
Characterization of an Acetogenin-Carrying Nanosuspension and Its Effect on Bacteria of Interest in the Poultry Industry

 , ,
, ,  , , and
, , and 
Abstract
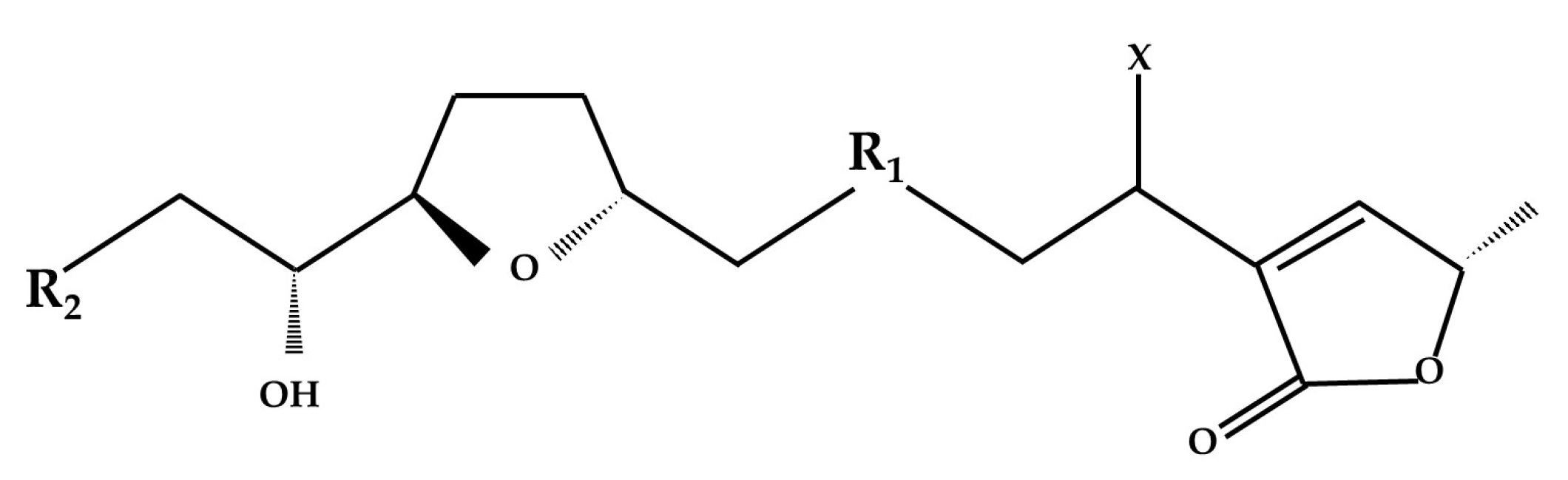
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Strains
2.3. Preparation of Acetogenin-Loaded Nanosuspensions
2.4. Particle Size, Polydispersity Index, and Zeta Potential of NSps-βCDSL-ACGs
2.5. Spectroscopic Characterization of NSps-βCDSL-ACGs
2.5.1. Fourier Transform Infrared Spectroscopy (FTIR)
2.5.2. Diffuse Reflectance UV-VIS Spectroscopy
2.5.3. Transmission Electron Microscopy (TEM)
2.6. Determination of the Antimicrobial Activity of NSps-βCDSL-ACGs
2.6.1. Determination of the Mean Inhibitory Concentration (IC50) and Minimum Inhibitory Concentration (MIC) of NSps-βCDSL-ACGs Using a Regression Analysis
2.6.2. Lethality of NSps-βCDSL-ACGs Against S. Enteritidis, S. Typhimurium, S. Infantis, and S. gallolyticus
2.7. Statistical Analysis
3. Results and Discussion
3.1. Particle Size, Polydispersity Index, and Z-Potential of NSps-βCDSL-ACGs
3.2. Spectroscopic Characterization of Isolated ACGs, Complex βCDSL, and NSps-βCDSL-ACGs
3.2.1. Fourier Transform Infrared Spectroscopy (FTIR)
3.2.2. Diffuse Reflectance Spectroscopy–UV-VIS Analysis
3.2.3. Transmission Electron Microscopy (TEM)
3.3. Determination of the Antimicrobial Activity of NSps-βCDSL-ACGs
3.3.1. Microdilution Plate for the Determination of Bacterial Inhibition of NSps-βCDSL-ACGs
3.3.2. Determination of the Mean Inhibitory Concentration (IC50) and Minimum Inhibitory Concentration (MIC) of NSps-βCDSL-ACGs Using Regression Analysis
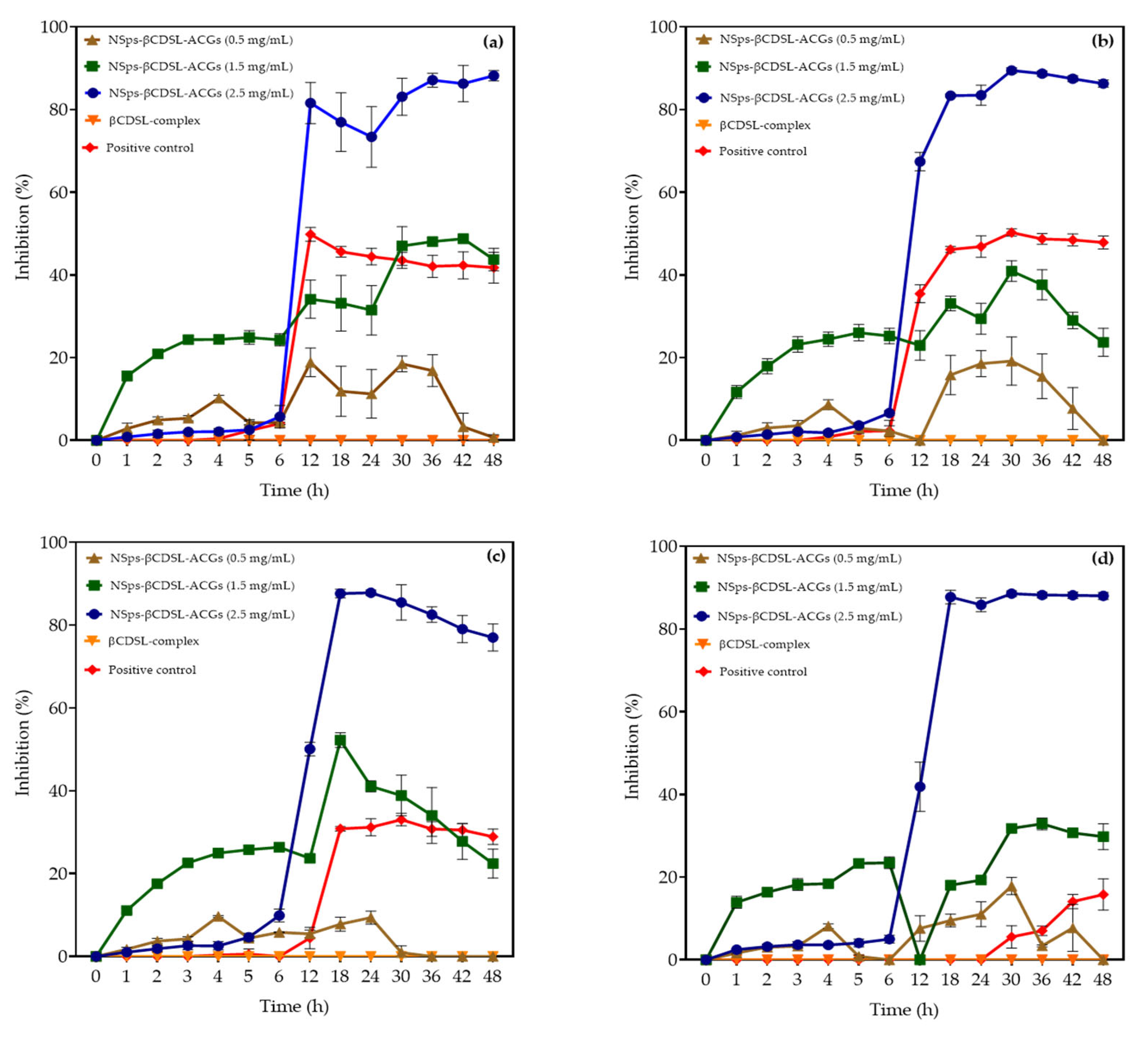
3.4. Lethality of NSps-βCDSL-ACGs Against S. Enteritidis, S. Typhimurium, S. Infantis, and S. gallolyticus
4. Conclusions
5. Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bermejo, A.; Figadère, B.; Zafra-Polo, M.C.; Barrachina, I.; Estornell, E.; Cortes, D. Acetogenins from Annonaceae: Recent progress in isolation, synthesis and mechanisms of action. Nat. Prod. Rep. 2005, 22, 269–303. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Hernández, G.; Zepeda-Vallejo, L.G.; García-Magaña, M.L.; López-García, U.M.; Aguilera-Aguirre, S.; Montalvo-González, E. Acetogenin content from soursop (Annona muricata L.) fruit seeds: Evaluation of different extraction conditions. Biotec 2022, 24, 12–19. [Google Scholar] [CrossRef]
- Aguilar-Hernández, G.; López-Romero, B.A.; Huerta-Castellanos, I.; Tellez-Isaías, G.; Montalvo-González, E. Acetogenins from Annona muricata as Antimicrobial Agents; IntechOpen: London, UK, 2024. [Google Scholar] [CrossRef]
