
Where Biology Meets Engineering: Scaling Up Microbial Nutraceuticals to Bridge Nutrition, Therapeutics, and Global Impact

 ,
,  ,
,
Abstract
- Introduction
- Microbial production of nutraceuticals
1. Probiotics
2. Prebiotics and Postbiotics
3. Polyunsaturated Fatty Acids
4. Exopolysaccharides (EPSs)
4.1. Microbial EPS Biosynthesis and Metabolic Pathways
4.2. Industrial Applications and Commercial Value
4.3. Future Perspectives and Next-Generation EPS Production
5. Oligosaccharides
6. Low-Calorie Sugars
7. Lactic Acid
8. Polyphenols
9. Selenium
10. Vitamins
10.1. Riboflavin (Vitamin B12)
10.2. Vitamin C (Ascorbic Acid)
10.3. Coenzyme Q10 (Ubiquinone)
10.4. Vitamin B7 (Biotin)
10.5. Carotenoids (e.g., β-Carotene)
10.6. Vitamin B9 (Folate)
10.7. Vitamin E (Tocopherol)
10.8. Vitamin K
11. Glutathione and α-Ketoglutarate
12. Gamma-Aminobutyric Acid (GABA)
13. Microbial Pigments
14. Bioactive Peptides
14.1. Mechanisms of Bioactivity
14.1.1. Antihypertensive Activity
14.1.2. Antioxidant Mechanism
14.1.3. Immunomodulation
14.1.4. Antimicrobial Activity
14.2. Production and Industrial Applications
14.3. Challenges and Future Perspectives
14.3.1. Low Yield and Prolonged Fermentation Time
14.3.2. Sensitivity to Environmental Factors
14.3.3. Challenges in Purification and Downstream Processing
15. Comparative Analysis of Traditional vs. Microbial Production
16. Relevance and Challenges of Commercial Nutraceutical Production
16.1. Strain Selection for Nutraceutical Production
16.2. Synthetic Biology and Heterologous Expression
16.3. Advances in CRISPR/Cas9 Technology
16.4. Adaptive Evolution: Enhancing Microbial Efficiency
16.5. Fermentation Technologies for Nutraceutical Production
16.6. Process Optimization
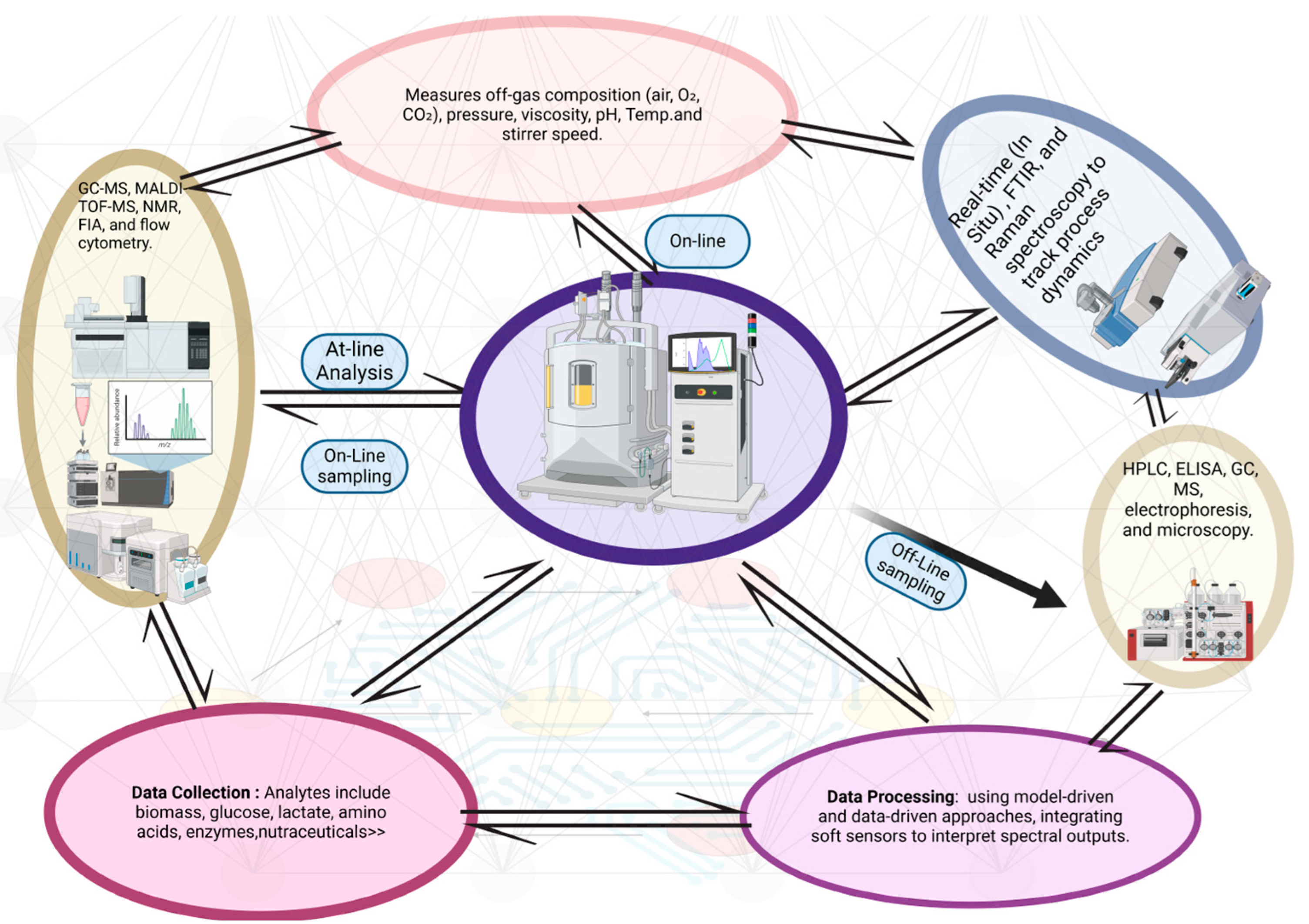
17. Microbial Intelligence (MI)
Unlocking the Potential of Microbial Nutraceuticals
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoti, G.; Matencio, A.; Rubin Pedrazzo, A.; Cecone, C.; Appleton, S.L.; Khazaei Monfared, Y.; Caldera, F.; Trotta, F. Nutraceutical Concepts and Dextrin-Based Delivery Systems. Int. J. Mol. Sci. 2022, 23, 4102. [Google Scholar] [CrossRef]
