
In Silico Evaluation of Lawsonia intracellularis Genes Orthologous to Genes Associated with Pathogenesis in Other Intracellular Bacteria
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Comparison of Lawsonia intracellularis Genomes Available at GenBank Database
2.2. Comparison of Orthologous Genes In Silico and Expression Assessment
2.3. Comparison of Orthologous Genes Observed in Other Enteropathogens Related to Genes Expressed in Pathogenic and Non-Pathogenic Strains of Lawsonia intracellularis
3. Results
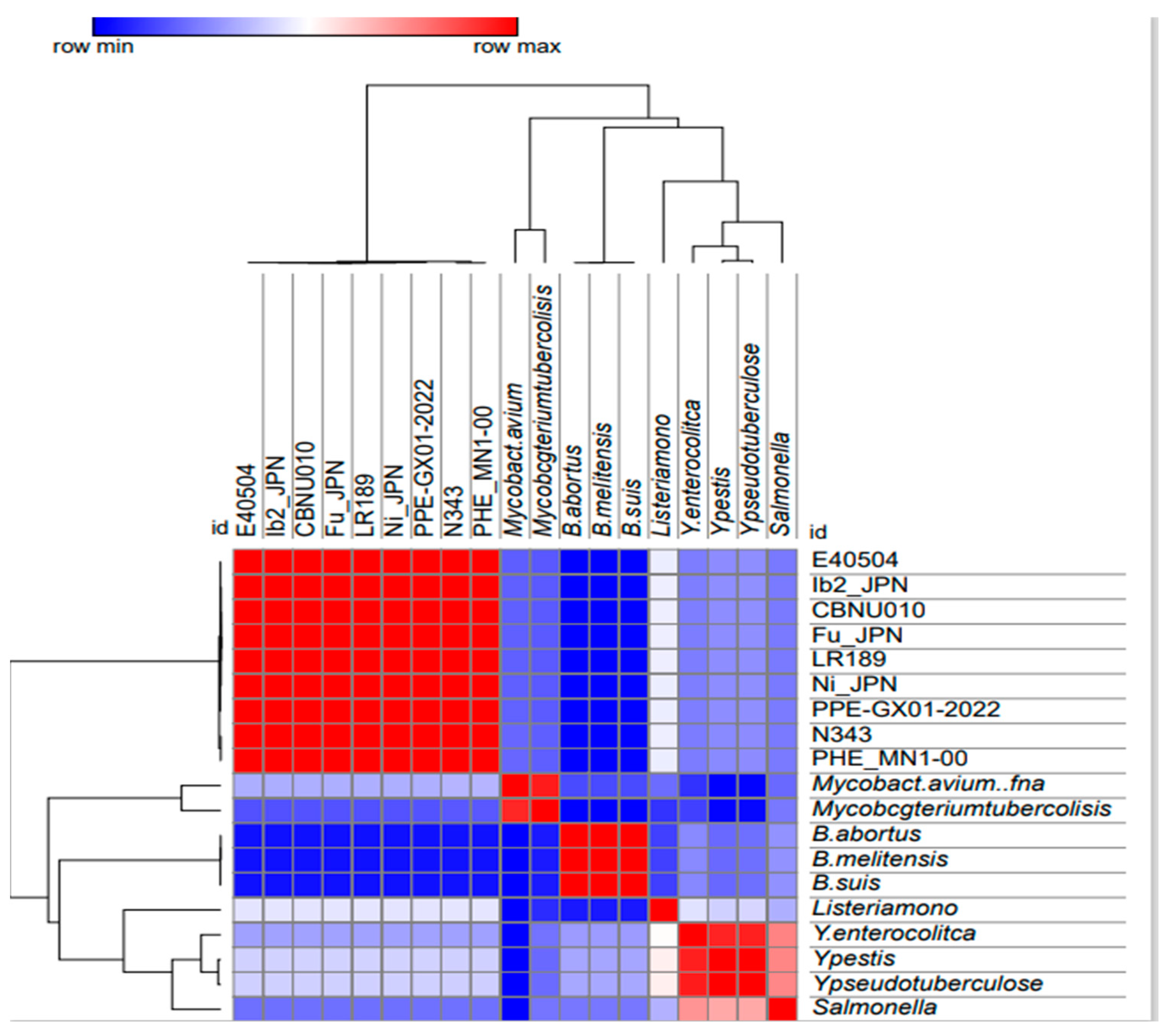
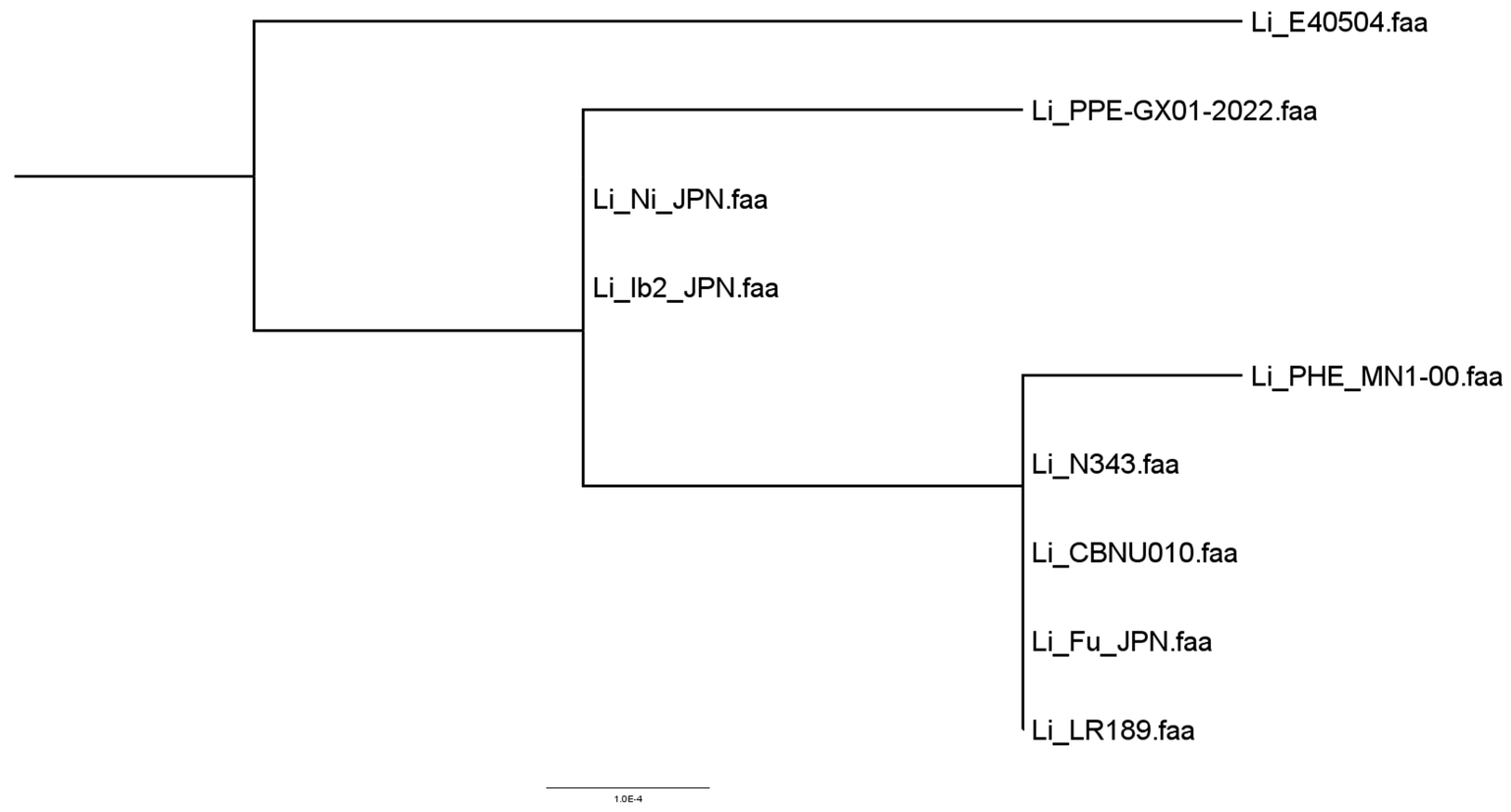
3.1. Similarity Assessment of L. intracellularis Genomes in GenBank
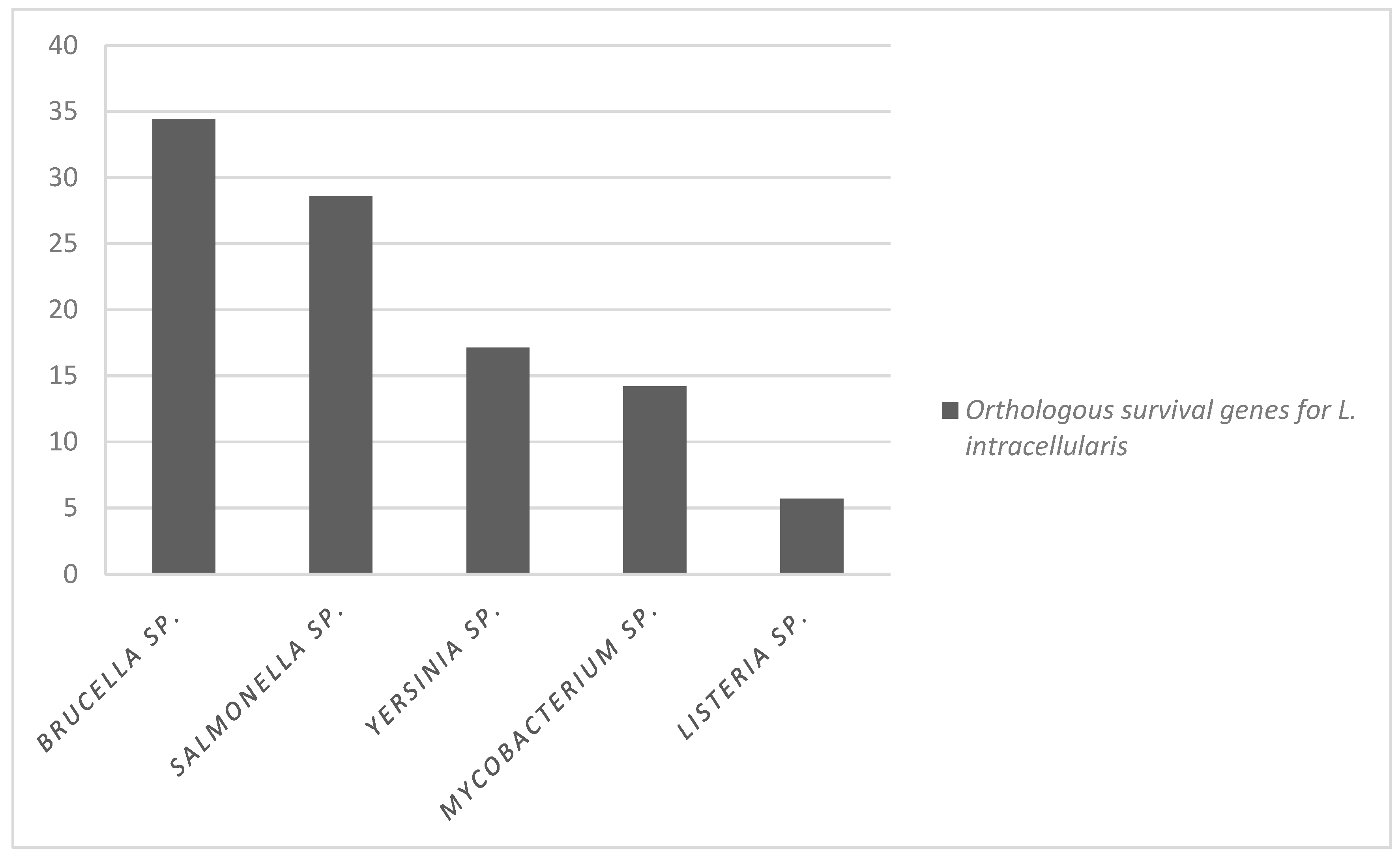
3.2. L. intracellularis Orthologous Genes to Genes Associated with Pathogenesis in Other Intracellular Bacteria
3.3. Results of Concatenated and Phylogenetically Analyzed Genes
3.4. Comparison Results of Orthologous Genes Observed in Other Enteropathogens Related to Genes Expressed in Pathogenic and Non-Pathogenic Strains of Lawsonia intracellularis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McOrist, S.; Gebhart, C.J.; Boid, R.; Barns, S.M. Characterization of Lawsonia intracellularis gen. nov., sp. nov., the obligately intracellular bacterium of porcine proliferative enteropathy. Int. J. Syst. Bacteriol. 1995, 45, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Rowland, A.C.; Lawson, G.H. Intestinal adenomatosis in the pig: Immunofluorescent and electron microscopic studies. Res. Vet. Sci. 1974, 17, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Lawson, G.H.K.; McOrist, S.; Jasni, S.; Mackie, R.A. Intracellular bacteria of porcine proliferative enteropathy: Cultivation and maintenance in vitro. J. Clin. Microbiol. 1993, 31, 1136–1142. [Google Scholar] [CrossRef] [PubMed]
- Lawson, G.H.; Gebhart, C.J. Proliferative enteropathy. J. Comp. Pathol. 2000, 122, 77–100. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, R.J.; Wee, B.A.; Yebra, G.; Bacigalupe, R.; Watson, E.; Guedes, R.M.C.; Jacobson, M.; Stadejek, T.; Archibald, A.L.; Fitzgerald, J.R.; et al. Metagenomic sequencing of clinical samples reveals a single widespread clone of Lawsonia intracellularis responsible for porcine proliferative enteropathy. Microb. Genom. 2020, 6, e000358. