
Iris setosa Pall. ex Link Extract Reveals Amoebicidal Activity against Acanthamoeba castellanii and Acanthamoeba polyphaga with Low Toxicity to Human Corneal Cells
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of ISE and ISE Subfractions
2.2. Cultivation of Acanthamoeba and Human Corneal Epithelium Cells
2.3. Amoebicidal Assay
2.4. Cytotoxicity Assay for HCE-2 Cells
2.5. Apoptosis/Necrosis Assay
2.6. Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick End-Labeling (TUNEL) Assay
2.7. Intracellular Reactive Oxygen Species (ROS) Assay
2.8. Mitochondrial Membrane Potential (MMP) Assay
2.9. ATP Assay
2.10. Caspase-3 Assay
2.11. Cysticidal Activity Assay
2.12. Statistical Analysis
3. Results
3.1. ISE Showed Amoebicidal Activity against Acanthamoeba Trophozoites
3.2. ISE-BuOH Showed Selective Amoebicidal Activity against Acanthamoeba Trophozoites
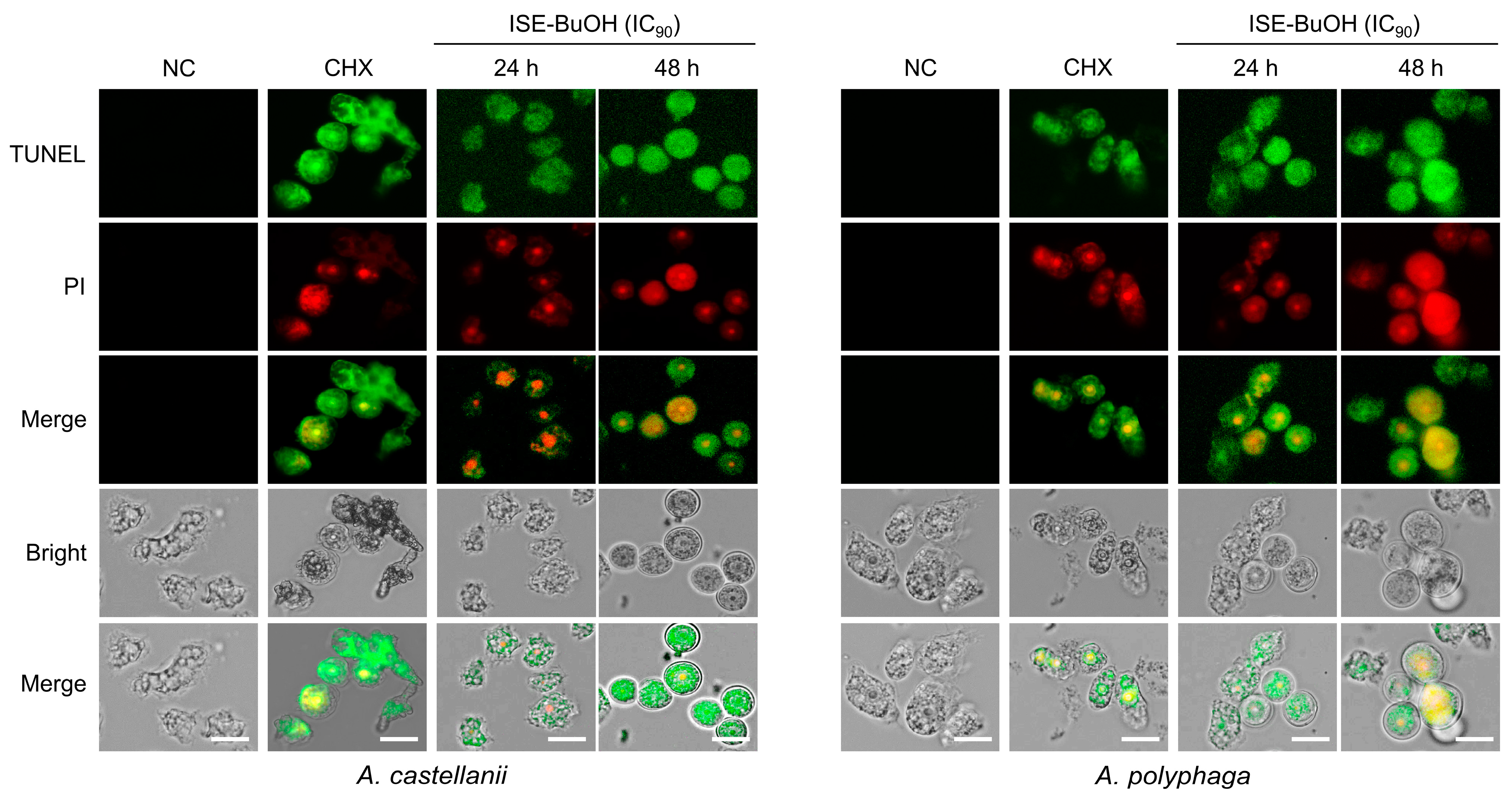
3.3. ISE-BuOH Induced Apoptosis-like Programmed Cell Death (PCD) in A. castellanii and A. polyphaga Trophozoites
3.4. ISE-BuOH Enhanced ROS Production in A. castellanii and A. polyphaga Trophozoites
3.5. ISE-BuOH Compromised Mitochondrial Function in A. castellanii and A. polyphaga Trophozoites
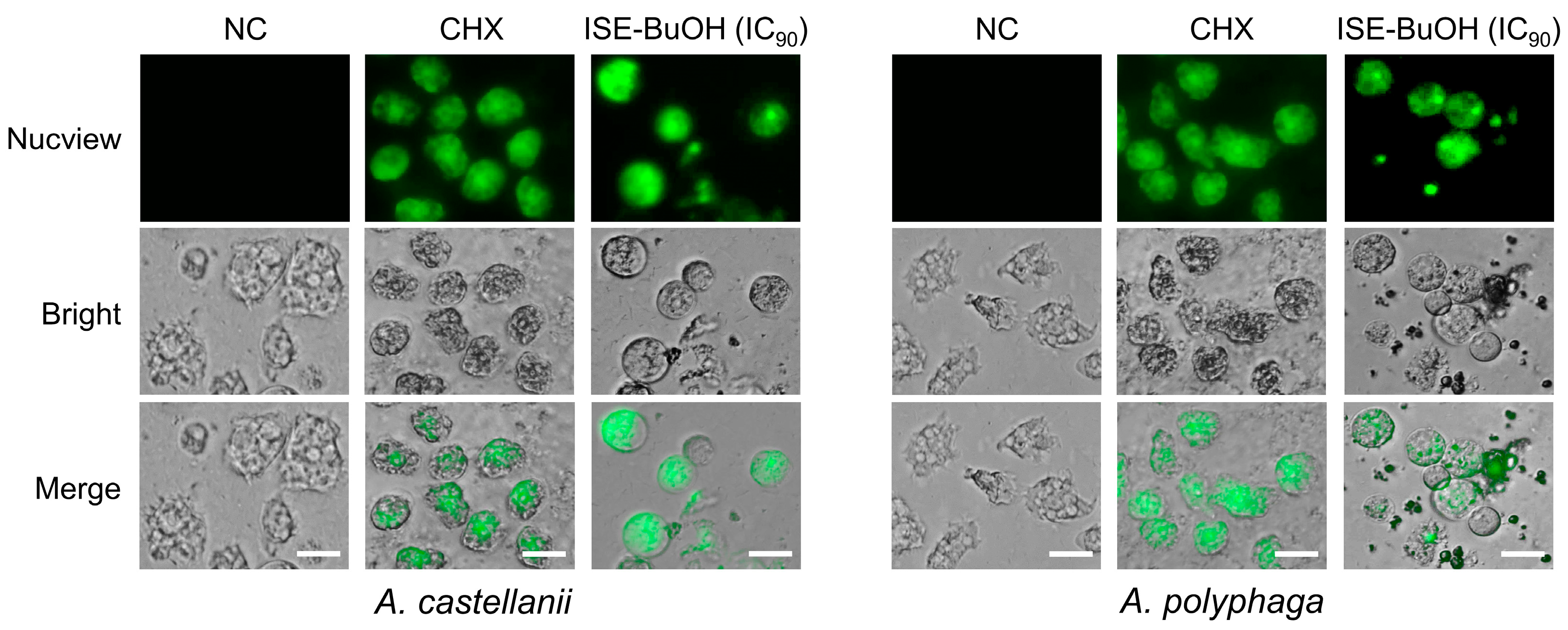
3.6. ISE-BuOH Increased Caspase-3 in A. castellanii and A. polyphaga Trophozoites
3.7. ISE-BuOH Revealed Partial Cysticidal Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Lacerda, A.G.; Lira, M. Acanthamoeba Keratitis: A Review of Biology, Pathophysiology and Epidemiology. Ophthalmic Physiol. Opt. 2021, 41, 116–135. [Google Scholar] [CrossRef] [PubMed]
- Marciano-Cabral, F.; Cabral, G. Acanthamoeba spp. as Agents of Disease in Humans. Clin. Microbiol. Rev. 2003, 16, 273–307. [Google Scholar] [CrossRef] [PubMed]
- Yanai, R.; Yamada, N.; Ueda, K.; Tajiri, M.; Matsumoto, T.; Kido, K.; Nakamura, S.; Saito, F.; Nishida, T. Evaluation of Povidone-Iodine as a Disinfectant Solution for Contact Lenses: Antimicrobial Activity and Cytotoxicity for Corneal Epithelial Cells. Contact Lens Anterior Eye 2006, 29, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Morales, J.; Khan, N.A.; Walochnik, J. An Update on Acanthamoeba Keratitis: Diagnosis, Pathogenesis and Treatment. Parasite 2015, 22, 10. