
Alteration in the Morphological and Transcriptomic Profiles of Acinetobacter baumannii after Exposure to Colistin
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Antimicrobial Susceptibility Testing
2.3. Multilocus Sequence Typing
2.4. Determination of Colistin-Resistance-Associated Gene Sequences
2.5. RNA Isolation and Reverse Transcription Quantitative PCR (RT-qPCR)
2.6. Whole-Genome Sequencing
2.7. Comparative Transcriptomics
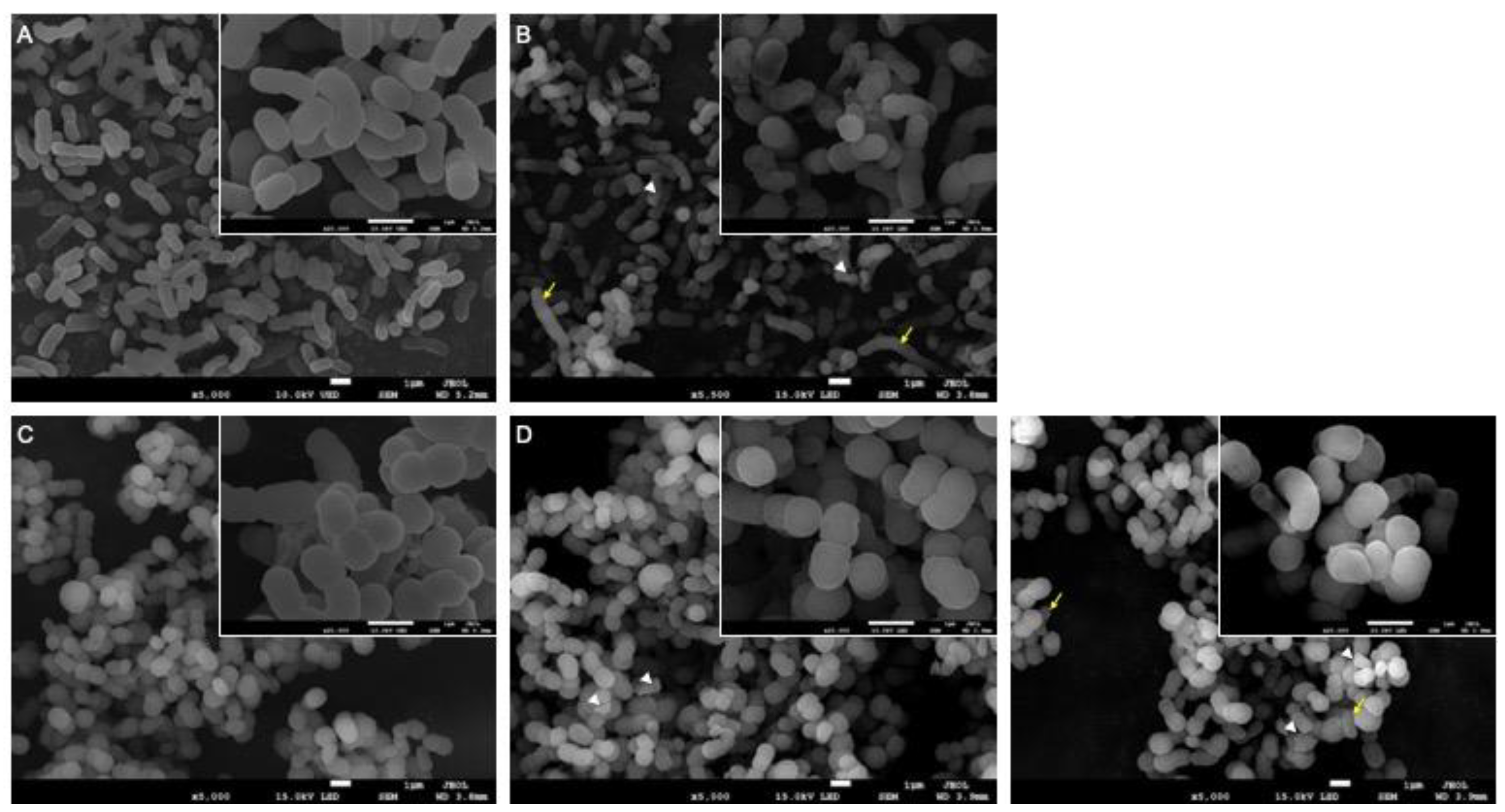
2.8. Scanning Electron Microscopy (SEM)
2.9. Statistical Analysis
3. Results
3.1. The A. baumannii Clinical Isolate and the Spontaneous Mutant
3.2. Genomic Features of the Z0317AB0082 Parental Isolate and the Colistin-Resistant Isogenic Mutant
3.3. Morphological Alterations Due to the PmrB Mutation
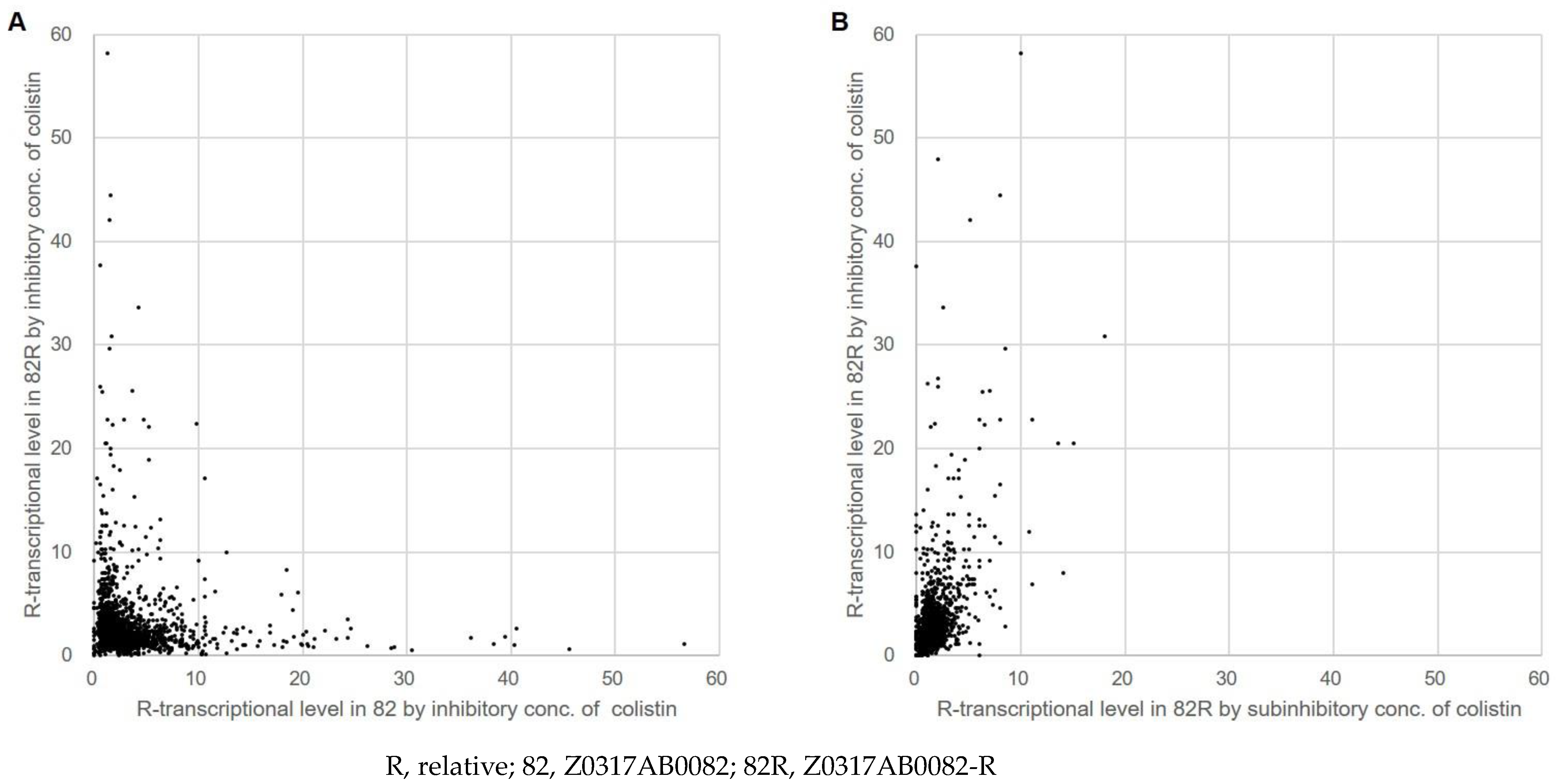
3.4. Effects of Colistin on the Transcriptomic Profiles of the Colistin-Susceptible and -Resistant Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dijkshoorn, L.; Nemec, A.; Seifert, H. An increasing threat in hospitals: Multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 2007, 5, 939–951. [Google Scholar] [CrossRef]
- Landman, D.; Georgescu, C.; Martin, D.A.; Quale, J. Polymyxins revisited. Clin. Microbiol. Rev. 2008, 21, 449–465. [Google Scholar] [CrossRef]
- Li, J.; Nation, R.L.; Turnidge, J.D.; Milne, R.W.; Coulthard, K.; Rayner, C.R.; Paterson, D.L. Colistin: The re-emerging antibiotic for multidrug-resistant Gram-negative bacterial infections. Lancet Infect. Dis. 2006, 6, 589–601. [Google Scholar] [CrossRef]
- Benedict, R.G.; Langlykke, A.F. Antibiotic activity of Bacillus polymyxa. J. Bacteriol. 1947, 54, 24. [Google Scholar]
- Kelesidis, T.; Falagas, M.E. The safety of polymyxin antibiotics. Expert. Opin. Drug Saf. 2015, 14, 1687–1701. [Google Scholar] [CrossRef]
- Moffatt, J.H.; Harper, M.; Harrison, P.; Hale, J.D.; Vinogradov, E.; Seemann, T.; Boyce, J.D. Colistin resistance in Acinetobacter baumannii is mediated by complete loss of lipopolysaccharide production. Antimicrob. Agents Chemother. 2010, 54, 4971–4977. [Google Scholar] [CrossRef]
- Arroyo, L.A.; Herrera, C.M.; Fernandez, L.; Hankins, J.V.; Trent, M.S.; Hancock, R.E. The pmrCAB operon mediates polymyxin resistance in Acinetobacter baumannii ATCC 17978 and clinical isolates through phosphoethanolamine modification of lipid A. Antimicrob Agents Chemother. 2011, 55, 3743–3751. [Google Scholar] [CrossRef]