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.M.; Swanson, D.L.; Barns, S.M.; Gebhart, C.J. Comparison of the 16S ribosomal DNA sequences from the intracellular agents of proliferative enteritis in a hamster, deer, and ostrich with the sequence of a porcine isolate of Lawsonia intracellularis. Int. J. Syst. Bacteriol. 1997, 47, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.; Thaiss, C.A.; Zeevi, D.; Dohnalová, L.; Zilberman-Schapira, G.; Mahdi, J.A.; David, E.; Savidor, A.; Korem, T.; Herzig, Y.; et al. Microbiota-Modulated Metabolites Shape the Intestinal Microenvironment by Regulating NLRP6 Inflammasome Signaling. Cell 2015, 163, 1428–1443. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Zhang, Q.; Beckler, D.; Munir, S.; Li, L.; Kinsley, K.; Herron, L.; Peterson, A.; May, B.; Singh, S.; et al. The Complete Genome Sequence of Lawsonia intracellularis: The Causative Agent of Proliferative Enteropathy. Available online: https://www.ncbi.nlm.nih.gov/nuccore/AM180252.1 (accessed on 10 July 2022).
- Sait, M.; Aitchison, K.; Wheelhouse, N.; Wilson, K.; Lainson, F.A.; Longbottom, D.; Smith, D.G.E. Genome sequence of Lawsonia intracellularis strain N343, isolated from a sow with hemorrhagic proliferative enteropathy. Genome Announc. 2013, 1, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.R.G.C. Differentiation of Lawsonia intracellularis Isolates from Different Animal Species Using Pulsed Field Gel Electrophoresis (PFGE). U.S. Patent Application No. 2014/0017268 A1, 16 January 2014. [Google Scholar]
- Sampieri, F.; Vannucci, F.A.; Allen, A.L.; Pusterla, N.; Antonopoulos, A.J.; Ball, K.R.; Thompson, J.; Dowling, P.M.; Hamilton, D.L.; Gebhart, C.J. Species-specificity of equine and porcine Lawsonia intracellularis isolates in laboratory animals. Can. J. Vet. Res. 2013, 77, 261–272. [Google Scholar]
- Vannucci, F.A.; Beckler, D.; Pusterla, N.; Mapes, S.M.; Gebhart, C.J. Attenuation of virulence of Lawsonia intracellularis after in vitro passages and its effects on the experimental reproduction of porcine proliferative enteropathy. Vet. Microbiol. 2013, 162, 265–269. [Google Scholar] [CrossRef]