- Fixsen, A. Disordered Eating, Food Landscapes, and Public Health. In The Construction of Eating Disorders. The Politics of Mental Health and Illness; Palgrave Macmillan: Cham, Switzerland, 2024. [Google Scholar] [CrossRef]
- Prabhu, P.P.; Mohanty, B.; Lobo, C.L.; Balusamy, S.R.; Shetty, A.; Perumalsamy, H.; Mahadev, M.; Mijakovic, I.; Dubey, A.; Singh, P. Harnessing the nutriceutics in early-stage breast cancer: Mechanisms, combinational therapy, and drug delivery. J. Nanobiotechnol. 2024, 22, 574. [Google Scholar] [CrossRef]
- Chandra, S.; Saklani, S.; Kumar, P.; Kim, B. Nutraceuticals: Pharmacologically Active Potent Dietary Supplements. Biomed. Res. Int. 2022, 2022, 2051017. [Google Scholar] [CrossRef]
- Jagtap, S. Microbial Bioprospecting for Nutraceuticals and Value-Added Compounds. Bioprospecting Microb. Divers. 2022, 2, 23–45. [Google Scholar]
- Rusu, A.V.; Trif, M.; Rocha, J.M. Microbial Secondary Metabolites via Fermentation Approaches for Dietary Supplementation Formulations. Molecules 2023, 28, 6020. [Google Scholar] [CrossRef]
- Chaturvedi, V.; Goswami, R.K.; Verma, P. Genetic Engineering for Enhancement of Biofuel Production in Microalgae. In Biorefineries: A Step Towards Renewable and Clean Energy; Springer: Singapore, 2021; pp. 539–559. [Google Scholar]
- Ailioaie, L.M.; Litscher, G. Probiotics, Photobiomodulation, and Disease Management: Controversies and Challenges. Int. J. Mol. Sci. 2021, 22, 4942. [Google Scholar] [CrossRef]
- Milner, E.; Stevens, B.; An, M.; Lam, V.; Ainsworth, M.; Dihle, P.; Stearns, J.; Dombrowski, A.; Rego, D.; Segars, K. Utilizing Probiotics for the Prevention and Treatment of Gastrointestinal Diseases. Front. Microbiol. 2021, 12, 689958. [Google Scholar] [CrossRef]
- Kumar, V.; Naik, B.; Kumar, A.; Khanduri, N.; Rustagi, S.; Kumar, S. Probiotics Media: Significance, Challenges, and Future Perspective—A Mini Review. Food Prod. Process. Nutr. 2022, 4, 17. [Google Scholar] [CrossRef]
- Pereira, A.L.F.; Rodrigues, S. Turning Fruit Juice Into Probiotic Beverages. In Fruit Juices; Academic Press: New York, NY, USA, 2018; pp. 279–287. [Google Scholar] [CrossRef]
