
The New Genus Caulinema Revealed New Insights into the Generic Relationship of the Order Ulotrichales (Ulvophyceae, Chlorophyta)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultures and Light Microscopy
- The strain CCAP 312/1 was isolated from sand grains in Kames Bay, Millport, Isle of Cumbrae, Scotland, UK (55°45′18″ N, −4°54′56″ E), by Michael Droop in 1958. This marine strain was cultivated in seawater medium (SWES, medium 5 in Schlösser [14]) at 20 °C at a light intensity of 20 µE/m2s and under a light:dark cycle of 14:10 h;
- The strain SAG 2661 was isolated from a sample collected from the Seeache by Unterach, Austria (47°48′8″ N, 13°27′1″ E), in 2021. This freshwater strain was grown in Bold’s basal medium (3N-BBM+V, medium 26a in Schlösser [15]) under the same conditions.
2.2. DNA Extraction, PCR, Sequencing, and Phylogenetic Analyses
3. Results
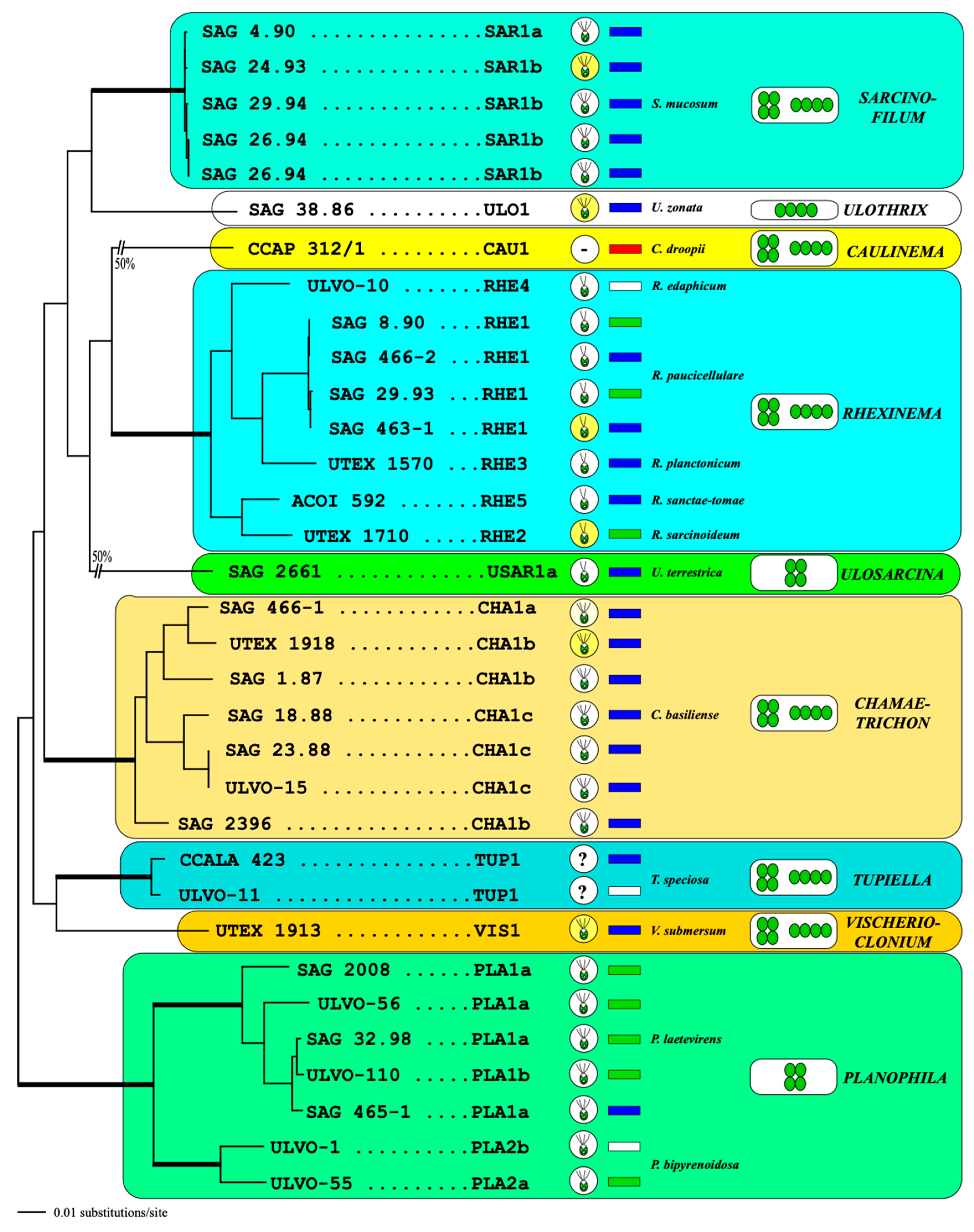
3.1. Morphology and Phenotypic Plasticity
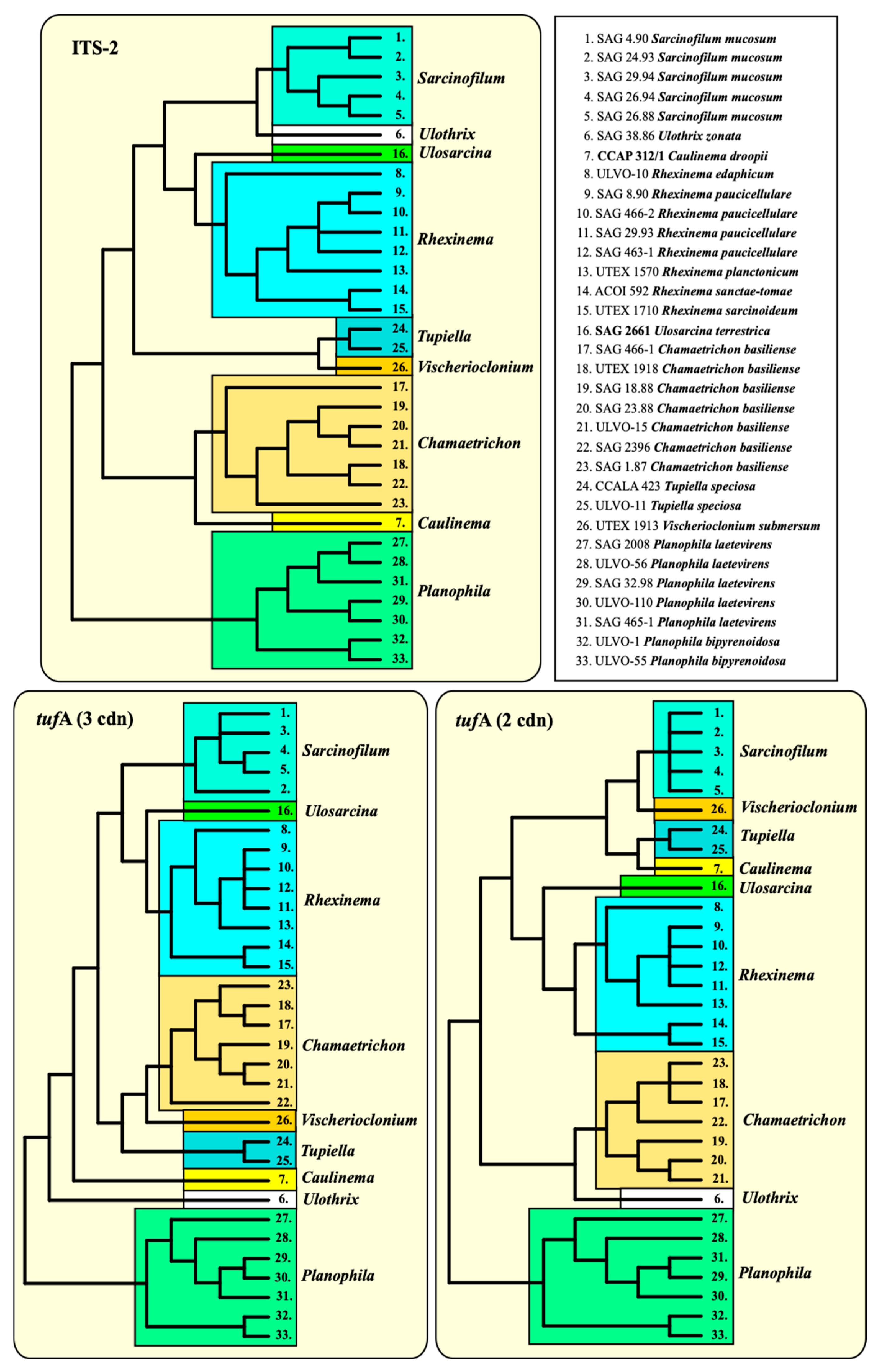
3.2. Phylogenetic Position of Both Investigated Strains
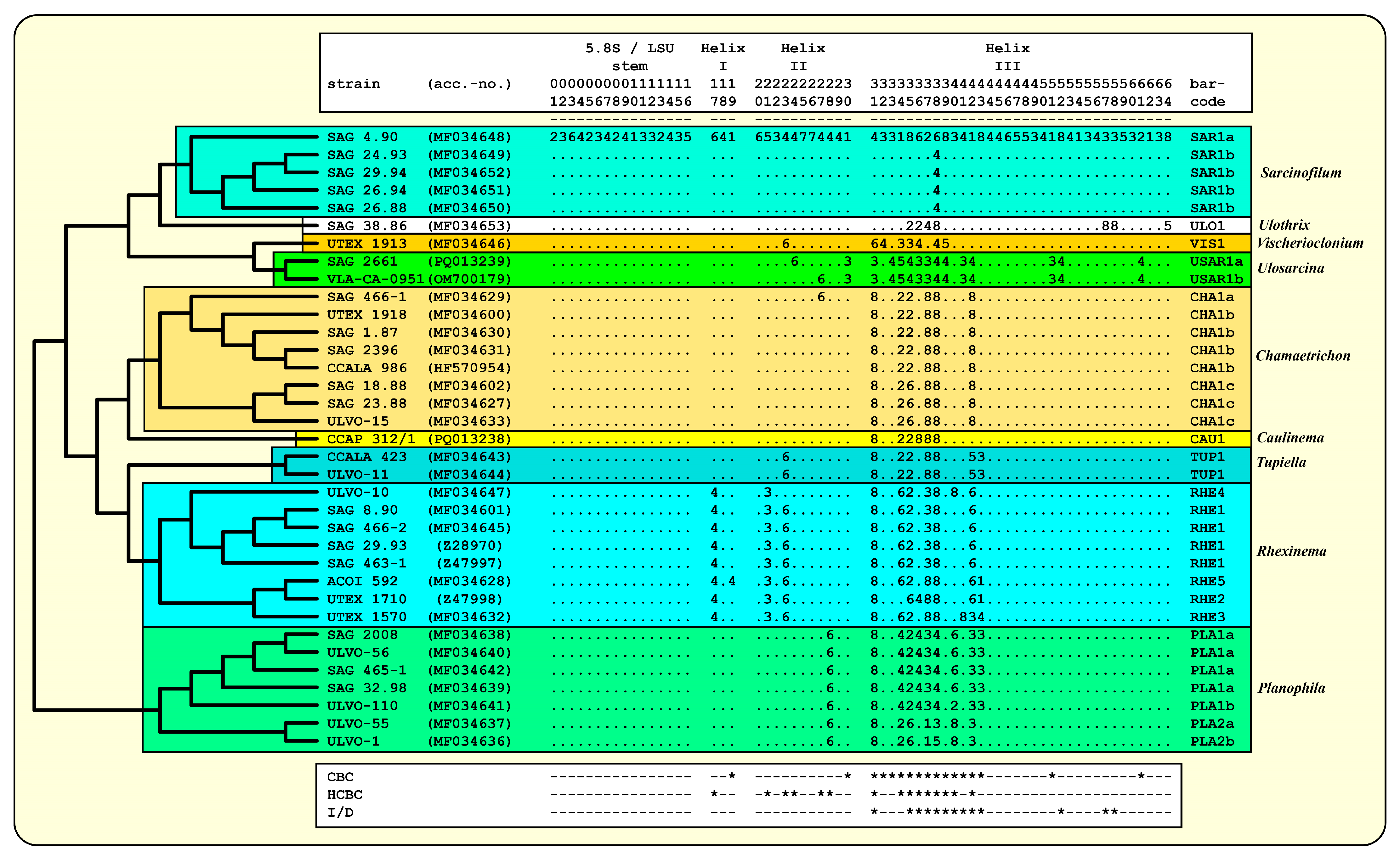
3.3. ITS-2/CBC Approach of the Ulotrichales for Species Delimitation
3.4. The Usage of the Chloroplast-Encoded Barcode Marker tufA among the Ulotrichales
4. Discussion
4.1. The Molecular Phylogeny and Systematics of the Order Ulotrichales
4.2. Taxonomic Revisions and Diagnoses
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mattox, K.R.; Stewart, K.D. Classification of the green algae: A concept based on comparative cytology. In The Systematics of Green Algae; Irvine, D.E.G., John, D.M., Eds.; Systematics Association 27; Academic Press: London, UK, 1984; pp. 29–72. [Google Scholar]
- Lokhorst, G.M. Current ideas on classification of the Ulotrichales Borzi. In The Systematics of Green Algae; Irvine, D.E.G., John, D.M., Eds.; Systematics Association 27; Academic Press: London, UK, 1984; pp. 179–206. [Google Scholar]