- Worley, M.J. Immune evasion and persistence in enteric bacterial pathogens. Gut Microbes 2023, 15, 2163839. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, F.A.; Pusterla, N.; Mapes, S.M.; Gebhart, C. Evidence of host adaptation in Lawsonia intracellularis infections. Vet. Res. 2012, 43, 53. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, F.A.; Foster, D.N.; Gebhart, C.J. Comparative Transcriptional Analysis of Homologous Pathogenic and Non-Pathogenic Lawsonia intracellularis Isolates in Infected Porcine Cells. PLoS ONE 2012, 7, e46708. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI reference sequences (RefSeq): A curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2007, 35, D61–D65. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.; Glover, R.; Humphris, S.; Elphinstone, J.; Toth, I. Genomics and Taxonomy in Diagnostics for Food Security: Soft-rotting enterobacterial plant pathogens. Anal. Methods 2015, 8, 12–24. [Google Scholar] [CrossRef]
- Bateman, A.; Martin, M.J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bursteinas, B.; et al. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed]
- Hickey, T.B.M.; Ziltener, H.J.; Speert, D.P.; Stokes, R.W. Mycobacterium tuberculosis employs Cpn60.2 as an adhesin that binds CD43 on the macrophage surface. Cell Microbiol. 2010, 12, 1634–1647. [Google Scholar] [CrossRef]
- Alberdi, M.P.; Watson, E.; McAllister, G.E.; Harris, J.D.; Paxton, E.A.; Thomson, J.R.; Smith, D.G. Expression by Lawsonia intracellularis of type III secretion system components during infection. Vet. Microbiol. 2009, 139, 298–303. [Google Scholar] [CrossRef]
- Homma, M.; DeRosier, D.J.; Macnab, R.M. Flagellar hook and hook-associated proteins of Salmonella typhimurium and their relationship to other axial components of the flagellum. J. Mol. Biol. 1990, 213, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Kinoshita, M.; Kida, M.; Takekawa, N.; Namba, K.; Imada, K.; Minamino, T. The FlhA linker mediates flagellar protein export switching during flagellar assembly. Commun. Biol. 2021, 4, 646. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Iino, T.; Kutuskake, K. Molecular characterization of the Salmonella typhimurium flhB operon and its protein products. J. Bacteriol. 1994, 176, 7630–7637. [Google Scholar] [CrossRef] [PubMed]