- Liang, Y.; Liu, M.; Pu, J.; Zhu, Z.; Gao, Z.; Zhou, Q.; Li, P. Probiotics and Their Metabolites Ameliorate Inflammatory Bowel Disease: A Critical Review. Infect. Microbes Dis. 2021, 3, 4–13. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Bron, P.A.; Kleerebezem, M.; Brummer, R.J.; Cani, P.D.; Mercenier, A.; MacDonald, T.T.; Wells, J.M. Can Probiotics Modulate Human Disease by Impacting Intestinal Barrier Function? Br. J. Nutr. 2017, 117, 93–107. [Google Scholar] [CrossRef]
- Rocha, C.S.; Coelho, M.C.; Souza, E.C.; de Macedo, F.H.; Leite, J.R.S.; Passos, T.S. Probiotics and Its Applications in Food and Health: A Review. Trends Food Sci. Technol. 2021, 110, 205–221. [Google Scholar] [CrossRef]
- Szajewska, H.; Skórka, A. Lactobacillus rhamnosus GG in the Prevention of Nosocomial Diarrhea in Children: A Randomized, Double-Blind, Placebo-Controlled Trial. J. Pediatr. 2009, 147, 304–308. [Google Scholar] [CrossRef]
- Nobaek, S.; Johansson, M.L.; Molin, G.; Ahrné, S.; Jeppsson, B. Alteration of Intestinal Microflora is Associated with Reduction in Abdominal Bloating and Pain in Patients with Irritable Bowel Syndrome. Am. J. Gastroenterol. 2000, 95, 1231–1238. [Google Scholar] [CrossRef]
- Fenster, K.; Freeburg, B.; Hollard, C.; Wong, C.; Rønhave Laursen, R.; Ouwehand, A.C. The Production and Delivery of Probiotics: A Review of a Practical Approach. Microorganisms 2019, 7, 83. [Google Scholar] [CrossRef]
- da Silva, T.F.; Glória, R.A.; Americo, M.F.; Freitas, A.S.; de Jesus, L.C.L.; Barroso, F.A.L.; Laguna, J.G.; Coelho-Rocha, N.D.; Tavares, L.M.; le Loir, Y.; et al. Unlocking the Potential of Probiotics: A Comprehensive Review on Research, Production, and Regulation of Probiotics. Probiotics Antimicrob. Proteins 2024, 16, 1687–1723. [Google Scholar] [CrossRef]
- Sloan, E.A. Top 10 Functional Food Trends. IFT. 2022. Available online: https://www.ift.org/news-and-publications/food-technology-magazine/issues/2022/april/features/top-10-functional-food-trends (accessed on 1 January 2024).