- Bravo-Alfaro, D.A.; Montalvo-González, E.; Zapien-Macías, J.M.; Sampieri-Moran, J.M.; García, S.H.; Luna-Bárcenas, G. Annonaceae acetogenins: A potential treatment for gynecological and breast cancer. Fitoterapia 2024, 178, 106187. [Google Scholar] [CrossRef]
- Hong, J.; Li, Y.; Li, Y.; Xiao, Y.; Kuang, H.; Wang, X. Annonaceous acetogenins nanosuspensions stabilized by PCL-PEG block polymer: Significantly improved antitumor efficacy. Int. J. Nanomed. 2016, 11, 3239–3253. [Google Scholar] [CrossRef]
- Hong, J.; Sun, Z.; Li, Y.; Guo, Y.; Liao, Y.; Liu, M.; Wang, X. Folate-modified Annonaceous acetogenins nanosuspensions and their improved antitumor efficacy. Int. J. Nanomed. 2017, 12, 5053–5067. [Google Scholar] [CrossRef]
- Li, H.; Li, Y.; Ao, H.; Bi, D.; Han, M.; Guo, Y.; Wang, X. Folate-targeting annonaceous acetogenins nanosuspensions: Significantly enhanced antitumor efficacy in HeLa tumor-bearing mice. Drug Deliv. 2018, 25, 880–887. [Google Scholar] [CrossRef]
- Han, J.; Zhou, X.; Fu, J.; Gao, G.; Zuo, C.; Guo, Y.; Han, M.; Wang, X. Annonaceous acetogenins nanosuspensions stabilized by poloxamer 188: Preparation, properties and in vivo evaluation. J. Drug Deliv. Sci. Technol. 2021, 66, 102376. [Google Scholar] [CrossRef]
- López-Romero, B.A.; Téllez-Isaías, G.; Luna-Bárcenas, G.; Zamudio-Ojeda, A.; Velázquez-Juárez, G.; Garcia-Magaña, M.L.; Montalvo-González, E. Nanosuspensions as carriers of Annona muricata acetogenins: Antibacterial activity against Enterococcus faecalis and Listeria monocytogenes. CYTA J. Food 2023, 21, 771–780. [Google Scholar] [CrossRef]
- Montalvo-González, E.; Tellez-Isaías, G.; López-Romero, B.A.; Zamudio-Ojeda, A.; Pérez-Ladrón de Guevara, H.; Castellanos-Huerta, I.; Pérez-Larios, A.; Hernández-Aguilar, G. Polyethylene glycol/soy lecithin-nanosuspensions as carriers of acetogenins: Optimization, characterization, antibacterial and hemolytic activity. Drug Deliv. Sci. Technol. 2024, 91, 105228. [Google Scholar] [CrossRef]
- Long, J.; Song, J.; Zhang, X.; Deng, M.; Xie, L.; Zhang, L.; Li, X. Tea saponins as natural stabilizers for the production of hesperidin nanosuspensions. Int. J. Pharm. 2020, 583, 119406. [Google Scholar] [CrossRef]
- Aguilar-Hernández, G.; López Romero, B.A.; Pérez-Larios, A.; Ruvalcaba-Gómez, J.M.; Castellanos-Huerta, I.; Téllez-Isaías, G.; Petrone-García, V.M.; Anaya-Esparza, L.M.; Montalvo-González, E. Antibacterial activity of crude extract and purified acetogenins from Annona muricata seeds. Appl. Sci. 2023, 13, 558. [Google Scholar] [CrossRef]