- O’Kelly, C.J.; Floyd, G.L. Correlations among patterns of sporangial structure and development, life histories, and ultrastructural features in the Ulvophyceae. In The Systematics of Green Algae; Irvine, D.E.G., John, D.M., Eds.; Systematics Association 27; Academic Press: London, UK, 1984; pp. 121–156. [Google Scholar]
- Kornmann, P. Die Ulotrichales, neu geordnet auf der Grundlage entwicklungsgeschichtlicher Befunde. Phycologia 1963, 3, 60–68. [Google Scholar] [CrossRef]
- Kornmann, P. Ontogenie und Lebenszyklus der Ulotrichales in phylogenetischer Sicht. Phycologia 1965, 4, 163–172. [Google Scholar] [CrossRef]
- Kornmann, P. Codiolophyceae, a new class of Chlorophyta. Helgol. Wiss. Meeresunters 1973, 25, 1–13. [Google Scholar] [CrossRef]
- Jónsson, S. Le cycle de développement du Spongomorpha lanosa (Roth) Kütz. et la nouvelle familie des Acrosiphoniacées. C. R. Hebd. Séanc. Acad. Sci. 1959, 248, 1565–1567. [Google Scholar]
- Lindstrom, S.C.; Hanic, L.A. The phylogeny of North American Urospora (Ulotrichales, Chlorophyta) based on sequence analysis of nuclear ribosomal genes, introns and spacers. Phycologia 2005, 44, 194–201. [Google Scholar] [CrossRef]
- Kornmann, P.; Sahling, P.-H. Meeresalgen von Helgoland: Ergänzung. Helgol. Meeresunters. 1983, 36, 1–65. [Google Scholar] [CrossRef]
- Darienko, T.; Rad Menéndez, C.; Campbell, C.N.; Pröschold, T. Molecular phylogeny of unicellular marine coccoid green algae revealed new insights into the systematics of the Ulvophyceae (Chlorophyta). Microorganisms 2021, 9, 1586. [Google Scholar] [CrossRef] [PubMed]
- Darienko, T.; Pröschold, T. Toward a monograph of non-marine Ulvophyceae using an integrative approach (Molecular phylogeny and systematics of terrestrial Ulvophyceae II.). Phytotaxa 2017, 324, 1–41. [Google Scholar] [CrossRef]