- Hazelbauer, G.L.; Falke, J.J.; Parkinson, J.S. Bacterial chemoreceptors: High-performance signaling in networked arrays. Trends Biochem. Sci. 2008, 33, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Stock, A.; Mottonen, J.; Chen, T.; Stock, J. Identification of a possible nucleotide binding site in CheW, a protein required for sensory transduction in bacterial chemotaxis. J. Biol. Chem. 1987, 262, 535–537. [Google Scholar] [CrossRef] [PubMed]
- Freer, E.; Moreno, E.; Moriyón, I.; Pizarro-Cerdá, J.; Weintraub, A.; Gorvel, J.P. Brucella-Salmonella lipopolysaccharide chimeras are less permeable to hydrophobic probes and more sensitive to cationic peptides and EDTA than are their native Brucella sp. counterparts. J. Bacteriol. 1996, 178, 5867–5876. [Google Scholar] [CrossRef] [PubMed]
- López-Goñi, I.; Guzmán-Verri, C.; Manterola, L.; Sola-Landa, A.; Moriyón, I.; Moreno, E. Regulation of Brucella virulence by the two-component system BvrR/BvrS. Vet. Microbiol. 2002, 90, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Hickey, T.B.M.; Thorson, L.M.; Speert, D.P.; Daffé, M.; Stokes, R.W. Mycobacterium tuberculosis Cpn60.2 and DnaK are located on the bacterial surface, where Cpn60.2 facilitates efficient bacterial association with macrophages. Infect. Immun. 2009, 77, 3389–3401. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, M.; Sia, J.K.; Bizzell, E.; Madan-Lala, R.; Rengarajan, J. Mycobacterium tuberculosis GroEL2 Modulates Dendritic Cell Responses. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.E.R.; Resende, T.P.; Armién, A.G.; Laub, R.P.; Vannucci, F.A.; Santos, R.L.; Gebhart, C.J.; Guedes, R.M.C. Survival of Lawsonia intracellularis in porcine peripheral blood monocyte-derived macrophages. PLoS ONE 2020, 15, e0236887. [Google Scholar] [CrossRef]
- Miller, S.I.; Kukral, A.M.; Mekalanos, J.J. A two-component regulatory system (phoP phoQ) controls Salmonella typhimurium virulence. Proc. Natl. Acad. Sci. USA 1989, 86, 5054–5058. [Google Scholar] [CrossRef] [PubMed]
- Sanowar, S.; Le Moual, H. Functional reconstitution of the Salmonella typhimurium PhoQ histidine kinase sensor in proteoliposomes. Biochem. J. 2005, 390 Pt 3, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.E.; Matsunaga, J.; Simons, E.L.; Simons, R.W. Structure and regulation of the Salmonella typhimurium rnc-era-recO operon. Biochimie 1996, 78, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Chain, P.S.G.; Carniel, E.; Larimer, F.W.; Lamerdin, J.; Stoutland, P.O.; Regala, W.M.; Georgescu, A.M.; Vergez, L.M.; Land, M.L.; Motin, V.L.; et al. Insights into the evolution of Yersinia pestis through whole-genome comparison with Yersinia pseudotuberculosis. Proc. Natl. Acad. Sci. USA 2004, 101, 13826–13831. [Google Scholar] [CrossRef]
