
Gut Microbes Associated with Neurodegenerative Disorders: A Comprehensive Review of the Literature

 , ,
, ,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Selection Criteria
2.3. Data Extraction
2.4. Data Analysis
3. Results
3.1. Type of Study
3.2. Related Disease
3.3. Methods of Evaluation
3.4. Estimation of Microbiome
3.5. Main Findings
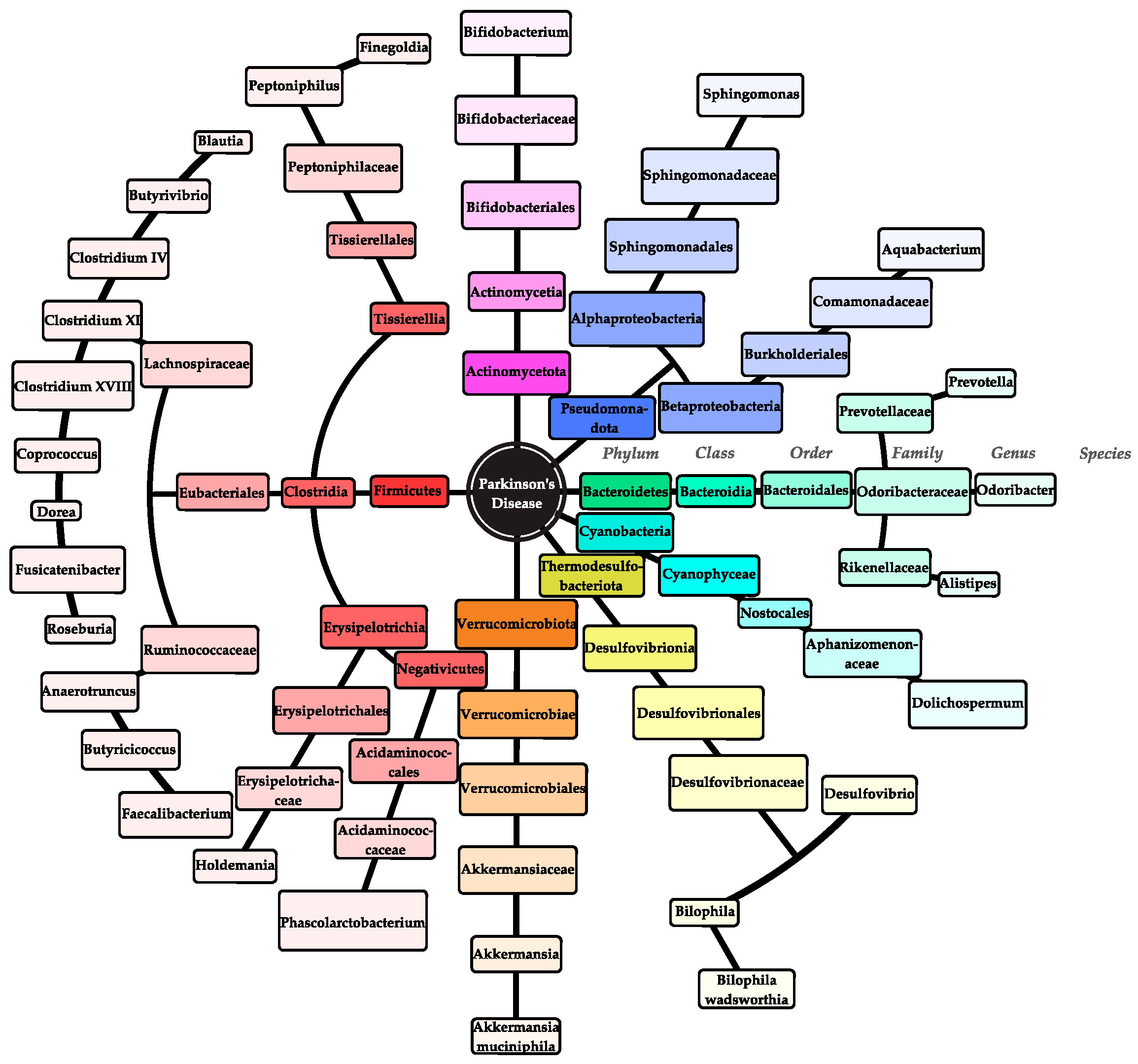
3.5.1. Parkinson’s Disease
3.5.2. Alzheimer’s Disease
3.5.3. Amyotrophic Lateral Sclerosis
3.5.4. Multiple System Atrophy
3.5.5. Creutzfeldt–Jakob Disease
3.5.6. Huntington’s Disease
3.5.7. Multiple Sclerosis
4. Discussion
5. Limitations and Future Directions
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, H.; Yang, F.; Zhang, S.; Xin, R.; Sun, Y. Genetic and environmental factors in Alzheimer’s and Parkinson’s diseases and promising therapeutic intervention via fecal microbiota transplantation. NPJ Park. Dis. 2021, 7, 70. [Google Scholar] [CrossRef]
- National Institute of Environmental Health Sciences. Neurodegenerative Diseases. Available online: https://www.niehs.nih.gov/research/supported/health/neurodegenerative (accessed on 24 July 2024).