- Hossain, M.J.; Attia, Y.; Ballah, F.M.; Islam, M.S.; Sobur, M.A.; Islam, M.A.; Ievy, S.; Rahman, A.; Nishiyama, A.; Islam, M.S. Zoonotic significance and antimicrobial resistance in Salmonella in poultry in Bangladesh for the period of 2011–2021. Zoonotic Dis. 2021, 1, 3–24. [Google Scholar] [CrossRef]
- Hao, H.; Cheng, G.; Iqbal, Z.; Ai, X.; Hussain, H.I.; Huang, Y.Z. Benefits and risks of antimicrobial use in food-producing animals. Front. Microbiol. 2014, 5, 288. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Van, B.; Emma, E.; Glennon, D.; Chen, M.G.; Timothy, P.; Robinson, G.; Simon, A.L.; Sebastian, B.; Ramanan, L. Reducing antimicrobial use in food animals. Science 2017, 357, 1350–1352. [Google Scholar] [CrossRef]
- Zhao, C.; Wang, Y.; Mulchandani, R.; Thomas, P.; Boeckel, V. Global surveillance of antimicrobial resistance in animals intended for human consumption using drug maps. Nat. Commun. 2024, 15, 763. [Google Scholar] [CrossRef]
- Shivaning, K.N.; Benakabhat, M.C.; Agalagandi, G.S.; Hiremath, J.; Shivanagowda, P.G.; Barbuddhe, S.B. Prevalence of Salmonella serotypes S. enteritidis and S. typhimurium in poultry and poultry products. J. Food Saf. 2020, 40, 12852. [Google Scholar] [CrossRef]
- Tsivelika, N.; Sarrou, E.; Gusheva, K.; Pankou, C.; Koutsos, T.; Chatzopoulou, P.; Mavromatis, A. Phenotypic variation of wild Chamomile (Matricaria chamomilla L.) populations and their evaluation for medicinally important essential oil. Biochem. Syst. Ecol. 2018, 80, 21–28. [Google Scholar] [CrossRef]
- Naiel, M.A.; Ismael, N.E.; Shehata, S.A. Rosemary leaf powder–supplemented diet enhances performance, antioxidant properties, immune status, and resistance against bacterial diseases in Nile Tilapia (Oreochromis niloticus). Agric. Food Sci. 2019, 511, 734264. [Google Scholar] [CrossRef]
- Khafaga, A.F.; Naiel, M.A.E.; Dawood, M.A.O.; Abdel-Latif, H.M.R. Dietary Origanum vulgare essential oil attenuates cypermethrin-induced biochemical changes, oxidative stress, histopathological alterations, apoptosis, and reduces DNA damage in Common carp [Cyprinus carpio]. Aquat. Toxicol. 2020, 228, 105624. [Google Scholar] [CrossRef]
- Seidavi, A.; Tavakoli, M.; Asroosh, F.; Colin, G.S.; Mohamed, E.A.E.; Mohamed, A.E.N.; Ayman, E.T.; Lofti, A.; Khaled, A.E.; Ayman, A.S. Antioxidant and antimicrobial activities of phytonutrients as antibiotic substitutes in poultry feeds. Environ. Sci. Pollut. Res. 2022, 29, 5006–5031. [Google Scholar] [CrossRef]