- Beceiro, A.; Moreno, A.; Fernández, N.; Vallejo, J.A.; Aranda, J.; Adler, B.; Harper, M.; Boyce, J.D.; Bou, G. Biological cost of different mechanisms of colistin resistance and their impact on virulence in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2014, 58, 518–526. [Google Scholar] [CrossRef]
- Yoon, E.J.; Kim, H.S.; Woo, H.; Choi, Y.J.; Won, D.; Choi, J.R.; Kim, Y.A.; Jeong, S.H. Trajectory of genetic alterations associated with colistin resistance in Acinetobacter baumannii during an in-hospital outbreak of infection. J. Antimicrob. Chemother. 2021, 77, 69–73. [Google Scholar] [CrossRef]
- Lin, M.F.; Lin, Y.Y.; Lan, C.Y. Contribution of EmrAB efflux pumps to colistin resistance in Acinetobacter baumannii. J. Microbiol. 2017, 55, 130–136. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Chandler, C.E.; Leung, L.M.; McElheny, C.L.; Mettus, R.T.; Shanks, R.M.; Liu, J.H.; Goodlett, D.R.; Ernst, R.K.; Doi, Y. Structural Modification of Lipopolysaccharide Conferred by mcr-1 in Gram-Negative ESKAPE Pathogens. Antimicrob. Agents Chemother. 2017, 61, e00580-17. [Google Scholar] [CrossRef]
- Jeannot, K.; Bolard, A.; Plesiat, P. Resistance to polymyxins in Gram-negative organisms. Int. J. Antimicrob. Agents. 2017, 49, 526–535. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk Susceptibility Tests, 13th ed.; CLSI Guideline M02-A13; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Document M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Yoon, E.J.; Hong, J.S.; Yang, J.W.; Lee, K.J.; Lee, H.; Jeong, S.H. Detection of mcr-1 Plasmids in Enterobacteriaceae Isolates From Human Specimens: Comparison With Those in Escherichia coli Isolates From Livestock in Korea. Ann. Lab. Med. 2018, 38, 555–562. [Google Scholar] [CrossRef]
- Sherman, E.X.; Wozniak, J.E.; Weiss, D.S. Methods to Evaluate Colistin Heteroresistance in Acinetobacter baumannii. Methods Mol. Biol. 2019, 1946, 39–50. [Google Scholar] [CrossRef]
- Bartual, S.G. Development of a multilocus sequence typing scheme for characterization of clinical isolates of Acinetobacter baumannii. J. Clin. Microbiol. 2005, 43, 4382–4390. [Google Scholar] [CrossRef]
- Lesho, E.; Yoon, E.J.; McGann, P.; Snesrud, E.; Kwak, Y.; Milillo, M.; Waterman, P.E. Emergence of colistin-resistance in extremely drug-resistant Acinetobacter baumannii containing a novel pmrCAB operon during colistin therapy of wound infections. J. Infect. Dis. 2013, 208, 1142–1151. [Google Scholar] [CrossRef]