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Oum, B.S.; Choi, H.Y.; Yu, H.S.; Lee, J.S. Cysticidal Effect on Acanthamoeba and Toxicity on Human Keratocytes by Polyhexamethylene Biguanide and Chlorhexidine. Cornea 2007, 26, 736–741. [Google Scholar] [CrossRef]
- Lim, N.; Goh, D.; Bunce, C.; Xing, W.; Fraenkel, G.; Poole, T.R.; Ficker, L. Comparison of Polyhexamethylene Biguanide and Chlorhexidine as Monotherapy Agents in the Treatment of Acanthamoeba Keratitis. Am. J. Ophthalmol. 2008, 145, 130–135. [Google Scholar] [CrossRef]
- Lorenzo-Morales, J.; Martín-Navarro, C.M.; López-Arencibia, A.; Arnalich-Montiel, F.; Piñero, J.E.; Valladares, B. Acanthamoeba Keratitis: An Emerging Disease Gathering Importance Worldwide? Trends Parasitol. 2013, 29, 181–187. [Google Scholar] [CrossRef]
- Boonhok, R.; Sangkanu, S.; Norouzi, R.; Siyadatpanah, A.; Mirzaei, F.; Mitsuwan, W.; Charong, N.; Wisessombat, S.; Pereira, M.D.L.; Rahmatullah, M.; et al. Amoebicidal Activity of Cassia angustifolia Extract and Its Effect on Acanthamoeba triangularis Autophagy-Related Gene Expression at the Transcriptional Level. Parasitology 2021, 148, 1074–1082. [Google Scholar] [CrossRef] [PubMed]
- Sangkanu, S.; Mitsuwan, W.; Mahboob, T.; Mahabusarakam, W.; Chewchanwuttiwong, S.; Siphakdi, P.; Jimoh, T.O.; Wilairatana, P.; Dolma, K.G.; de Lourdes Pereira, M.; et al. Phytochemical, Anti-Acanthamoeba, and Anti-Adhesion Properties of Garcinia mangostana Flower as Preventive Contact Lens Solution. Acta Trop. 2022, 226, 106266. [Google Scholar] [CrossRef]
- Maliński, M.P.; Budzianowski, J.; Kikowska, M.; Derda, M.; Jaworska, M.M.; Mlynarczyk, D.T.; Szukalska, M.; Florek, E.; Thiem, B. Two Ecdysteroids Isolated from Micropropagated Lychnis flos-Cuculi and the Biological Activity of Plant. Molecules 2021, 26, 904. [Google Scholar] [CrossRef]
- Mitsuwan, W.; Sangkanu, S.; Romyasamit, C.; Kaewjai, C.; Jimoh, T.O.; de Lourdes Pereira, M.; Siyadatpanah, A.; Kayesth, S.; Nawaz, M.; Rahmatullah, M.; et al. Curcuma longa Rhizome Extract and Curcumin Reduce the Adhesion of Acanthamoeba triangularis Trophozoites and Cysts in Polystyrene Plastic Surface and Contact Lens. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 218–229. [Google Scholar] [CrossRef]