- López-Romero, B.A.; Luna-Bárcenas, G.; García-Magaña, M.L.; Anaya-Esparza, L.M.; Zepeda-Vallejo, L.G.; López-García, U.M.; Ortiz-Basurto, R.I.; Aguilar-Hernández, G.; Pérez-Larios, A.; Montalvo-González, E. Extraction of acetogenins using thermosonication-assisted extraction from Annona muricata seeds and their antifungal activity. Molecules 2022, 27, 6045. [Google Scholar] [CrossRef] [PubMed]
- George, M.M.; Sayes, C.; Zechmann, B. Hyperspectral imaging as a tool to detect and characterize nanoparticles in complex Biofluids. Microsc. Microanal. 2020, 26, 2748–2750. [Google Scholar] [CrossRef]
- Kimpe, A.; Decostere, A.; Martel, A.; Haesebrouck, F.; Devriese, L.A. Prevalence of antimicrobial resistance among pigeon isolates of Streptococcus gallolyticus, Escherichia coli and Salmonella enterica serotype Typhimurium. Avian Pathol. J. WVPA 2002, 31, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Habrun, B.; Šimpraga, B.; Kompes, G.; Krstulović, F. Antimicrobial resistance and serotyping of Salmonella enterica subsp. enterica isolated from poultry in Croatia. Vet. Arhiv. 2012, 82, 371–381. [Google Scholar]
- Provenzani, R.; San-Martin-, P.; Galindo-Hassan, G. Multisubstituted pyrimidines effectively inhibit bacterial growth and biofilm formation of Staphylococcus aureus. Sci. Rep. 2021 11, 7931. [CrossRef]
- Bloomfield, S.F. Methods for assessing antimicrobial activity. In Mechanisms of Action of Chemical Biocides: Their Study and Exploitation; Denyer, S.P., Hugo, W.B., Eds.; Blackwell Scientific Publications: Hoboken, NJ, USA, 2024; pp. 1–6. [Google Scholar]
- Zielińska, A.; Carreiró, F.; Oliveira, A.M.; Neves, A.; Pires, B.; Venkatesh, D.N.; Durazzo, A.; Lucarini, M.; Eder, P.; Silva, A.M. Polymeric nanoparticles: Production, characterization, toxicology and ecotoxicology. Molecules 2020, 25, 3731. [Google Scholar] [CrossRef]
- Hamed, R.; Alhadidi, H. Dissolved microneedles loaded with HFI minoxidil nanosuspension for hair growth. AAPS PharmSciTech 2024, 25, 75. [Google Scholar] [CrossRef]
- Li, M.; Li, S.; Li, Y.; Li, X.; Yang, G.; Li, M.; Xie, Y.; Su, W.; Wu, J.; Jia, L.; et al. Cationic liposomes co-deliver chemotherapeutics and siRNA for the treatment of breast cancer. Eur. J. Med. Chem. 2022, 233, 114198. [Google Scholar] [CrossRef]
- Hidalgo, J.R.; Neske, A.; Iramain, M.A.; Alvarez, P.E.; Bongiorno, P.L.; Brandán, S.A. Experimental isolation and spectroscopic characterization of squamocin acetogenin combining FT-IR, FT-Raman and UV–Vis spectra with DFT calculations. J. Mol. Struct. 2020, 1219, 128610. [Google Scholar] [CrossRef]