- Turmel, M.; Otis, C.; Lemieux, C. Divergent copies of the large inverted repeat in the chloroplast genomes of ulvophycean green algae. Sci. Rep. 2017, 7, 994. [Google Scholar] [CrossRef]
- Hou, Z.; Ma, X.; Shi, X.; Yang, L.; Xiao, S.; de Clerck, O.; Leliaert, F.; Zhong, B. Phylotranscriptomic insights into a Mesoproterozoic–Neoproterozoic origin and early radiation of green seaweeds (Ulvophyceae). Nat. Comm. 2022, 13, 1610. [Google Scholar] [CrossRef]
- Schlösser, U.G. SAG-Sammlung von Algenkulturen at the University of Göttingen. Bot. Acta 1994, 107, 113–186. [Google Scholar] [CrossRef]
- Schlösser, U.G. Additions to the culture collections of algae since 1994. Bot. Acta 1997, 110, 424–429. [Google Scholar] [CrossRef]
- Darienko, T.; Rad Menéndez, C.; Campbell, C.; Pröschold, T. Are there any true marine Chlorella species? Molecular phylogenetic assessment and ecology of marine Chlorella-like organisms, including description of Droopiella gen. nov. Syst. Biodivers. 2019, 17, 811–829. [Google Scholar] [CrossRef] [PubMed]
- Marin, B.; Palm, A.; Klingberg, M.; Melkonian, M. Phylogeny and taxonomic revision of plastid-containing euglenophytes based on SSU rDNA sequence comparison and synapomorphic signatures in the SSU rRNA secondary structure. Protist 2003, 154, 99–145. [Google Scholar] [CrossRef] [PubMed]
- Fama, P.; Wysor, B.; Kooistra, W.; Zuccarello, G.C. Molecular phylogeny of the genus Caulerpa (Caulerpales, Chlorophyta) inferred from chloroplast tufA gene. J. Phycol. 2002, 38, 1040–1050. [Google Scholar] [CrossRef]
- Saunders, G.W.; Kucera, H. An evaluation of rbcL, tufA, UPA, LSU and ITS as DNA barcode markers for the marine green macroalgae. Crypt. Algol. 2010, 31, 487–528. [Google Scholar]
- Swofford, D.L. PAUP* Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Jow, H.; Hudelot, C.; Rattray, M.; Higgs, P. Bayesian phylogenetics using an RNA substitution model applied to early mammalian evolution. Mol. Biol. Evol. 2002, 19, 1591–1601. [Google Scholar] [CrossRef]