- Marco, M.L.; Sanders, M.E.; Gänzle, M.; Arrieta, M.C.; Cotter, P.D.; De Vuyst, L.; Hill, C.; Holzapfel, W.; Lebeer, S.; Merenstein, D.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on Fermented Foods. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 196–208. [Google Scholar] [CrossRef]
- Cha, K.H.; Lee, E.H.; Yoon, H.S.; Lee, J.H.; Kim, J.Y.; Kang, K.; Park, J.-S.; Jin, J.B.; Ko, G.; Pan, C.-H. Effects of Fermented Milk Treatment on Microbial Population and Metabolomic Outcomes in a Three-Stage Semi-Continuous Culture System. Food Chem. 2018, 263, 216–224. [Google Scholar] [CrossRef]
- Hwang, C.F.; Chen, J.N.; Huang, Y.T.; Mao, Z.Y. Biomass Production of Lactobacillus plantarum LP02 Isolated from Infant Feces with Potential Cholesterol-Lowering Ability. Afr. J. Biotechnol. 2011, 10, 7010–7020. [Google Scholar]
- Yuste, A.; Arosemena, E.L.; Calvo, M. Study of the Probiotic Potential and Evaluation of the Survival Rate of Lactiplantibacillus plantarum Lyophilized as a Function of Cryoprotectant. Sci. Rep. 2021, 11, 19078. [Google Scholar] [CrossRef]
- Ji, J.; Jin, W.; Liu, S.-J.; Jiao, Z.; Li, X. Probiotics, Prebiotics, and Postbiotics in Health and Disease. MedComm 2023, 4, e420. [Google Scholar] [CrossRef]
- Elazzazy, A.M.; Mobarki, M.O.; Baghdadi, A.M.; Bataweel, N.M.; Al-Hejin, A.M. Optimization of Culture Conditions and Batch Process Control for the Augmented Production of Bacteriocin by Bacillus Species. Microorganisms 2024, 12, 651. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef]
- Gamal, R.; Shreadah, M.A. Marine Microalgae and Their Industrial Biotechnological Applications: A Review. J. Genet. Eng. Biotechnol. 2024, 22, 100407. [Google Scholar] [CrossRef]
- Dourou, M.; Dritsas, P.; Baeshen, M.N.; Elazzazy, A.; Al-Farga, A.; Aggelis, G. High-added value products from microalgae and prospects of aquaculture wastewaters as microalgae growth media. FEMS Microbiol. Lett. 2020, 367, fnaa081. [Google Scholar] [CrossRef]
- Kousparou, C.; Fyrilla, M.; Stephanou, A.; Patrikios, I. DHA/EPA (Omega-3) and LA/GLA (Omega-6) as Bioactive Molecules in Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 10717. [Google Scholar] [CrossRef]
- Albracht-Schulte, K.; Gonzalez, S.; Jackson, A.; Wilson, S.; Ramalingam, L.; Kalupahana, N.S.; Moustaid-Moussa, N. Eicosapentaenoic Acid Improves Hepatic Metabolism and Reduces Inflammation Independent of Obesity in High-Fat-Fed Mice and in HepG2 Cells. Nutrients 2019, 11, 599. [Google Scholar] [CrossRef]
- Qian, C.; Wang, Q.; Qiao, Y.; Xu, Z.; Zhang, L.; Xiao, H.; Lin, Z.; Wu, M.; Xia, W.; Yang, H.; et al. Arachidonic Acid in Aging: New Roles for Old Players. J. Adv. Res. 2024, 4, S2090-1232(24)00180-2. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mohanty, A.K.; Rao, U.; Poddar, A. Microbial Polyunsaturated Fatty Acids (PUFAs) for Human Health: A Comprehensive Review of Evidence from In Vivo Preclinical Models. Discov. Appl. Sci. 2024, 6, 524. [Google Scholar] [CrossRef]