- Chen, L.; Lin, J.; Lu, H.; Zhang, X.; Wang, C.; Liu, H.; Zhou, T. Deciphering colistin heteroresistance in Acinetobacter baumannii clinical isolates from Wenzhou, China. J. Antibiot. 2020, 73, 463–470. [Google Scholar] [CrossRef]
- Coyne, S.; Guigon, G.; Courvalin, P.; Périchon, B. Screening and quantification of the expression of antibiotic resistance genes in Acinetobacter baumannii with a microarray. Antimicrob. Agents Chemother. 2010, 54, 333–340. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Monnet, D.L. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Héritier, C.; Poirel, L.; Fournier, P.-E.; Claverie, J.-M.; Raoult, D.; Nordmann, P. Characterization of the naturally occurring oxacillinase of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2005, 49, 4174–4179. [Google Scholar] [CrossRef]
- Zhang, G.; Leclercq, S.O.; Tian, J.; Wang, C.; Yahara, K.; Ai, G.; Feng, J. A new subclass of intrinsic aminoglycoside nucleotidyltransferases, ANT(3”)-II, is horizontally transferred among Acinetobacter spp. by homologous recombination. PLoS Genet. 2017, 13, e1006602. [Google Scholar] [CrossRef]
- Cafiso, V.; Stracquadanio, S.; Lo Verde, F.; Gabriele, G.; Mezzatesta, M.L.; Caio, C.; Stefani, S. Colistin Resistant A. baumannii: Genomic and Transcriptomic Traits Acquired Under Colistin Therapy. Front. Microbiol. 2018, 9, 3195. [Google Scholar] [CrossRef]
- Otero, F.; Santiso, R.; Tamayo, M.; Fernández, J.L.; Bou, G.; Lepe, J.A.; Cisneros, J.M. Rapid Detection of Antibiotic Resistance in Gram-Negative Bacteria Through Assessment of Changes in Cellular Morphology. Microb. Drug Resist. 2017, 23, 157–162. [Google Scholar] [CrossRef]
- Park, Y.K.; Lee, J.Y.; Ko, K.S. Transcriptomic analysis of colistin-susceptible and colistin-resistant isolates identifies genes associated with colistin resistance in Acinetobacter baumannii. Clin. Microbiol. Infect. 2015, 21, 765.e1–765.e7. [Google Scholar] [CrossRef]
- Cianciulli Sesso, A.; Lilić, B.; Amman, F.; Wolfinger, M.T.; Sonnleitner, E.; Bläsi, U. Gene Expression Profiling of Pseudomonas aeruginosa upon Exposure to Colistin and Tobramycin. Front. Microbiol. 2021, 12, 626715. [Google Scholar] [CrossRef]
- Sampson, T.R.; Liu, X.; Schroeder, M.R.; Kraft, C.S.; Burd, E.M.; Weiss, D.S. Rapid killing of Acinetobacter baumannii by polymyxins is mediated by a hydroxyl radical death pathway. Antimicrob. Agents Chemother. 2012, 56, 5642–5649. [Google Scholar] [CrossRef]