- Ghezzi, L.; Cantoni, C.; Rotondo, E.; Galimberti, D. The Gut Microbiome-Brain Crosstalk in Neurodegenerative Diseases. Biomedicines 2022, 10, 1486. [Google Scholar] [CrossRef]
- Mejía-Granados, D.M.; Villasana-Salazar, B.; Coan, A.C.; Rizzi, L.; Balthazar, M.L.F.; Godoi, A.B.; Canto, A.M.D.; Rosa, D.C.D.; Silva, L.S.; Tacla, R.D.R.; et al. Gut microbiome in neuropsychiatric disorders. Arq. Neuropsiquiatr. 2022, 80, 192–207. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, S.D.; Elarova, I.; Ruben, M.; Tan, F.; Counts, S.E.; Eberwine, J.H.; Trojanowski, J.Q.; Hemby, S.E.; Mufson, E.J.; Che, S. Single-cell gene expression analysis: Implications for neurodegenerative and neuropsychiatric disorders. Neurochem. Res. 2004, 29, 1053–1064. [Google Scholar] [CrossRef]
- Gupta, R.; Advani, D.; Yadav, D.; Ambasta, R.K.; Kumar, P. Dissecting the Relationship Between Neuropsychiatric and Neurodegenerative Disorders. Mol. Neurobiol. 2023, 60, 6476–6529. [Google Scholar] [CrossRef]
- Wen, H.; Jung, H.; Li, X. Drug Delivery Approaches in Addressing Clinical Pharmacology-Related Issues: Opportunities and Challenges. Aaps. J. 2015, 17, 1327–1340. [Google Scholar] [CrossRef] [PubMed]
- Cuttler, K.; Hassan, M.; Carr, J.; Cloete, R.; Bardien, S. Emerging evidence implicating a role for neurexins in neurodegenerative and neuropsychiatric disorders. Open Biol. 2021, 11, 210091. [Google Scholar] [CrossRef]
- Tsamakis, K.; Galinaki, S.; Alevyzakis, E.; Hortis, I.; Tsiptsios, D.; Kollintza, E.; Kympouropoulos, S.; Triantafyllou, K.; Smyrnis, N.; Rizos, E. Gut Microbiome: A Brief Review on Its Role in Schizophrenia and First Episode of Psychosis. Microorganisms 2022, 10, 1121. [Google Scholar] [CrossRef]
- Merkouris, E.; Mavroudi, T.; Miliotas, D.; Tsiptsios, D.; Serdari, A.; Christidi, F.; Doskas, T.K.; Mueller, C.; Tsamakis, K. Probiotics’ Effects in the Treatment of Anxiety and Depression: A Comprehensive Review of 2014–2023 Clinical Trials. Microorganisms 2024, 12, 411. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Badal, V.D.; Vaccariello, E.D.; Murray, E.R.; Yu, K.E.; Knight, R.; Jeste, D.V.; Nguyen, T.T. The Gut Microbiome, Aging, and Longevity: A Systematic Review. Nutrients 2020, 12, 3759. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P. Influence of Foods and Nutrition on the Gut Microbiome and Implications for Intestinal Health. Int. J. Mol. Sci. 2022, 23, 9588. [Google Scholar] [CrossRef]
- Wilson, A.S.; Koller, K.R.; Ramaboli, M.C.; Nesengani, L.T.; Ocvirk, S.; Chen, C.; Flanagan, C.A.; Sapp, F.R.; Merritt, Z.T.; Bhatti, F.; et al. Diet and the Human Gut Microbiome: An International Review. Dig. Dis. Sci. 2020, 65, 723–740. [Google Scholar] [CrossRef] [PubMed]
- Miwa, H.; Kondo, T. Alteration of eating behaviors in patients with Parkinson’s disease: Possibly overlooked? Neurocase 2008, 14, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zhan, Z.; Zhang, L.; Zhu, J.; Liu, Y.; Zhang, L.; Ge, J.; Zhao, Y.; Zhang, L.; Dong, J. Prevalence and Risk Factors for Malnutrition in Patients With Parkinson’s Disease. Front. Neurol. 2020, 11, 533731. [Google Scholar] [CrossRef]
- Harach, T.; Marungruang, N.; Duthilleul, N.; Cheatham, V.; Mc Coy, K.D.; Frisoni, G.; Neher, J.J.; Fåk, F.; Jucker, M.; Lasser, T.; et al. Reduction of Abeta amyloid pathology in APPPS1 transgenic mice in the absence of gut microbiota. Sci. Rep. 2017, 7, 41802. [Google Scholar] [CrossRef]
- Vogt, N.M.; Kerby, R.L.; Dill-McFarland, K.A.; Harding, S.J.; Merluzzi, A.P.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Zetterberg, H.; Blennow, K.; et al. Gut microbiome alterations in Alzheimer’s disease. Sci. Rep. 2017, 7, 13537. [Google Scholar] [CrossRef]
- Liu, P.; Wu, L.; Peng, G.; Han, Y.; Tang, R.; Ge, J.; Zhang, L.; Jia, L.; Yue, S.; Zhou, K.; et al. Altered microbiomes distinguish Alzheimer’s disease from amnestic mild cognitive impairment and health in a Chinese cohort. Brain Behav. Immun. 2019, 80, 633–643. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.; Jansen Steur, E.N.; Braak, E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Hor, J.W.; Lim, S.Y.; Khor, E.S.; Chong, K.K.; Song, S.L.; Ibrahim, N.M.; Teh, C.S.J.; Chong, C.W.; Hilmi, I.N.; Tan, A.H. Fecal Calprotectin in Parkinson’s Disease and Multiple System Atrophy. J. Mov. Disord. 2022, 15, 106–114. [Google Scholar] [CrossRef]