- Kothri, M.; Mavrommati, M.; Elazzazy, A.M.; Baeshen, M.N.; Moussa, T.A.A.; Aggelis, G. Microbial sources of polyunsaturated fatty acids (PUFAs) and the prospect of organic residues and wastes as growth media for PUFA-producing microorganisms. FEMS Microbiol. Lett. 2020, 367, fnaa028. [Google Scholar] [CrossRef]
- Panchal, S.K.; Heimann, K.; Brown, L. Improving Undernutrition with Microalgae. Nutrients 2024, 16, 3223. [Google Scholar] [CrossRef]
- Yu, B.S.; Pyo, S.; Lee, J.; Han, K. Microalgae: A Multifaceted Catalyst for Sustainable Solutions in Renewable Energy, Food Security, and Environmental Management. Microb. Cell Fact. 2024, 23, 308. [Google Scholar] [CrossRef]
- Phukon, L.C.; Sudhir, P.; Singh, A.; Pandey, A.K.; Rai, A.K. Microbial Bioprocesses for Production of Nutraceuticals and Functional Foods. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2022; pp. 1–29. ISBN 978-0128235065. [Google Scholar]
- Kalampounias, G.; Dritsas, P.; Karayannis, D.; Androutsopoulou, T.; Gardeli, C.; Papanikolaou, S.; Aggelis, G.; Katsoris, P. Polyunsaturated Fatty Acids from Thamnidium elegans and Mortierella alpina Suppress Prostate Cancer Cell Proliferation and Migration. Fermentation 2024, 10, 578. [Google Scholar] [CrossRef]
- Friedlander, J.; Tsakraklides, V.; Kamineni, A.; Greenhagen, E.H.; Consiglio, A.L.; MacEwen, K.; Crabtree, D.V.; Afshar, J.; Nugent, R.L.; Hamilton, M.A.; et al. Engineering of a high lipid producing Yarrowia lipolytica strain. Biotechnol. Biofuels 2016, 9, 1. [Google Scholar] [CrossRef]
- Xu, P.; Qiao, K.; Stephanopoulos, G. Engineering oxidative stress defense pathways to build a robust lipid production platform in Yarrowia lipolytica. Biotechnol. Bioeng. 2017, 114, 1521–1530. [Google Scholar] [CrossRef]
- El-Baz, H.A.; Elazzazy, A.M.; Saleh, T.S.; Dourou, M.; Mahyoub, J.A.; Baeshen, M.N.; Madian, H.R.; Aggelis, G. Enzymatic Synthesis of Glucose Fatty Acid Esters Using SCOs as Acyl Group-Donors and Their Biological Activities. Appl. Sci. 2021, 11, 2700. [Google Scholar] [CrossRef]
- Anschau, A.; Xavier, M.C.A.; Hernalsteens, S.; Franco, T.T. Effect of feeding strategies on lipid production by Lipomyces starkeyi. Bioresour. Technol. 2014, 157, 214–222. [Google Scholar] [CrossRef]
- Brandenburg, J.; Blomqvist, J.; Pickova, J.; Bonturi, N.; Sandgren, M.; Passoth, V. Lipid production from hemicellulose with Lipomyces starkeyi in a pH-regulated fed-batch cultivation. Yeast 2016, 33, 451–462. [Google Scholar] [CrossRef]
- Zhang, S.; Skerker, J.M.; Rutter, C.D.; Maurer, M.J.; Arkin, A.P.; Rao, C.V. Engineering Rhodosporidium toruloides for increased lipid production. Biotechnol. Bioeng. 2015, 113, 1056–1066. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Liu, H.; Zhang, J. Enhanced lipid production with undetoxified corncob hydrolysate by Rhodotorula glutinis using a high cell density culture strategy. Bioresour. Technol. 2015, 180, 32–39. [Google Scholar] [CrossRef]
- Thangavelu, K.; Sundararaju, P.; Srinivasan, N.; Muniraj, I.; Uthandi, S. Simultaneous lipid production for biodiesel feedstock and decontamination of sago processing wastewater using Candida tropicalis ASY2. Biotechnol. Biofuels 2020, 13, 1. [Google Scholar] [CrossRef]