- Aguilar-Villalva, R.; Molina, G.A.; España-Sánchez, B.L.; Díaz-Peña, L.F.; Elizalde-Mata, A.; Valerio, E.; Estevez, M. Antioxidant capacity and antibacterial activity from Annona cherimola phytochemicals by ultrasound-assisted extraction and its comparison to conventional methods. Arab. J. Chem. 2021, 14, 103239. [Google Scholar] [CrossRef]
- Ferreira, G.G.; Quaresma, A.C.S.; Brandão, D.L.D.N.; Marinho, A.M.D.R.; Siqueira, J.E.D.S.; Correa, K.L.; Silva-Júnior, J.O.C.; Percario, S.; Dolabela, M.F. Evaluation of genotoxicity and toxicity of Annona muricata L. seeds and in silico studies. Molecules 2023, 28, 231. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Arruda, T.; Assis Silva, R.R.; Suprani-Marques, C.; Robledo-Fialho, A.; Campos-Bernardes, P.; Veloso de Oliveira, T.; Olavo de Oliveira, S.; Muranyi, P.; Ferreira-Soares, N.F. β-cyclodextrin versus hydroxypropyl-β-cyclodextrin: Is inclusion complexation a suitable alternative to improve the properties of hop-derived β-acids. Food Hydrocoll. 2024, 14, 109622. [Google Scholar] [CrossRef]
- Wang, X.; Luo, Z.; Xiao, Z. Preparation, characterization and thermal stability of β-cyclodextrin/soy lecithin inclusion complex. Carbohr. Polym. 2014, 101, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Zafra-Polo, M.C.; González, M.C.; Estornell, E.; Sahpaz, S.; Cortes, D. Acetogenins from Annonaceae, inhibitors of mitochondrial complex I. Phytochemistry 1996, 42, 253–271. [Google Scholar] [CrossRef] [PubMed]
- Pinto, N.C.C.; Campos, L.M.; Evangelista, A.C.S.; Lemos, A.O.; Silva, T.P.; Melo, R.C.N.; Fabri, R.L. Antimicrobial Annona muricata L. (soursop) extract targets the cell membranes of Gram-positive and Gram-negative bacterias. Ind. Crops Prod. 2017, 107, 332–340. [Google Scholar] [CrossRef]
- Zhang, F.; Cheng, W. The mechanism of bacterial resistance and potential bacteriostatic strategies. Antibiotics 2022, 11, 1215. [Google Scholar] [CrossRef]
- Thung, T.Y.; Mahyudin, N.A.; Basri, D.F.; Wan Mohamed Radzi, C.W.J.; Nakaguchi, Y.; Nishibuchi, M.; Radu, S. Prevalence and antibiotic resistance of Salmonella enteritidis and Salmonella typhimurium in raw chicken meat at retail markets in Malaysia. Poult. Sci. 2016, 95, 1888–1893. [Google Scholar] [CrossRef]
- Eleraky, N.E.; Allam, A.; Hassan, S.B.; Omar, M.M. Nanomedicine fight against antibacterial resistance: An overview of the recent pharmaceutical innovations. Pharmaceutics 2020, 12, 142. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | NSps-βCDSL-ACGs | ||
|---|---|---|---|
| 1 mg/mL | 3 mg/mL | 5 mg/mL | |