- Morais, L.H.; Hara, D.B.; Bicca, M.A.; Poli, A.; Takahashi, R.N. Early signs of colonic inflammation, intestinal dysfunction, and olfactory impairments in the rotenone-induced mouse model of Parkinson’s disease. Behav. Pharmacol. 2018, 29, 199–210. [Google Scholar] [CrossRef]
- Pellegrini, C.; Fornai, M.; Colucci, R.; Tirotta, E.; Blandini, F.; Levandis, G.; Cerri, S.; Segnani, C.; Ippolito, C.; Bernardini, N.; et al. Alteration of colonic excitatory tachykininergic motility and enteric inflammation following dopaminergic nigrostriatal neurodegeneration. J. Neuroinflamm. 2016, 13, 146. [Google Scholar] [CrossRef]
- Wu, S.; Yi, J.; Zhang, Y.G.; Zhou, J.; Sun, J. Leaky intestine and impaired microbiome in an amyotrophic lateral sclerosis mouse model. Physiol. Rep. 2015, 3, e12356. [Google Scholar] [CrossRef]
- Zhang, Y.; Ogbu, D.; Garrett, S.; Xia, Y.; Sun, J. Aberrant enteric neuromuscular system and dysbiosis in amyotrophic lateral sclerosis. Gut Microbes 2021, 13, 1996848. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Shen, J.; Chen, K.; Zhou, J.; Liao, Q.; Lu, K.; Yuan, J.; Bi, F. The alteration of gut microbiome and metabolism in amyotrophic lateral sclerosis patients. Sci. Rep. 2020, 10, 12998. [Google Scholar] [CrossRef]
- Nicholson, K.; Bjornevik, K.; Abu-Ali, G.; Chan, J.; Cortese, M.; Dedi, B.; Jeon, M.; Xavier, R.; Huttenhower, C.; Ascherio, A.; et al. The human gut microbiota in people with amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Frontotemporal Degener. 2021, 22, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Wasser, C.I.; Mercieca, E.C.; Kong, G.; Hannan, A.J.; McKeown, S.J.; Glikmann-Johnston, Y.; Stout, J.C. Gut dysbiosis in Huntington’s disease: Associations among gut microbiota, cognitive performance and clinical outcomes. Brain Commun. 2020, 2, fcaa110. [Google Scholar] [CrossRef]
- Qian, Y.; Yang, X.; Xu, S.; Wu, C.; Song, Y.; Qin, N.; Chen, S.D.; Xiao, Q. Alteration of the fecal microbiota in Chinese patients with Parkinson’s disease. Brain Behav. Immun. 2018, 70, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lu, G.; Luo, E.; Wu, B.; Li, Z.; Guo, J.; Xia, Z.; Zheng, C.; Su, Q.; Zeng, Y.; et al. Oral, Nasal, and Gut Microbiota in Parkinson’s Disease. Neuroscience 2022, 480, 65–78. [Google Scholar] [CrossRef]
- Weis, S.; Meisner, A.; Schwiertz, A.; Unger, M.M.; Becker, A.; Faßbender, K.; Schnell, S.; Schäfer, K.H.; Egert, M. Association between Parkinson’s disease and the faecal eukaryotic microbiota. NPJ Park. Dis. 2021, 7, 101. [Google Scholar] [CrossRef]
- Raghavan, K.; Dedeepiya, V.D.; Yamamoto, N.; Ikewaki, N.; Sonoda, T.; Iwasaki, M.; Kandaswamy, R.S.; Senthilkumar, R.; Preethy, S.; Abraham, S.J.K. Benefits of Gut Microbiota Reconstitution by Beta 1,3-1,6 Glucans in Subjects with Autism Spectrum Disorder and other Neurodegenerative Diseases. J. Alzheimers Dis. 2023, 94, S241–S252. [Google Scholar] [CrossRef] [PubMed]
- Murros, K.E.; Huynh, V.A.; Takala, T.M.; Saris, P.E.J. Desulfovibrio Bacteria Are Associated With Parkinson’s Disease. Front. Cell Infect. Microbiol. 2021, 11, 652617. [Google Scholar] [CrossRef]
- Weis, S.; Schwiertz, A.; Unger, M.M.; Becker, A.; Faßbender, K.; Ratering, S.; Kohl, M.; Schnell, S.; Schäfer, K.H.; Egert, M. Effect of Parkinson’s disease and related medications on the composition of the fecal bacterial microbiota. NPJ Parkinsons Dis. 2019, 5, 28. [Google Scholar] [CrossRef]
- Vascellari, S.; Palmas, V.; Melis, M.; Pisanu, S.; Cusano, R.; Uva, P.; Perra, D.; Madau, V.; Sarchioto, M.; Oppo, V.; et al. Gut Microbiota and Metabolome Alterations Associated with Parkinson’s Disease. mSystems 2020, 5, e00561-20. [Google Scholar] [CrossRef]
- Hertel, J.; Harms, A.C.; Heinken, A.; Baldini, F.; Thinnes, C.C.; Glaab, E.; Vasco, D.A.; Pietzner, M.; Stewart, I.D.; Wareham, N.J.; et al. Integrated Analyses of Microbiome and Longitudinal Metabolome Data Reveal Microbial-Host Interactions on Sulfur Metabolism in Parkinson’s Disease. Cell Rep. 2019, 29, 1767–1777.e8. [Google Scholar] [CrossRef] [PubMed]
