |
| Chlorella powder | - | 0.08 | 0.08 | - |
| M | - | 2.21 | - | - |
| Group 1 | Group 2 | p-Value |
|---|---|---|
| Control (n = 3) | Diet2 (n = 3) | 0.049 |
| Diet2 (n = 3) | Megabee (n = 3) | 0.049 |
| Diet2 (n = 3) | Diet3 (n = 3) | 0.049 |
| Diet2 (n = 3) | Dietl (n = 3) | 0.049 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Maigoro, A.Y.; Lee, J.-H.; Frunze, O.; Kwon, H.-W. The Improving Effects of Probiotic-Added Pollen Substitute Diets on the Gut Microbiota and Individual Health of Honey Bee (Apis mellifera L.). Microorganisms 2024, 12, 1567. https://doi.org/10.3390/microorganisms12081567
Kim H, Maigoro AY, Lee J-H, Frunze O, Kwon H-W. The Improving Effects of Probiotic-Added Pollen Substitute Diets on the Gut Microbiota and Individual Health of Honey Bee (Apis mellifera L.). Microorganisms. 2024; 12(8):1567. https://doi.org/10.3390/microorganisms12081567
Chicago/Turabian StyleKim, Hyunjee, Abdulkadir Yusif Maigoro, Jeong-Hyeon Lee, Olga Frunze, and Hyung-Wook Kwon. 2024. "The Improving Effects of Probiotic-Added Pollen Substitute Diets on the Gut Microbiota and Individual Health of Honey Bee (Apis mellifera L.)" Microorganisms 12, no. 8: 1567. https://doi.org/10.3390/microorganisms12081567
APA StyleKim, H., Maigoro, A. Y., Lee, J.-H., Frunze, O., & Kwon, H.-W. (2024). The Improving Effects of Probiotic-Added Pollen Substitute Diets on the Gut Microbiota and Individual Health of Honey Bee (Apis mellifera L.). Microorganisms, 12(8), 1567. https://doi.org/10.3390/microorganisms12081567