- Higgs, P.; Jameson, D.; Jow, H.; Rattray, M. The evolution of tRNA-Leu genes in animal mitochondrial genomes. J. Mol. Evol. 2003, 57, 435–445. [Google Scholar] [CrossRef]
- Hudelot, C.; Gowri-Shankar, V.; Jow, H.; Rattray, M.; Higgs, P. RNA-based phylogenetic methods: Application to mammalian mitochondrial RNA sequences. Mol. Phylogen. Evol. 2003, 28, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Gibson, A.; Gowri-Shankar, V.; Higgs, P.; Rattray, M. A comprehensive analysis of mammalian mitochondrial genome base composition and improved phylogenetic methods. Mol. Biol. Evol. 2005, 22, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Telford, M.J.; Wise, M.J.; Gowri-Shankar, V. Consideration of RNA secondary structure significantly improves likelihood-based estimates of phylogeny: Examples from the bilateria. Mol. Biol. Evol. 2005, 22, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acid Res. 2003, 31, 3406–3615. [Google Scholar] [CrossRef] [PubMed]
- Darty, K.; Denise, A.; Ponty, Y. VARNA: Interactive drawing and editing of the RNA secondary structure. Bioinformatics 2009, 25, 1974–1975. [Google Scholar] [CrossRef] [PubMed]
- Gontcharov, A.A.; Nikulin, A.Y.; Nikulin, V.Y.; Allaguvatova, R.Z.; Bagmet, V.B.; Abdullin, S.R. Ulosarcina terrestrica gen. nov., sp. nov., a new ulvophycean sarcinoid alga from the Russian Far East. Plants 2022, 11, 3228. [Google Scholar] [CrossRef] [PubMed]
- Mai, J.C.; Coleman, A.W. The internal transcribed spacer 2 exhibits a common secondary structure in green algae and flowering plants. J. Mol. Evol. 1997, 44, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Sluiman, H.J. The green algal class Ulvophyceae. An ultrastructural survey and classification. Crypt. Bot. 1989, 1, 83–94. [Google Scholar]
- van den Hoek, C.; Stam, W.T.; Olsen, J.L. The emergence of a new chlorophytan system, and Dr. Kornmann’s contribution thereto. Helgol. Meeresunters 1988, 42, 339–383. [Google Scholar] [CrossRef]