- Guerfali, M.; Ayadi, I.; Sassi, H.E.; Belhassen, A.; Gargouri, A.; Belghith, H. Biodiesel-derived crude glycerol as alternative feedstock for single cell oil production by the oleaginous yeast Candida viswanathii Y-E4. Ind. Crops Prod. 2020, 145, 112103. [Google Scholar] [CrossRef]
- Burgstaller, L.; Oliver, L.; Dietrich, T.; Neureiter, M. Pilot scale production of single cell oil by Apiotrichum brassicae and Pichia kudriavzevii from acetic acid and propionic acid. Appl. Sci. 2023, 13, 4674. [Google Scholar] [CrossRef]
- Wang, G.; Olofsson-Dolk, M.; Hansson, F.G.; Donati, S.; Li, X.; Chang, H.; Cheng, J.; Dahlin, J.; Borodina, I. Engineering Yeast Yarrowia lipolytica for Methanol Assimilation. ACS Synth. Biol. 2021, 10, 3537–3550. [Google Scholar] [CrossRef]
- Nguyen, H.-T.; Pham, T.-T.; Nguyen, P.-T.; Le-Buanec, H.; Rabetafika, H.N.; Razafindralambo, H.L. Advances in Microbial Exopolysaccharides: Present and Future Applications. Biomolecules 2024, 14, 1162. [Google Scholar] [CrossRef]
- Alshawwa, S.Z.; Alshallash, K.S.; Ghareeb, A.; Elazzazy, A.M.; Sharaf, M.; Alharthi, A.; Abdelgawad, F.E.; El-Hossary, D.; Jaremko, M.; Emwas, A.-H.; et al. Assessment of Pharmacological Potential of Novel Exopolysaccharide Isolated from Marine Kocuria sp. Strain AG5: Broad-Spectrum Biological Investigations. Life 2022, 12, 1387. [Google Scholar] [CrossRef]
- Mouro, C.; Gomes, A.P.; Gouveia, I.C. Microbial Exopolysaccharides: Structure, Diversity, Applications, and Future Frontiers in Sustainable Functional Materials. Polysaccharides 2024, 5, 241–287. [Google Scholar] [CrossRef]
- Aloraini, G.S.; Albureikan, M.O.I.; Shahlol, A.M.A.; Shamrani, T.; Daghistani, H.; El-Nablaway, M.; Tharwat, N.A.; Elazzazy, A.M.; Basyony, A.F.; Ghareeb, A. Biomedical and therapeutic potential of marine-derived Pseudomonas sp. strain AHG22 exopolysaccharide: A novel bioactive microbial metabolite. Rev. Adv. Mater. Sci. 2024, 63, 20240016. [Google Scholar] [CrossRef]
- Tan, R.; Lyu, Y.; Zeng, W.; Zhou, J. Enhancing Scleroglucan Production by Sclerotium rolfsii WSH-G01 Through a pH-Shift Strategy Based on Kinetic Analysis. Bioresour. Technol. 2019, 293, 122098. [Google Scholar] [CrossRef]
- Zeng, W.; Wang, J.; Shan, X.; Yu, S.; Zhou, J. Efficient Production of Scleroglucan by Sclerotium rolfsii and Insights into Molecular Weight Modification by High-Pressure Homogenization. Front. Bioeng. Biotechnol. 2021, 9, 748213. [Google Scholar] [CrossRef]
- Sheng, J.; Ling, P.; Wang, F. Constructing a Recombinant Hyaluronic Acid Biosynthesis Operon and Producing Food-Grade Hyaluronic Acid in Lactococcus lactis. J. Ind. Microbiol. Biotechnol. 2015, 42, 197–206. [Google Scholar] [CrossRef]
- Sarnaik, A.; Abernathy, M.H.; Han, X.; Ouyang, Y.; Xia, K.; Chen, Y.; Cress, B.; Zhang, F.; Lali, A.; Pandit, R.; et al. Metabolic Engineering of Cyanobacteria for Photoautotrophic Production of Heparosan, a Pharmaceutical Precursor of Heparin. Algal Res. 2019, 37, 57–63. [Google Scholar] [CrossRef]
- Netrusov, A.I.; Liyaskina, E.V.; Kurgaeva, I.V.; Liyaskina, A.U.; Yang, G.; Revin, V.V. Exopolysaccharides Producing Bacteria: A Review. Microorganisms 2023, 11, 1541. [Google Scholar] [CrossRef]