- Hegelmaier, T.; Lebbing, M.; Duscha, A.; Tomaske, L.; Tönges, L.; Holm, J.B.; Bjørn Nielsen, H.; Gatermann, S.G.; Przuntek, H.; Haghikia, A. Interventional Influence of the Intestinal Microbiome Through Dietary Intervention and Bowel Cleansing Might Improve Motor Symptoms in Parkinson’s Disease. Cells 2020, 9, 376. [Google Scholar] [CrossRef]
- Zhuang, Z.; Yang, R.; Wang, W.; Qi, L.; Huang, T. Associations between gut microbiota and Alzheimer’s disease, major depressive disorder, and schizophrenia. J. Neuroinflamm. 2020, 17, 288. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, Y.; Quan, M.; Zhao, H.; Jia, J. Gut Microbiota Changes and Their Correlation with Cognitive and Neuropsychiatric Symptoms in Alzheimer’s Disease. J. Alzheimers Dis. 2021, 81, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Brenner, D.; Hiergeist, A.; Adis, C.; Mayer, B.; Gessner, A.; Ludolph, A.C.; Weishaupt, J.H. The fecal microbiome of ALS patients. Neurobiol. Aging 2018, 61, 132–137. [Google Scholar] [CrossRef]
- Wan, L.; Zhou, X.; Wang, C.; Chen, Z.; Peng, H.; Hou, X.; Peng, Y.; Wang, P.; Li, T.; Yuan, H.; et al. Alterations of the Gut Microbiota in Multiple System Atrophy Patients. Front. Neurosci. 2019, 13, 1102. [Google Scholar] [CrossRef]
- Barone, M.; Mendozzi, L.; D’Amico, F.; Saresella, M.; Rampelli, S.; Piancone, F.; La Rosa, F.; Marventano, I.; Clerici, M.; d’Arma, A.; et al. Influence of a High-Impact Multidimensional Rehabilitation Program on the Gut Microbiota of Patients with Multiple Sclerosis. Int. J. Mol. Sci. 2021, 22, 7173. [Google Scholar] [CrossRef]
- Guo, Y.; Xu, Y.; Lin, X.; Zhen, Z.; Yi, F.; Guan, H.; Shi, Q.; Sun, W.; Yang, A.; Dong, X.; et al. Creutzfeldt-Jakob Disease: Alterations of Gut Microbiota. Front. Neurol. 2022, 13, 832599. [Google Scholar] [CrossRef]
- Su, X. Elucidating the Beta-Diversity of the Microbiome: From Global Alignment to Local Alignment. mSystems 2021, 6, e0036321. [Google Scholar] [CrossRef]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480.e12. [Google Scholar] [CrossRef]
- Hill-Burns, E.M.; Debelius, J.W.; Morton, J.T.; Wissemann, W.T.; Lewis, M.R.; Wallen, Z.D.; Peddada, S.D.; Factor, S.A.; Molho, E.; Zabetian, C.P.; et al. Parkinson’s disease and Parkinson’s disease medications have distinct signatures of the gut microbiome. Mov. Disord. 2017, 32, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Fanciulli, A.; Wenning, G.K. Multiple-system atrophy. N. Engl. J. Med. 2015, 372, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, C.; Chen, Z.; Peng, Y.; Peng, H.; Hou, X.; Ye, W.; Qiu, R.; Xia, K.; Tang, B.; et al. Association of TNF-α rs1799964 and IL-1β rs16944 polymorphisms with multiple system atrophy in Chinese Han population. Int. J. Neurosci. 2018, 128, 761–764. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. The Microbiome-Gut-Brain Axis in Health and Disease. Gastroenterol. Clin. N. Am. 2017, 46, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Kaupmann, K. Don’t worry ‘B’ happy!: A role for GABA(B) receptors in anxiety and depression. Trends Pharmacol. Sci. 2005, 26, 36–43. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Jiang, C.; Li, G.; Huang, P.; Liu, Z.; Zhao, B. The Gut Microbiota and Alzheimer’s Disease. J. Alzheimers Dis. 2017, 58, 1–15. [Google Scholar] [CrossRef]
- Cani, P.D. Human gut microbiome: Hopes, threats and promises. Gut 2018, 67, 1716–1725. [Google Scholar] [CrossRef]
- Ordoñez-Rodriguez, A.; Roman, P.; Rueda-Ruzafa, L.; Campos-Rios, A.; Cardona, D. Changes in Gut Microbiota and Multiple Sclerosis: A Systematic Review. Int. J. Environ. Res. Public Health 2023, 20, 4624. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.M.M. Microbiota-Brain-Gut Axis and Neurodegenerative Diseases. Curr. Neurol. Neurosci. Rep. 2017, 17, 94. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell Infect. Microbiol. 2021, 11, 625913. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Type of Study | n |
|---|---|
| Cohort study | 13 |
| Case–control study | 4 |