- Van den Hoek, C.; Mann, D.G.; Jahns, H.M. Algae. An Introduction to Phycology; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Floyd, G.L.; O’Kelly, C.J. Motile cell ultrastructure and the circumscription of the orders Ulotrichales and Ulvales (Ulvophyceae, Chlorophyta). Amer. J. Bot. 1984, 71, 111–120. [Google Scholar] [CrossRef]
- Mattox, K.R.; Stewart, K.D. Observations on the zoospore of Pseudendoclonium basiliense and Trichosarcina polymorpha (Chlorophyceae). Can. J. Bot. 1973, 51, 1425–1430. [Google Scholar] [CrossRef]
- Chesnoy, L.; Jónsson, S. Etude ultrastructurale du développement du zygote calcicole d’une Chlorophycée, le Monostroma grevillei (Thuret) Wittr. C. R. Hebd. Séanc. Acad. Sci. Sér. D 1973, 276, 299–302. [Google Scholar]
- Jónsson, S.; Chesnoy, L. Etude ultrastructurale de l’incorporation des axonémes flagellaires dans les zygotes du Monostroma grevillei (Thuret) Wittr., Chlorophycée marine. C. R. Hebd. Séanc. Acad. Sci. Sér. D 1974, 278, 1537–1560. [Google Scholar]
- O’Kelly, C.J.; Floyd, G.L.; Dube, M.A. The fine structure of motile cells in the genera Ulvaria and Monostroma, with special reference to the taxonomic position of Monostroma oxyspermum (Ulvophyceae, Chlorophyta). Plant Syst. Evol. 1984, 144, 179–199. [Google Scholar] [CrossRef]
- Hoops, H.J.; Floyd, G.L.; Swanson, J.A. Ultrastructure of the biflagellate motile cells of Ulvaria oxysperma (Kütz.) Bliding and phylogenetic relationships among ulvaphycean algae. Amer. J. Bot. 1982, 69, 150–159. [Google Scholar] [CrossRef]
- Sluiman, H.J.; Roberts, K.R.; Stewart, K.D.; Mattox, K.R. Comparative cytology and taxonomy of the Ulvaphyceae. I. The zoospore of Ulothrix zonata (Chlorophyta). J. Phycol. 1980, 16, 537–545. [Google Scholar] [CrossRef]