- DelVecchio, V.G.; Kapatral, V.; Redkar, R.J.; Patra, G.; Mujer, C.; Los, T.; Ivanova, N.; Anderson, I.; Bhattacharyya, A.; Lykidis, A.; et al. The genome sequence of the facultative intracellular pathogen Brucella melitensis. Proc. Natl. Acad. Sci. USA 2002, 99, 443–448. [Google Scholar] [CrossRef]
- Glaser, P.; Frangeul, L.; Buchrieser, C.; Rusniok, C.; Amend, A.; Baquero, F.; Berche, P.; Bloecker, H.; Brandt, P.; Chakraborty, T.; et al. Comparative genomics of Listeria species. Science 2001, 294, 849–852. [Google Scholar] [CrossRef]
- Zheng, H.; Lu, L.; Wang, B.; Pu, S.; Zhang, X.; Zhu, G.; Shi, W.; Zhang, L.; Wang, H.; Wang, S.; et al. Genetic basis of virulence attenuation revealed by comparative genomic analysis of Mycobacterium tuberculosis strain H37Ra versus H37Rv. PLoS ONE 2008, 3, e2375. [Google Scholar] [CrossRef]
- Bagg, A.; Neilands, J.B. Ferric uptake regulation protein acts as a repressor, employing iron (II) as a cofactor to bind the operator of an iron transport operon in Escherichia coli. Biochemistry 1987, 26, 5471–5477. [Google Scholar] [CrossRef]
- Willett, J.W.; Herrou, J.; Briegel, A.; Rotskoff, G.; Crosson, S. Structural asymmetry in a conserved signaling system that regulates division, replication, and virulence of an intracellular pathogen. Proc. Natl. Acad. Sci. USA 2015, 112, E3709–E3718. [Google Scholar] [CrossRef]
- Dozot, M.; Boigegrain, R.-A.; Delrue, R.-M.; Hallez, R.; Ouahrani-Bettache, S.; Danese, I.; Letesson, J.-J.; De Bolle, X.; Köhler, S. The stringent response mediator Rsh is required for Brucella melitensis and Brucella suis virulence, and for expression of the type IV secretion system virB. Cell Microbiol. 2006, 8, 1791–1802. [Google Scholar] [CrossRef]
- Snel, B.; Huynen, M.A.; Dutilh, B.E. Genome trees and the nature of genome evolution. Annu. Rev. Microbiol. 2005, 59, 191. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus_Tag | Species | Gene | Description |
|---|---|---|---|
| PHE_MN1-00_00947 | Salmonella enterica serovar Typhimurium | sctN1 (invC, spaI, spaL) | SPI-1 ATPase of type 3 secretion system |