| Particle size (nm) | 207.34 ± 7.63 c | 228.18 ± 10.05 b | 239.12 ± 3.82 a |
| PDI | 0.178 ± 0.024 b | 0.136 ± 0.021 a | 0.150 ± 0.029 ab |
| Z potential (mV) | −50.36 ± 0.90 a | −49.02 ± 1.01 b | −47.18 ± 0.58 c |
| Bacteria | IC50 (mg/mL) | MIC (mg/mL) | R2 |
|---|---|---|---|
| Salmonella Enteritidis | 1.74 | 0.0919 | 0.9985 |
| Salmonella Typhimurium | 2.00 | 0.0519 | 0.9980 |
| Salmonella Infantis | 1.98 | 0.053 | 0.9952 |
| Streptococcus gallolyticus | 1.92 | 0.121 | 0.9983 |
| Bacteria | * Positive Control | NSps-βCDSL-ACGs | ||
|---|---|---|---|---|
| 0.5 mg/mL | 1.5 mg/ mL | 2.5 mg/mL | ||
| S. Enteritidis | 1.08 ± 0.03 c | 1.07 ± 0.47 b | 1.70 ± 0.13 a | 1.54 ± 0.15 a |
| S. Typhimurium | 1.29 ± 0.35 c | 1.88 ± 0.23 b | 3.03 ± 0.32 a | 3.19 ± 0.10 a |
| S. Infantis | 0.73 ± 0.63 ab | 0.16 ± 0.03 b | 0.25 ± 0.60 ab | 0.98 ± 0.30 a |
| S. gallolyticus | 1.42 ± 0.39 a | 0.65 ± 0.58 ab | 1.10 ± 0.25 ab | 1.16 ± 0.15 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Romero, B.A.; Aguilar-Hernández, G.; Hargis, B.M.; García-Magaña, M.d.L.; López-García, U.M.; Ortiz-Basurto, R.I.; Zamudio-Ojeda, A.; Latorre, J.D.; Montalvo-González, E. Characterization of an Acetogenin-Carrying Nanosuspension and Its Effect on Bacteria of Interest in the Poultry Industry. Microorganisms 2025, 13, 18. https://doi.org/10.3390/microorganisms13010018
López-Romero BA, Aguilar-Hernández G, Hargis BM, García-Magaña MdL, López-García UM, Ortiz-Basurto RI, Zamudio-Ojeda A, Latorre JD, Montalvo-González E. Characterization of an Acetogenin-Carrying Nanosuspension and Its Effect on Bacteria of Interest in the Poultry Industry. Microorganisms. 2025; 13(1):18. https://doi.org/10.3390/microorganisms13010018
Chicago/Turabian StyleLópez-Romero, Brandon A., Gabriela Aguilar-Hernández, Billy M. Hargis, María de Lourdes García-Magaña, Ulises M. López-García, Rosa I. Ortiz-Basurto, Adalberto Zamudio-Ojeda, Juan D. Latorre, and Efigenia Montalvo-González. 2025. "Characterization of an Acetogenin-Carrying Nanosuspension and Its Effect on Bacteria of Interest in the Poultry Industry" Microorganisms 13, no. 1: 18. https://doi.org/10.3390/microorganisms13010018
APA StyleLópez-Romero, B. A., Aguilar-Hernández, G., Hargis, B. M., García-Magaña, M. d. L., López-García, U. M., Ortiz-Basurto, R. I., Zamudio-Ojeda, A., Latorre, J. D., & Montalvo-González, E. (2025). Characterization of an Acetogenin-Carrying Nanosuspension and Its Effect on Bacteria of Interest in the Poultry Industry. Microorganisms, 13(1), 18. https://doi.org/10.3390/microorganisms13010018