- Sluiman, H.J.; Roberts, K.R.; Stewart, K.D.; Mattox, K.R. Comparative cytology and taxonomy of the Ulvophyceae. IV. The zoospore of Ulothrix zonata (Chlorophyta). Acta Bot. Neerl. 1983, 32, 257–269. [Google Scholar] [CrossRef]
- Lokhorst, G.M.; Star, W. The ultrastructure of Ulothrix mucosa. II. The flagellar apparatus of the zoospore. Can. J. Bot. 1986, 64, 166–176. [Google Scholar] [CrossRef]
- Nakayama, T.; Inouye, I. Ultrastructure of the biflagellate gametes of Collinsiella cava (Ulvophyceae, Chlorophyta). Phycol. Res. 2000, 48, 63–73. [Google Scholar] [CrossRef]
- Watanabe, S.; Floyd, G.L. Ultrastructure of the motile cells of the prostrate filamentous green algae Protoderma sarcinoidea and Chamaetrichon capsulatum. Plant Syst. Evol. 1992, 179, 73–87. [Google Scholar] [CrossRef]
- Friedl, T. Evolution of the polyphyletic genus Pleurastrum (Chlorophyta): Inferences from nuclear-encoded ribosomal DNA sequences and motile cell ultrastructure. Phycologia 1996, 35, 456–469. [Google Scholar] [CrossRef]
- Friedl, T.; Zeltner, C. Assessing the relationships of some coccoid green lichen algae and the Microthamniales (Chlorophyta) with 18S ribosomal RNA gene sequence comparisons. J. Phycol. 1994, 30, 500–506. [Google Scholar] [CrossRef]
- Pombert, J.F.; Otis, C.; Lemieux, C.; Turmel, M. The chloroplast genome sequence of the green alga Pseudendoclonium akinetum (Ulvophyceae) reveals unusual structural features and new insights into the branching order of chlorophyte lineages. Mol. Biol. Evol. 2005, 22, 1903–1918. [Google Scholar] [CrossRef] [PubMed]
- Carlile, A.L.; O’Kelly, C.J.; Sherwood, A.R. The green algal genus Cloniophora represents a novel lineage in the Ulvales: A proposal for Cloniophoraceae fam. nov. J. Phycol. 2011, 47, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- O’Kelly, C.J.; Wysor, B.; Bellows, W.K. Collinsiella (Ulvophyceae, Chlorophyta) and other ulotrichalean taxa with shell-boring sporophytes form a monophyletic clade. Phycologia 2004, 43, 41–49. [Google Scholar] [CrossRef]