| Randomized pilot clinical study | 1 |
| Two-sample bi-directional Mendelian randomization analysis | 1 |
| Disease | n | Authors |
|---|---|---|
| Parkinson’s disease | 9 | Qian et al. (2018) [29], Li et al. (2022) [30], Weis et al. (2021) [31], Raghavan et al. (2023) * [32], Murros et al. (2021) [33], Weis et al. (2019) [34], Vascellari et al. (2020) [35], Hertel et al. (2019) [36], Hegelmaier et al. (2020) [37] |
| Alzheimer’s disease * | 4 | Liu et al. (2019) [19], Zhuang et al. (2020) ** [38], Raghavan et al. (2023) * [32], Zhou et al. (2021) [39] |
| Amyotrophic lateral sclerosis | 3 | Zeng et al. (2020) [26], Brenner et al. (2017) [40], Nicholson et al. (2020) [27] |
| Multiple system atrophy | 1 | Wan et al. (2019) [41] |
| Multiple sclerosis | 1 | Barone et al. (2021) [42] |
| Huntington’s disease | 1 | Wasser et al. (2020) [28] |
| Creutzfeldt–Jakob disease | 1 | Guo et al. (2022) [43] |
| Evaluation | n | Disease | Evaluation | n | Disease |
|---|---|---|---|---|---|
| MMSE | 7 | PD, AD, CJD, MSA | Laboratory tests | 1 | AD |
| MoCA | 5 | PD, AD, CJD | Hopkins Verbal Learning Test-Revised | 1 | HD |
| Hoehn and Yahr Scale | 4 | PD | HD-CAB | 1 | HD |
| UPDRS | 3 | PD | CAP | 1 | HD |
| El Escorial Criteria | 3 | ALS | Mass spectrometry | 1 | PD |
| NMS-Quest | 2 | PD | MDS-UPDRS | 1 | PD |
| HAMD | 2 | PD | MFIS-5 | 1 | MS |
| HAMA | 2 | PD | 6MWT | 1 | MS |
| UHDRS | 1 | HD | FFQ | 1 | MS |
| UMSARS | 1 | MSA | GC-MS analysis | 1 | PD |
| Trail Making Test | 1 | HD | Emotion Recognition Task | 1 | HD |
| Symbol Digit Modalities Test | 1 | HD | MRI | 1 | AD |
| Paraclinical tests | 1 | MS | CSF evaluation | 1 | CJD |
| Paced tapping | 1 | HD | CDR-SB | 1 | CJD |
| OTS of Cambridge | 1 | HD | CDR | 1 | AD |
| NPI | 1 | AD | |||
| a/a | Articles/Authors | Type of Study | Related Disease | Method of Evaluation | Estimation of Microbiome-Sequencing Approach | Main Findings |
|---|---|---|---|---|---|---|
| 1 | Qian et al. (2018) [29] | Case–control study | Parkinson’s disease | Hoehn and Yahr Scale, UPDRS total, Part III scores, NMS-Quest, HAMA, HAMD, MMSE, MoCA | Amplification of the V3-V4 region of the bacterial 16S rRNA gene | The relative abundance of the genera Clostridium IV, Aquabacterium, Holdemania, Sphingomonas, Clostridium XVIII, Butyricicoccus, and Anaerotruncus increased in the feces of PD patients. The genera Escherichia/Shigella were negatively associated with disease duration. Aquabacterium, Peptococcus, and Sphingomonas in feces were associated with motor complications. The genera Butyricicoccus and Clostridium XlVb were associated with cognitive impairment. |
| 2 | Wan et al. (2019) [41] | Cohort study | Multiple system atrophy | UMSARS, MMSE | Metagenomic sequencing libraries | The gut microbiota of MSA patients were characterized by increased proportions of the genus Akkermansia and the species Roseburia hominis, Akkermansia muciniphila, Alistipes onderdonkii, Streptococcus parasanguinis, and Staphylococcus xylosus, and decreased proportions of the genera Megamonas, Bifidobacterium, Blautia, and Aggregatibacter and the species Bacteroides coprocola, Megamonas funiformis, Bifidobacterium pseudocatenulatum, Clostridium nexile, Bacteroides plebeius, and Granulicatella adiacens. |
| 3 | Liu et al. (2019) [19] | Cohort study | Alzheimer’s disease | MMSE, MoCA, MRI, laboratory tests | Amplification of the V3-V4 region of the bacterial 16S rRNA gene | The proportion of phylum Firmicutes significantly reduced (decreased abundance of the families Clostridiaceae, Lachnospiraceae, and Ruminococcaceae and the genera Blautia and Ryminococcus), whereas Proteobacteria was highly enriched in AD patients. Gammaproteobacteria, Enterobacteriales, and Enterobacteriaceae showed a progressive enriched prevalence from healthy controls to AD patients. The family Enterobacteriaceae was positively associated with the severity of AD. The relative abundance of Bacteroidetes was significantly enriched in the pre-onset stage of AD and unexpectedly decreased in the AD group to the control group. |