- Asadpoor, M.; Peeters, C.; Henricks, P.A.J.; Varasteh, S.; Pieters, R.J.; Folkerts, G.; Braber, S. Anti-Pathogenic Functions of Non-Digestible Oligosaccharides in Vitro. Nutrients 2020, 12, 1789. [Google Scholar] [CrossRef]
- Nobre, C.; Alves Filho, E.G.; Fernandes, F.A.N.; Brito, E.S.; Rodrigues, S.; Teixeira, J.A.; Rodrigues, L.R. Production of Fructo-Oligosaccharides by Aspergillus ibericus and Their Chemical Characterization. LWT—Food Sci. Technol. 2018, 89, 58–64. [Google Scholar] [CrossRef]
- Kumar, R.; Næss, G.; Sørensen, M. Xylooligosaccharides from Lignocellulosic Biomass and Their Applications as Nutraceuticals: A Review on Their Production, Purification, and Characterization. J. Sci. Food Agric. 2024, 104, 7765–7775. [Google Scholar] [CrossRef]
- Hashmi, Z.; Idriss, I.M.; Khalid, D.; Abbas, S.H.; Ali, S.O.; Bozdar, M.M.; Usman, T.; Hamid, M.S.; Solangi, N.H. Production of Microorganism-Based Low-Calorie Sugars. In Application of Bio-Additives for the Food Industry; Springer Nature: Cham, Switzerland, 2024; pp. 89–129. [Google Scholar]
- Balakumaran, P.A.; Divakar, K.; Sindhu, R.; Pandey, A.; Binod, P. Gut Microbes: Role in Production of Nutraceuticals. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2022; pp. 273–299. [Google Scholar]
- Lee, K.W.; Kim, G.S.; Baek, A.H.; Hwang, H.S.; Kwon, D.Y.; Kim, S.G.; Lee, S.Y. Isolation and Characterization of Kimchi Starters Leuconostoc mesenteroides PBio03 and Leuconostoc mesenteroides PBio104 for Manufacture of Commercial Kimchi. J. Microbiol. Biotechnol. 2020, 30, 1060–1066. [Google Scholar] [CrossRef]
- Meng, L.; Diao, M.; Wang, Q.; Li, J.; Zhang, Y.; Liu, X.; Zhang, H.; Liu, Q.; Li, Y.; Xie, J. Efficient Biosynthesis of Resveratrol via Combining Phenylalanine and Tyrosine Pathways in Saccharomyces cerevisiae. Microb. Cell Fact. 2023, 22, 46. [Google Scholar] [CrossRef]
- Liu, H.; Song, Y.; Fan, X.; Wang, C.; Lu, X.; Tian, Y. Yarrowia lipolytica as an Oleaginous Platform for the Production of Value-Added Fatty Acid-Based Bioproducts. Front. Microbiol. 2021, 11, 608662. [Google Scholar] [CrossRef]
- Phannarangsee, Y.; Jiawkhangphlu, B.; Thanonkeo, S.; Sornchamrassri, L.; Sommart, T.; Sangthong, K.; Rittirat, K.; Somrith, K. Sorbitol production from mixtures of molasses and sugarcane bagasse hydrolysate using the thermally adapted Zymomonas mobilis ZM AD41. Sci. Rep. 2024, 14, 5563. [Google Scholar] [CrossRef]
- Huang, L.; Wang, W.; Wang, K.; Zhang, L.; Li, X.; Zhang, Y.; Chen, C.; Wang, Z. Protein rational design and modification of erythrose reductase for the improvement of erythritol production in Yarrowia lipolytica. Bioprocess Biosyst. Eng. 2024, 47, 1659–1668. [Google Scholar] [CrossRef]
- Kim, K.; Hou, C.Y.; Choe, D.; Kang, M.; Cho, S.; Sung, B.H.; Lee, D.H.; Lee, S.G.; Kang, T.J.; Cho, B.K. Adaptive Laboratory Evolution of Escherichia coli W Enhances Gamma-Aminobutyric Acid Production Using Glycerol as the Carbon Source. Metab. Eng. 2022, 69, 59–72. [Google Scholar] [CrossRef]