| PHE_MN1-00_00595 | Yersinia pseudotuberculosis serotype I and Yersinia enterocolitica | sctN (yscN) | Type 3 secretion system ATPase |
| PHE_MN1-00_00605 | Yersinia enterocolitica | LCD | Low calcium response locus protein D |
| PHE_MN1-00_00585 | Yersinia enterocolitica | flhA | Flagellar biosynthesis protein FlhA |
| PHE_MN1-00_01288PHE_MN1-00_00596 | Yersinia pseudotuberculosis serotype I | sctL (lcrKC, yscL) | Type 3 secretion system stator protein |
| PHE_MN1-00_01265 | Brucella abortus | bvrR | Flagellar transcriptional regulator FtcR |
| PHE_MN1-00_00693 | Mycobacterium avium sub paratuberculosis, Mycobacterium tuberculosis. | groEL1,2 (groL2, hsp65, mopA). | Chaperonin GroEL 1,2 |
| PHE_MN1-00_01294 | Salmonella enterica sorovar Typhimurium | cheW | CheW protein chemotaxis |
| PHE_MN1-00_00296 | Brucella melitensis. Listeria monocytogenes serovar 1/2a | rho | Rho transcription termination factor |
| PHE_MN1-00_00828 | Salmonella enterica sorovar Typhimurium | flgK (flaS, flaW) | Flagellar hook-associated protein 1 |
| PHE_MN1-00_00510 | Yersinia enterocolitica | yadB (gluQ) (unreviewed) | Glutamyl-Q tRNA(Asp) synthetase |
| PHE_MN1-00_00430 | Salmonella enterica sorovar Typhimurium | rsep (yaeL) | RseP sigma-E protease regulator |
| PHE_MN1-00_00972 | Salmonella enterica sorovar Typhimurium | phoQ (phoZ) | Virulence sensor histidine kinase PhoQ |
| PHE_MN1-00_01197 | Brucella abortus | bvrS (not revised) | Histidine kinase |
| PHE_MN1-00_01283 | Salmonella enterica sorovar Typhimurium | sctC2 (spiA, ssaC) | SPI-2 type 3 secretion system secretin |
| Locus_Tag. | Species | Gene | Description |
|---|---|---|---|
| PHE_MN1-00_01458 PHE_MN1-00_00396 | Mycobacterium tuberculosis., Salmonella Typhimurium | dnaB | Replicative DNA helicase |
| PHE_MN1-00_00545 | Yersinia pestis, Salmonella enterica sorovar Typhimurium, Brucella abortus, Brucella suis biovar 1. Brucella melitensis biotype 1 | ssb | Single-stranded DNA-binding protein |
| PHE_MN1-00_00974 | Salmonella enterica sorovar Typhimurium. Yersinia pseudotuberculosis serotype I | sfsA | Sugar-fermentation-stimulating protein A |
| PHE_MN1-00_00191 | Brucella melitensis biotype 1. Brucella abortus. Brucella suis biovar 1 | rsh | GTP pyrophosphokinase rsh |