| 4 | Li et al. (2022) [30] | Case–control study | Parkinson’s disease | Hoehn and Yahr Scale, NMS, HAMD, HAMA, MoCA, MMSE, MDS-UPDRS | PCR amplification of the V5-V6 regions | Significant differences were found in microbiota composition of the gut between PD patients and healthy controls after adjusting for age, gender, and body mass index (BMI). The taxa class Clostridia, order Clostridiales, and family Ruminococcaceae in the gut microbiota were associated with weight and MMSE score. |
| 5 | Weis et al. (2021) [31] | Cohort study | Parkinson’s disease | N/A | Sequencing library for the V6-V7 regions of eukariotic 18S rRNA genes, PCR amplification | The proportions of the genera Aspergillus, Cercomonas, and Heteromita, as well as three unknown genus equivalent features of the phylum division Charophyta, the order Chromulinales, and the clade Opisthokonta, significantly decreased in PD patients. The relative abundance of genus Geotrichum significantly increased in PD patients. |
| 6 | Zhuang et al. (2020) [38] | Two-sample bi-directional Mendelian randomization analysis | Alzheimer’s disease, schizophrenia | N/A | Bacterial 16S rRNA gene sequencing | There was an increase in the relative abundance of Blautia associated with risk of AD. Elevated levels of the gut metabolite GABA were associated with a lower risk of AD. The increased proportion of the Enterobacteriaceae family and Enterobacteriales order were potentially related to a higher risk of schizophrenia (SCZ), while the Gammaproteobacteria class was associated with a lower risk of SCZ. Gut production of serotonin was potentially associated with a higher risk of SCZ. The increased relative abundance of the Bacilli class was associated with a higher risk of MDD. There was a lower relative abundance of the Erysipelotrichaceae family, the Erysipelotrichales order, and the Erysipelotrichia class and a higher relative abundance of unclassified Porphyromonadaceae in AD patients. MDD was related to increased proportion of unclassified Clostridiales, OTU16802 Bacteroides, and unclassified Prevotellaceae. SCZ was associated with an increased percentage of OTU10589 unclassified Enterobacteriaceae and decreased proportion of unclassified Erysipelotrichaceae. |
| 7 | Raghavan et al. (2023) [32] | Randomized pilot clinical study | Alzheimer’s disease, Parkinson’s disease | N/A | Metagenomic sequencing libraries, Polymerase Chain Reaction (PCR) | The phylum Firmicutes was the most abundant followed by Bacteroidetes. The abundance of Enterobacter decreased to almost zero, while the abundance of Prevotella increased in ASD patients. The abundance of Lactobacillus and Escherichia coli decreased in ASD patients. The proportions of Blautia spp., Coprobacillus sp. and several Clostridium spp. decreased. |
| 8 | Guo et al. (2022) [43] | Cohort study | Creutzfeldt–Jakob disease | CSF evaluation, MMSE, MoCA, CDR-SB | PCR amplification of the V3-V4 regions of bacterial 16S rRNA genes | It was found that at the phyla level, the relative abundance of Actinobacteria and Fusobacteria significantly increased in the CJD group. At the class level, there was a significant enrichment of Fusobacteriia, Actinobacteria, and Alphaproteobacteria in the CJD group. However, it was found that the abundance of Negativicutes decreased in the CJD group compared to healthy controls. At the family level, significant increases were found in the proportions of Fusobacteriaceae, Bifidobacterium, Succinivibrionaceae, and Enterococcaceae within the CJD group. At the genus level, Fusobacterium, Succinivibrio, Enterococcus, and Ruminococcus gnavus groups and Tyzzerella 4 were present at significantly higher levels in the CJD group, while the abundance of Coprococcus 1, Lachnospiraceae_ND3007, Pseudobutyrivibrio, Roseburia, and Holdemanella decreased in the CJD group. |
| 9 | Murros et al. (2021) [33] | Cohort study | Parkinson’s disease | Hoehn and Yahr Scale, MMSE | Polymerase Chain Reaction (PCR), bacterial 16S rRNA gene sequencing, HydA gene sequencing | It was found that all PD patients harbored Desulfovibrio bacteria in their gut microbiota, and the abundance of these bacteria increased in PD patients compared to control/healthy cohort. The concentration of Desulfovibrio species was correlated with the severity of PD. |