- Gerson, S.; Betts, J.W.; Lucaßen, K.; Nodari, C.S.; Wille, J.; Josten, M.; Higgins, P.G. Investigation of Novel pmrB and eptA Mutations in Isogenic Acinetobacter baumannii Isolates Associated with Colistin Resistance and Increased Virulence In Vivo. Antimicrob. Agents Chemother. 2019, 63, 01586-18. [Google Scholar] [CrossRef]
- Choi, A.H.; Slamti, L.; Avci, F.Y.; Pier, G.B.; Maira-Litrán, T. The pgaABCD locus of Acinetobacter baumannii encodes the production of poly-beta-1-6-N-acetylglucosamine, which is critical for biofilm formation. J. Bacteriol. 2009, 191, 5953–5963. [Google Scholar] [CrossRef]
- Almutairi, M.M. Synergistic activities of colistin combined with other antimicrobial agents against colistin-resistant Acinetobacter baumannii clinical isolates. PLoS ONE 2022, 17, e0270908. [Google Scholar] [CrossRef]
- Srisakul, S.; Wannigama, D.L.; Higgins, P.G.; Hurst, C.; Abe, S.; Hongsing, P.; Saethang, T.; Luk-In, S.; Liao, T.; Kueakulpattana, N.; et al. Overcoming addition of phosphoethanolamine to lipid A mediated colistin resistance in Acinetobacter baumannii clinical isolates with colistin-sulbactam combination therapy. Sci. Rep. 2022, 12, 11390. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Cho, J.H.; Kim, H.J.; Han, S.H.; Jeong, S.H.; Byun, M.K. Colistin monotherapy versus colistin/rifampicin combination therapy in pneumonia caused by colistin-resistant Acinetobacter baumannii: A randomised controlled trial. J. Glob. Antimicrob. Resist. 2019, 17, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Dickstein, Y.; Lellouche, J.; Ben Dalak Amar, M.; Schwartz, D.; Nutman, A.; Daitch, V.; Yahav, D.; Leibovici, L.; Skiada, A.; Antoniadou, A.; et al. Treatment Outcomes of Colistin- and Carbapenem-resistant Acinetobacter baumannii Infections: An Exploratory Subgroup Analysis of a Randomized Clinical Trial. Clin. Infect. Dis. 2019, 69, 769–776. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agents | Antimicrobial Susceptibility | |
|---|---|---|
| Z0317AB0082 | Z0317AB0082-R | |
| Penicillin + β-lactamase inhibitor | ||
| Ampicillin + sulbactam | R | R |
| Extended-spectrum cephalosporins | ||
| Cefoxitin | R | R |
| Ceftazidime | R | R |
| Cefepime | R | R |
| Carbapenems | ||
| Ertapenem * | R | R |
| Imipenem | R | R |
| Meropenem | R | R |
| Doripenem | R | R |
| Fluoroquinolones | ||
| Ciprofloxacin | R | R |
| Levofloxacin | R | R |