- Fakae, L.B.; Stevenson, C.W.; Zhu, X.-Q.; Elsheikha, H.M. In Vitro Activity of Camellia sinensis (Green Tea) against Trophozoites and Cysts of Acanthamoeba castellanii. Int. J. Parasitol. Drugs Drug Resist. 2020, 13, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Nayeri, T.; Bineshian, F.; Khoshzaban, F.; Asl, A.D.; Ghaffarifar, F. Evaluation of the Effects of Rumex obtusifolius Seed and Leaf Extracts Against Acanthamoeba: An in Vitro Study. Infect. Disord. Drug Targets 2020, 21, 211–219. [Google Scholar] [CrossRef]
- Sifaoui, I.; Rodríguez-Expósito, R.L.; Reyes-Batlle, M.; Rizo-Liendo, A.; Piñero, J.E.; Bazzocchi, I.L.; Lorenzo-Morales, J.; Jiménez, I.A. Ursolic Acid Derivatives as Potential Agents against Acanthamoeba spp. Pathogens 2019, 8, 130. [Google Scholar] [CrossRef]
- Mahboob, T.; Azlan, A.M.; Shipton, F.N.; Boonroumkaew, P.; Nor Azman, N.S.; Sekaran, S.D.; Ithoi, I.; Tan, T.C.; Samudi, C.; Wiart, C.; et al. Acanthamoebicidal Activity of Periglaucine A and Betulinic Acid from Pericampylus glaucus (Lam.) Merr. in Vitro. Exp. Parasitol. 2017, 183, 160–166. [Google Scholar] [CrossRef]
- Sifaoui, I.; López-Arencibia, A.; Martín-Navarro, C.M.; Reyes-Batlle, M.; Wagner, C.; Chiboub, O.; Mejri, M.; Valladares, B.; Abderrabba, M.; Piñero, J.E.; et al. Programmed Cell Death in Acanthamoeba castellanii Neff Induced by Several Molecules Present in Olive Leaf Extracts. PLoS ONE 2017, 12, e0183795. [Google Scholar] [CrossRef] [PubMed]
- Lê, H.G.; Choi, J.S.; Hwang, B.S.; Jeong, Y.T.; Kang, J.M.; Võ, T.C.; Cho, P.Y.; Lee, Y.K.; Yoo, W.G.; Hong, Y.; et al. Phragmites australis (Cav.) Trin. Ex Steud. Extract Induces Apoptosis-like Programmed Cell Death in Acanthamoeba castellanii Trophozoites. Plants 2022, 11, 3459. [Google Scholar] [CrossRef]
- Lê, H.G.; Kang, J.-M.; Võ, T.C.; Yoo, W.G.; Hong, Y.; Na, B.-K. (–)-Epicatechin Reveals Amoebicidal Activity against Acanthamoeba castellanii by Activating the Programmed Cell Death Pathway. Phytomedicine 2024, 125, 155389. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cui, Y.; Zhao, C. Flavonoids of the Genus Iris (Iridaceae). Mini-Rev. Med. Chem. 2010, 10, 643–661. [Google Scholar] [CrossRef]