| PHE_MN1-00_00172 | Brucella suis biovar 1. Brucella abortus. Brucella melitensis biotype 1, Yersinia enterocolitica. Mycobacterium tuberculosis. Salmonella enterica sorovar Typhimurium | ribH | 6,7-dimethyl-8-ribothylumazine synthase 2 |
| PHE_MN1-00_00339 | Salmonella enterica sorovar Typhimurium. Brucella melitensis biotype 1 Yersinia pseudotuberculosis. Mycobacterium tuberculosis. Listeria monocytogenes | recO | RecO DNA repair protein |
| PHE_MN1-00_00988 | Salmonella enterica sorovar Typhimurium. Brucella melitensis biotype 1 | lnt (cutE) | Apolipoprotein N-acyltransferase |
| PHE_MN1-00_00378 | Salmonella Typhimurium | livH | High-affinity branched-chain amino acid transport system permease protein |
| PHE_MN1-00_01265 | Brucella abortus | ctrA | Cell cycle response regulator CtrA |
| PHE_MN1-00_01068 | Yersinia enterocolitica. Mycobacterium tuberculosis. Brucella melitensis | rplW | 50S ribosomal protein L23 |
| PHE_MN1-00_01312 | Mycobacterium tuberculosis | eccA1 | ESX-1 secretion system protein EccA1 |
| PHE_MN1-00_00581 | Brucella abortus | cpdR | CpdR response regulator receptor protein |
| PHE_MN1-00_00498 | Brucella melitensis biotype 1 | pyrG | CTP synthase |
| PHE_MN1-00_00072 | Mycobacterium tuberculosis | eccCa1 (snm1) | ESX-1 secretion system protein EccCa1 |
| PHE_MN1-00_00278 | Salmonella Typhimurium | hypA | HypA hydrogenase maturation factor |
| PHE_MN1-00_01140 | Salmonella Typhimurium | epmA (genX, yjeA) | Elongation factor P--(R)-beta-lysine ligase |
| PHE_MN1-00_01197 | Brucella abortus | cckA | CckA sensor kinase |
| PHE_MN1-00_00036 | Brucella abortus biovar 1. Brucella melitensis biotype 1 Yersinia pestis | fur | Ferric uptake regulatory protein |
| PHE_MN1-00_01306 | Brucella abortus | recA | RecA protein |
| PHE_MN1-00_00504 | Salmonella Typhimurium | rpoN | RNA polymerase factor sigma-54 |
| Locus Tag | Orthologous Genes | Genes Highly Expressed in Pathogenic L. intracellularis | Genes Highly Expressed in Non-Pathogenic L. intracellularis |
|---|---|---|---|
| PHE_MN1-00_01294 | cheW | cheW chemotaxis signal transduction protein | ____ |
| PHE_MN1-00_00595 | sctN (yscN) | ____ | yscN type III secretion system ATPase |
| PHE_MN1-00_00828 | flgK (flaS, flaW) | ____ | flgK flagellar hook-associated protein |
| PHE_MN1-00_00693 | groEL1 and 2 (groL2, hsp65, mopA). | GroEL (Chaperonin) | GroEL (Chaperonin) |