| Aminoglycosides | ||
| Gentamicin | I | I |
| Tobramycin | S | S |
| Amikacin | S | S |
| Tetracyclines | ||
| Tetracycline | R | R |
| Minocycline | S | S |
| Folate pathway inhibitor | ||
| Trimethoprim-sulfamethoxazole | S | S |
| Polymyxins | ||
| Colistin ** | I | R |
| No. | Type | Position | Z0317AB0082 | Z0317AB0082-R |
|---|---|---|---|---|
| 1 | SNP | In a transferase CDS composing an insertion sequence | T | C |
| 2 | SNP | In a transferase CDS composing an insertion sequence | C | T |
| 3 | SNP | In the pmrB gene | C | T |
| 4 | SNP | In a transferase CDS composing an insertion sequence | A | T |
| 5 | SNP | In a transferase CDS composing an insertion sequence | G | A |
| 6 | SNP | In a transferase CDS composing an insertion sequence | T | C |
| 7 | SNP | In a transferase CDS composing an insertion sequence | C | G |
| 8 | SNP | In a transferase CDS composing an insertion sequence | G | A |
| 9 | SNP | In a transferase CDS composing an insertion sequence | A | C |
| 10 | SNP | Intergenic region | G | A |
| 11 | SNP | Intergenic region | T | C |
| 12 | SNP | Intergenic region | G | A |
| 13 | SNP | Intergenic region | A | G |
| 14 | Insertion | Intergenic region | GTG | GTTTG |
| By Colistin Treatment | Z0317AB0082 | Z0317AB0082-R | ||||
|---|---|---|---|---|---|---|
| Inhibitory Concentration | Subinhibitory Concentration | Inhibitory Concentration | ||||
| Stay | 343 | 11.4% | 720 | 23.2% | 311 | 10.0% |
| Downregulation | 219 | 7.3% | 369 | 11.9% | 170 | 5.5% |
| Upregulation | 2435 | 81.2% | 2021 | 65.0% | 2629 | 84.5% |
| Total * | 2997 | 100% | 3110 | 100% | 3110 | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, E.-J.; Mo, J.W.; Kim, J.-w.; Jeong, M.C.; Yoo, J.S. Alteration in the Morphological and Transcriptomic Profiles of Acinetobacter baumannii after Exposure to Colistin. Microorganisms 2024, 12, 1644. https://doi.org/10.3390/microorganisms12081644
Yoon E-J, Mo JW, Kim J-w, Jeong MC, Yoo JS. Alteration in the Morphological and Transcriptomic Profiles of Acinetobacter baumannii after Exposure to Colistin. Microorganisms. 2024; 12(8):1644. https://doi.org/10.3390/microorganisms12081644
Chicago/Turabian StyleYoon, Eun-Jeong, Jun Won Mo, Jee-woong Kim, Min Chul Jeong, and Jung Sik Yoo. 2024. "Alteration in the Morphological and Transcriptomic Profiles of Acinetobacter baumannii after Exposure to Colistin" Microorganisms 12, no. 8: 1644. https://doi.org/10.3390/microorganisms12081644