- Xie, G.; Qin, X.; Chen, Y.; Wen, R.; Wu, S.; Qin, M. Alkaloids from the Rhizomes of Iris germanica. Chem. Nat. Compd. 2017, 53, 196–198. [Google Scholar] [CrossRef]
- Kukula-Koch, W.; Sieniawska, E.; Widelski, J.; Urjin, O.; Głowniak, P.; Skalicka-Woźniak, K. Major Secondary Metabolites of Iris spp. Phytochem. Rev. 2015, 14, 51–80. [Google Scholar] [CrossRef]
- Wollenweber, E.; Stevens, J.; Klimo, K.; Knauft, J.; Frank, N.; Gerhäuser, C. Cancer Chemopreventive in Vitro Activities of Isoflavones Isolated from Iris germanica. Planta Med. 2003, 69, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Bensari, S.; Ouelbani, R.; Yilmaz, M.A.; Bensouici, C.; Gokalp, E.; Khelifi, D. Phytochemical Profiles of Iris unguicularis Poir. with Antioxidant, Antibacterial, and Anti-Alzheimer Activities. Acta Sci. Nat. 2020, 7, 74–87. [Google Scholar] [CrossRef]
- Benoit-Vical, F.; Imbert, C.; Bonfils, J.-P.; Sauvaire, Y. Antiplasmodial and Antifungal Activities of Iridal, a Plant Triterpenoid. Phytochemistry 2003, 62, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Nazir, N. Immunomodulatory Activity of Isoflavones Isolated from Iris kashmiriana: Effect on T-Lymphocyte Proliferation and Cytokine Production in Balb/c Mice. Biomed. Prev. Nutr. 2013, 3, 151–157. [Google Scholar] [CrossRef]
- Khatib, S.; Faraloni, C.; Bouissane, L. Exploring the Use of Iris Species: Antioxidant Properties, Phytochemistry, Medicinal and Industrial Applications. Antioxidants 2022, 11, 526. [Google Scholar] [CrossRef] [PubMed]
- Bowers, B.; Korn, E.D. The Fine Structure of Acanthamoeba castellanii (Neff Strain). II. Encystment. J. Cell Biol. 1969, 41, 786–805. [Google Scholar] [CrossRef] [PubMed]
- Baig, A.M.; Iqbal, J.; Khan, N.A. In Vitro Efficacies of Clinically Available Drugs against Growth and Viability of an Acanthamoeba castellanii Keratitis Isolate Belonging to the T4 Genotype. Antimicrob. Agents Chemother. 2013, 57, 3561–3567. [Google Scholar] [CrossRef] [PubMed]