| Locus_Tag. | Orthologous Genes | Genes Expressed in Pathogenic L. intracellularis | Genes Expressed in Non-Pathogenic L. intracellularis |
|---|---|---|---|
| PHE_MN1-00_00172 | ribH | ribH riboflavin synthase beta-chain | ____ |
| PHE_MN1-00_00378 | livH | livH branched-chain amino acid ABC transporter (permease) | ____ |
| PHE_MN1-00_01068 | rplW | rplW. 50S ribosomal protein L23 | ____ |
| PHE_MN1-00_00278 | hypA | hypA zinc finger protein | ____ |
| PHE_MN1-00_00974 | sfsA | sfsA DNA-binding protein, stimulates sugar fermentation | ____ |
| PHE_MN1-00_00339 | recO | recO DNA repair protein RecO (recombination protein O) | ____ |
| PHE_MN1-00_00036 | Fur | fur Fe2+/Zn2+ uptake regulation proteins | ____ |
| PHE_MN1-00_00504 | rpoN | rpoN Sigma54-like protein | ____ |
| Strain | Assembly Accession | Gene | Strand | Start | Stop |
|---|---|---|---|---|---|
| PHE_MN1-00 | GCF_000055945.1 | bvrR | + | 1400172 | 1400540 |
| PHE_MN1-00 | GCF_000055945.1 | cpdR | − | 644060 | 644440 |
| PHE_MN1-00 | GCF_000055945.1 | phoQ | − | 1091827 | 1093269 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suarez-Duarte, M.E.; Santos, R.L.; Pereira, C.E.R.; Resende, T.P.; Araujo, M.D.; Correia, P.A.; Barbosa, J.C.R.; Laub, R.P.; Rodrigues, D.L.N.; Aburjaile, F.F.; et al. In Silico Evaluation of Lawsonia intracellularis Genes Orthologous to Genes Associated with Pathogenesis in Other Intracellular Bacteria. Microorganisms 2024, 12, 1596. https://doi.org/10.3390/microorganisms12081596
Suarez-Duarte ME, Santos RL, Pereira CER, Resende TP, Araujo MD, Correia PA, Barbosa JCR, Laub RP, Rodrigues DLN, Aburjaile FF, et al. In Silico Evaluation of Lawsonia intracellularis Genes Orthologous to Genes Associated with Pathogenesis in Other Intracellular Bacteria. Microorganisms. 2024; 12(8):1596. https://doi.org/10.3390/microorganisms12081596
Chicago/Turabian StyleSuarez-Duarte, Mirtha E., Renato L. Santos, Carlos E. R. Pereira, Talita P. Resende, Matheus D. Araujo, Paula A. Correia, Jessica C. R. Barbosa, Ricardo P. Laub, Diego L. N. Rodrigues, Flavia F. Aburjaile, and et al. 2024. "In Silico Evaluation of Lawsonia intracellularis Genes Orthologous to Genes Associated with Pathogenesis in Other Intracellular Bacteria" Microorganisms 12, no. 8: 1596. https://doi.org/10.3390/microorganisms12081596