| 10 | Weis et al. (2019) [34] | Cohort study | Parkinson’s disease | Hoehn and Yahr Scale | Sequencing of the bacterial 16S rRNA genes (V4 and V5 regions), PCR amplification | Within the PD group, a relative decrease in bacterial taxa associated with health-promoting, anti-inflammatory, neuroprotective, or other beneficial effects on the epithelial barrier, such as Fusicatenibacter, was observed. The data confirm the previously reported effects of catechol-O-methyltransferase (COMT) inhibitors on the fecal microbiota of PD patients and suggest a possible effect of L-dopa medication on the relative abundance of several bacterial genera. |
| 11 | Wasser et al. (2020) [28] | Cohort study | Huntington’s disease | UHDRS, CAP, HD-CAB, Hopkins Verbal Learning Test-Revised, Symbol Digit Modalities Test, Trail Making Test, Paced tapping, Emotion Recognition Task, OTS of Cambridge | Sequencing of the bacterial 16S rRNA genes | Intestinal microbiome measurements revealed significant differences in the microbial communities between the combined Huntington’s disease gene expansion carrier (HDGEC) group and healthy controls. Major shifts in microbial community structure were also detected at the phylum and family levels, and functional pathways and enzymes that were affected in our HDGEC group were identified. Associations between gut bacteria, cognitive performance, and clinical outcomes were also discovered within the HDGEC group. |
| 12 | Vascellari et al. (2020) [35] | Cohort study | Parkinson’s disease | GC-MS analysis | Sequencing of the bacterial 16S rRNA genes (V3 and V4 regions) | The most significant changes within the PD group emphasized a reduction in bacterial taxa associated with anti-inflammatory/neuroprotective effects, particularly in the Lachnospiraceae family and its key members, such as Butyrivibrio, Pseudobutyrivibrio, Coprococcus, and Blautia. A direct evaluation of fecal metabolites revealed changes in several classes of metabolites. Most of the altered metabolites correlate strongly with the abundance of members of the Lachnospiraceae family, suggesting that these gut bacteria correlate with altered metabolic rates in PD. |
| 13 | Barone et al. (2021) [42] | Cohort study | Multiple sclerosis | MFIS-5, 6MWT, FFQ, paraclinical tests | Sequencing of the bacterial 16S rRNA genes (V3 and V4 regions), PCR amplification | B-HIPE led to a modulation of MS-typical dysbiosis with reduced concentrations of pathobionts and a replenishment of beneficial short-chain fatty acid producers. This partial restoration of a eubiotic profile may help to counteract the inflammatory tone typically seen in MS, which is supported by reduced circulating lipopolysaccharide levels and decreased populations of pro-inflammatory lymphocytes. Improved physical performance and reduced fatigue were also noted. |
| 14 | Hertel et al. (2019) [36] | Cohort study | Parkinson’s disease | UPDRS, mass spectrometry | N/A | The longitudinal trajectory of metabolites was associated with the interconversion of methionine and cysteine via cystathionine, which differed between PD patients and controls. Dopaminergic medication showed strong lipidomic signatures. Taurine-conjugated bile acids correlated with the severity of motor symptoms, while low levels of sulfated taurolithocholate were associated with PD incidence in the general population. Computational modeling predicted changes in sulfur metabolism, driven by A. muciniphila and B. wadsworthia, which is consistent with the changed metabolome. |
| 15 | Hegelmaier et al. (2020) [37] | Case–control study | Parkinson’s disease | UPDRS | Sequencing of the bacterial 16S rRNA genes (V1-V3 regions) | UDPRS III improved significantly and the levodopa-equivalent daily dose decreased after a vegetarian diet and fecal enema in a one-year follow-up. In addition, a significant correlation was observed between the diversity of the gut microbiome and the UPDRS III as well as the abundance of Ruminococcaceae. The abundance of Clostridiaceae also significantly reduced after an enema. |