- Kaczanowski, S.; Sajid, M.; Reece, S.E. Evolution of Apoptosis-like Programmed Cell Death in Unicellular Protozoan Parasites. Parasit. Vectors 2011, 4, 44. [Google Scholar] [CrossRef]
- Deponte, M. Programmed Cell Death in Protists. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 1396–1405. [Google Scholar] [CrossRef]
- Reece, S.E.; Pollitt, L.C.; Colegrave, N.; Gardner, A. The Meaning of Death: Evolution and Ecology of Apoptosis in Protozoan Parasites. PLoS Pathog. 2011, 7, e1002320. [Google Scholar] [CrossRef]
- Arnoult, D.; Akarid, K.; Grodet, A.; Petit, P.X.; Estaquier, J.; Ameisen, J.C. On the Evolution of Programmed Cell Death: Apoptosis of the Unicellular Eukaryote Leishmania major Involves Cysteine Proteinase Activation and Mitochondrion Permeabilization. Cell Death Differ. 2002, 9, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Bertholet, S.; Debrabant, A.; Muller, J.; Duncan, R.; Nakhasi, H.L. Programmed Cell Death in the Unicellular Protozoan Parasite Leishmania. Cell Death Differ. 2002, 9, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Lakhani, S.A.; Masud, A.; Kuida, K.; Porter, G.A.; Booth, C.J.; Mehal, W.Z.; Inayat, I.; Flavell, R.A. Caspases 3 and 7: Key Mediators of Mitochondrial Events of Apoptosis. Science 2006, 311, 847–851. [Google Scholar] [CrossRef]
- Lê, H.G.; Kang, J.M.; Võ, T.C.; Na, B.K. Kaempferol Induces Programmed Cell Death in Naegleria fowleri. Phytomedicine 2023, 119, 154994. [Google Scholar] [CrossRef] [PubMed]
- Yousefsani, B.S.; Boozari, M.; Shirani, K.; Jamshidi, A.; Dadmehr, M. A Review on Phytochemical and Therapeutic Potential of Iris germanica. J. Pharm. Pharmacol. 2021, 73, 611–625. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Houghton, P.J.; Hylands, P.J. Cytotoxic Effects of Compounds from Iris tectorum on Human Cancer Cell Lines. J. Ethnopharmacol. 2008, 118, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Kostić, A.Ž.; Gašić, U.M.; Pešić, M.B.; Stanojević, S.P.; Barać, M.B.; Mačukanović-Jocić, M.P.; Avramov, S.N.; Tešić, Ž.L. Phytochemical Analysis and Total Antioxidant Capacity of Rhizome, Above-Ground Vegetative Parts and Flower of Three Iris Species. Chem. Biodivers. 2019, 16, e1800565. [Google Scholar] [CrossRef]