- Friedl, T.; O’Kelly, C.J. Phylogenetic relationships of green algae assigned to the genus Planophila. Eur. J. Phycol. 2002, 37, 373–384. [Google Scholar] [CrossRef]
- Wetherbee, R.; Verbruggen, H. Kraftionema allantoideum, a new genus and family of Ulotrichales (Chlorophyta) adapted for survival in high intertidal pools. J. Phycol. 2016, 52, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Skaloud, P.; Rindi, F.; Boedecker, C.; Leliaert, F. Chlorophyta: Ulvophyceae. In Süßwasserflora von Mitteleuropa 13; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Springer Spektrum: Berlin-Heidelberg, Germany, 2018; pp. 1–288. [Google Scholar]
- Bast, F. Taxonomic reappraisal of Monostromataceae (Ulvophyceae: Chlorophyta) based on multi-locus phylogeny. Webbia 2015, 70, 43–57. [Google Scholar] [CrossRef]
- Berger-Perrot, Y.; Thomas, J.C. Étude ultrastructurale comparée du pyrénoïde et des parois dans les genres Ulothrix, Chlorothrix et Urospora. Phycologia 1982, 21, 355–369. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darienko, T.; Rad-Menéndez, C.; Pröschold, T. The New Genus Caulinema Revealed New Insights into the Generic Relationship of the Order Ulotrichales (Ulvophyceae, Chlorophyta). Microorganisms 2024, 12, 1604. https://doi.org/10.3390/microorganisms12081604
Darienko T, Rad-Menéndez C, Pröschold T. The New Genus Caulinema Revealed New Insights into the Generic Relationship of the Order Ulotrichales (Ulvophyceae, Chlorophyta). Microorganisms. 2024; 12(8):1604. https://doi.org/10.3390/microorganisms12081604
Chicago/Turabian StyleDarienko, Tatyana, Cecilia Rad-Menéndez, and Thomas Pröschold. 2024. "The New Genus Caulinema Revealed New Insights into the Generic Relationship of the Order Ulotrichales (Ulvophyceae, Chlorophyta)" Microorganisms 12, no. 8: 1604. https://doi.org/10.3390/microorganisms12081604