| 16 | Zeng et al. (2020) [26] | Cohort study | Amyotrophic lateral sclerosis | El Escorial Criteria, UPLC | Sequencing of the bacterial 16S rRNA genes (V4 region), PCR amplification | The analysis showed an apparent change in the microbial structure of ALS patients, with Bacteroidetes upregulated at the phylum level and several microbes upregulated at the genus level, while Firmicutes were downregulated at the phylum level and Megamonas at the genus level compared to healthy controls. In addition, decreased gene function associated with metabolic pathways was observed in ALS patients. |
| 17 | Brenner et al. (2017) [40] | Cohort study | Amyotrophic lateral sclerosis | El Escorial Criteria | Quantification of 16S rDNA copy numbers by qRT-PCR, amplification of V3-V6 16S rDNA regions, PiCRUSt | Comparing the 2 groups, the diversity and abundance of the bacterial taxa on the different taxonomic levels as well as PiCRUSt-predicted metagenomes were almost indistinguishable. Significant differences between ALS patients and healthy controls were only observed with regard to the overall number of microbial species (operational taxonomic units) and in the abundance of uncultured Ruminococcaceae. |
| 18 | Nicholson et al. (2020) [27] | Case–control study | Amyotrophic lateral sclerosis | El Escorial Criteria, ALSFRS-R | Sequencing of the bacterial 16S rRNA genes (V4 region), Illumina HiSeq | The relative abundance of the dominant butyrate-producing bacteria, Eubacterium rectale and Roseburia intestinalis, was significantly lower in ALS patients compared to HC. These findings lend support to the inference that the gut microbiota could be a risk factor for ALS. |
| 19 | Zhou et al. (2021) [39] | Cohort study | Alzheimer’s disease | MMSE, MoCA, CDR, NPI | Sequencing of the bacterial 16S rRNA genes (V3 and V4 regions), PCR amplification, PiCRUSt | The fecal microbial composition of AD patients was quite distinct from that of HC. Bifidobacterium, Sphingomonas, Lactobacillus, and Blautia were enriched, while Odoribacter, Anaerobacterium, and Papillibacter were reduced. AD patients with NPS showed decreased abundance of Chitinophagaceae, Taibaiella, and Anaerobacterium compared with those without NPS. Functional pathways were different between AD and HC and between AD patients with and without NPS. A correlation analysis showed that Sphingomonas correlated negatively with MMSE; Anaerobacterium and Papillibacter correlated positively with MMSE and negatively with CDR. Cytophagia, Rhodospirillaceae, and Cellvibrio correlated positively with NPS, while Chitinophagaceae, Taibaiella, and Anaerobacterium correlated negatively with NPS. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koutsokostas, C.; Merkouris, E.; Goulas, A.; Aidinopoulou, K.; Sini, N.; Dimaras, T.; Tsiptsios, D.; Mueller, C.; Nystazaki, M.; Tsamakis, K. Gut Microbes Associated with Neurodegenerative Disorders: A Comprehensive Review of the Literature. Microorganisms 2024, 12, 1735. https://doi.org/10.3390/microorganisms12081735
Koutsokostas C, Merkouris E, Goulas A, Aidinopoulou K, Sini N, Dimaras T, Tsiptsios D, Mueller C, Nystazaki M, Tsamakis K. Gut Microbes Associated with Neurodegenerative Disorders: A Comprehensive Review of the Literature. Microorganisms. 2024; 12(8):1735. https://doi.org/10.3390/microorganisms12081735
Chicago/Turabian StyleKoutsokostas, Christos, Ermis Merkouris, Apostolos Goulas, Konstantina Aidinopoulou, Niki Sini, Theofanis Dimaras, Dimitrios Tsiptsios, Christoph Mueller, Maria Nystazaki, and Konstantinos Tsamakis. 2024. "Gut Microbes Associated with Neurodegenerative Disorders: A Comprehensive Review of the Literature" Microorganisms 12, no. 8: 1735. https://doi.org/10.3390/microorganisms12081735
APA StyleKoutsokostas, C., Merkouris, E., Goulas, A., Aidinopoulou, K., Sini, N., Dimaras, T., Tsiptsios, D., Mueller, C., Nystazaki, M., & Tsamakis, K. (2024). Gut Microbes Associated with Neurodegenerative Disorders: A Comprehensive Review of the Literature. Microorganisms, 12(8), 1735. https://doi.org/10.3390/microorganisms12081735