- Sayyed, K.; Hdayed, I.; Tabcheh, M.; Abdel-Razzak, Z.; El-Bitar, H. Antioxidant Properties of the Lebanese Plant Iris x Germanica L. Crude Extracts and Antagonism of Chlorpromazine Toxicity on Saccharomyces Cerevisiae. Drug Chem. Toxicol. 2022, 45, 1168–1179. [Google Scholar] [CrossRef]
- Xie, G.-Y.; Qin, X.-Y.; Liu, R.; Wang, Q.; Lin, B.-B.; Wang, G.-K.; Xu, G.-K.; Wen, R.; Qin, M.-J. New Isoflavones with Cytotoxic Activity from the Rhizomes of Iris germanica L. Nat. Prod. Res. 2013, 27, 2173–2177. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.-H.; Tsai, C.-W.; Tsai, R.-T.; Liu, S.-P.; Chan, T.-M.; Ho, Y.-C.; Lin, H.-L.; Chen, Y.-M.; Hung, H.-S.; Chiu, S.-C.; et al. Irisflorentin Modifies Properties of Mouse Bone Marrow-Derived Dendritic Cells and Reduces the Allergic Contact Hypersensitivity Responses. Cell Transpl. 2015, 24, 573–588. [Google Scholar] [CrossRef]
- Abdel-Baki, P.M.; El-Sherei, M.M.; Khaleel, A.E.; Abdel-Sattar, E.; Salem, M.A.; Okba, M.M. Correlation between Secondary Metabolites of Iris confusa Sealy and Iris pseudacorus L. and Their Newly Explored Antiprotozoal Potentials. BMC Complement. Med. Ther. 2023, 23, 465. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.-R.; Wang, X.; Liu, Y.-F.; Zhang, C.-L.; Ni, G.; Chen, R.-Y.; Yu, D.-Q. Bioactive Flavonoid Glycosides from Whole Plants of Iris japonica. Phytochem. Lett. 2017, 19, 141–144. [Google Scholar] [CrossRef]
- Boonhok, R.; Sangkanu, S.; Chuprom, J.; Srisuphanunt, M.; Norouzi, R.; Siyadatpanah, A.; Mirzaei, F.; Mitsuwan, W.; Wisessombat, S.; de Lourdes Pereira, M.; et al. Peganum harmala Extract Has Antiamoebic Activity to Acanthamoeba triangularis Trophozoites and Changes Expression of Autophagy-Related Genes. Pathogens 2021, 10, 842. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.S.; Kim, S.; Kim, D.S.; Yu, H.S.; Lee, J.E. The amoebicidal effect of Torreya nucifera extract on Acanthamoeba lugdunensis. PLoS ONE 2023, 18, e0281141. [Google Scholar] [CrossRef] [PubMed]
- Mitsuwan, W.; Bunsuwansakul, C.; Leonard, T.E.; Laohaprapanon, S.; Hounkong, K.; Bunluepuech, K.; Kaewjai, C.; Mahboob, T.; Sumudi Raju, C.; Dhobi, M.; et al. Curcuma longa ethanol extract and Curcumin inhibit the growth of Acanthamoeba triangularis trophozoites and cysts isolated from water reservoirs at Walailak University, Thailand. Pathog. Glob. Health 2020, 114, 194–204. [Google Scholar] [CrossRef]
- Fakae, L.B.; Harun, M.S.R.; Ting, D.S.J.; Dua, H.S.; Cave, G.W.V.; Zhu, X.Q.; Stevenson, C.W.; Elsheikha, H.M. Camellia sinensis solvent extract, epigallocatechin gallate and caffeine confer trophocidal and cysticidal effects against Acanthamoeba castellanii. Acta Trop. 2023, 237, 106729. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, D. Encystment in Acanthamoeba castellanii: A Review. Exp. Parasitol. 2014, 145, S20–S27. [Google Scholar] [CrossRef] [PubMed]
- Dudley, R.; Jarroll, E.L.; Khan, N.A. Carbohydrate Analysis of Acanthamoeba castellanii. Exp. Parasitol. 2009, 122, 338–343. [Google Scholar] [CrossRef]
- Johnston, S.P.; Sriram, R.; Qvarnstrom, Y.; Roy, S.; Verani, J.; Yoder, J.; Lorick, S.; Roberts, J.; Beach, M.J.; Visvesvara, G. Resistance of Acanthamoeba Cysts to Disinfection in Multiple Contact Lens Solutions. J. Clin. Microbiol. 2009, 47, 2040–2045. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lê, H.G.; Hwang, B.S.; Choi, J.-S.; Jeong, Y.T.; Kang, J.-M.; Võ, T.C.; Oh, Y.T.; Na, B.-K. Iris setosa Pall. ex Link Extract Reveals Amoebicidal Activity against Acanthamoeba castellanii and Acanthamoeba polyphaga with Low Toxicity to Human Corneal Cells. Microorganisms 2024, 12, 1658. https://doi.org/10.3390/microorganisms12081658
Lê HG, Hwang BS, Choi J-S, Jeong YT, Kang J-M, Võ TC, Oh YT, Na B-K. Iris setosa Pall. ex Link Extract Reveals Amoebicidal Activity against Acanthamoeba castellanii and Acanthamoeba polyphaga with Low Toxicity to Human Corneal Cells. Microorganisms. 2024; 12(8):1658. https://doi.org/10.3390/microorganisms12081658
Chicago/Turabian StyleLê, Hương Giang, Buyng Su Hwang, Ji-Su Choi, Yong Tae Jeong, Jung-Mi Kang, Tuấn Cường Võ, Young Taek Oh, and Byoung-Kuk Na. 2024. "Iris setosa Pall. ex Link Extract Reveals Amoebicidal Activity against Acanthamoeba castellanii and Acanthamoeba polyphaga with Low Toxicity to Human Corneal Cells" Microorganisms 12, no. 8: 1658. https://doi.org/10.3390/microorganisms12081658