- Vanapalli, K.R.; Bhar, R.; Maity, S.K.; Dubey, B.K.; Kumar, S.; Kumar, V. Life Cycle Assessment of Fermentative Production of Lactic Acid from Bread Waste Based on Process Modelling Using Pinch Technology. Sci. Total Environ. 2023, 905, 167051. [Google Scholar] [CrossRef]
- Schroedter, L.; Schneider, R.; Venus, J. Transforming waste wood into pure L-(+)-lactic acid: Efficient use of mixed sugar media through cell-recycled continuous fermentation. Bioresour. Technol. 2024, 419, 132010. [Google Scholar] [CrossRef]
- Li, C.; Gao, M.; Zhu, W.; Wang, N.; Liu, J.; Zhang, H.; Li, Y.; Zhang, Y. Recent Advances in the Separation and Purification of Lactic Acid from Fermentation Broth. Process Biochem. 2021, 104, 142–151. [Google Scholar] [CrossRef]
- Rawoof, S.A.A.; Kumar, D.; Mani, K.Y.; Devaraj, T.; Subramanian, S. Production of Optically Pure Lactic Acid by Microbial Fermentation: A Review. Environ. Chem. Lett. 2021, 19, 539–556. [Google Scholar] [CrossRef]
- Di Lorenzo, R.; Serra, I.; Porro, D.; Branduardi, P. State of the Art on the Microbial Production of Industrially Relevant Organic Acids. Catalysts 2022, 12, 2. [Google Scholar] [CrossRef]
- El Oirdi, M. Harnessing the Power of Polyphenols: A New Frontier in Disease Prevention and Therapy. Pharmaceuticals 2024, 17, 692. [Google Scholar] [CrossRef]
- Bolat, E.; Sarıtaş, S.; Duman, H.; Eker, F.; Akdaşçi, E.; Karav, S.; Witkowska, A.M. Polyphenols: Secondary Metabolites with a Biological Impression. Nutrients 2024, 16, 2550. [Google Scholar] [CrossRef]
- Yu, W.; Zhang, Y.; Lu, Y.; Liu, Z.; Wang, J.; Li, X.; Zhang, Y.; Chen, M. Recent Research on the Bioactivity of Polyphenols Derived from Edible Fungi and Their Potential in Chronic Disease Prevention. J. Funct. Foods 2025, 124, 106627. [Google Scholar] [CrossRef]
- Arshadi, N.; Nouri, H.; Moghimi, H. Increasing the production of the bioactive compounds in medicinal mushrooms: An omics perspective. Microb. Cell Fact. 2023, 22, 11. [Google Scholar] [CrossRef]
- Mejía-Manzano, L.A.; Ortiz-Alcaráz, C.I.; Parra Daza, L.E.; Suarez Medina, L.; Vargas-Cortez, T.; Fernández-Niño, M.; González Barrios, A.F.; González-Valdez, J. Saccharomyces cerevisiae Biofactory to Produce Naringenin Using a Systems Biology Approach and a Bicistronic Vector Expression Strategy in Flavonoid Production. Microbiol. Spectrum 2024, 12, e0337423. [Google Scholar] [CrossRef]
- Kieliszek, M.; Sandoval Serrano, S.N. The Importance of Selenium in Food Enrichment Processes. J. Trace Elem. Med. Biol. 2023, 79, 127260. [Google Scholar] [CrossRef]
- Zare, H.; Owlia, P.; Vahidi, H.; Ebrahimipour, G.; Gharibi, D.; Shahverdi, A.R. Simultaneous Optimization of the Production of Organic Selenium and Cell Biomass in Saccharomyces cerevisiae by Plackett–Burman and Box–Behnken Design. Iran. J. Pharm. Res. 2018, 17, 1081–1092. [Google Scholar]
- Shelke, A.V.; Kumar, A.; Singh, S.; Sahu, P.K.; Sharma, A.; Patel, A.K. Vitamins as Nutraceuticals for Microbial Infections. In Preventive and Therapeutic Role of Vitamins as Nutraceuticals; Apple Academic Press: New York, NY, USA, 2024; pp. 91–106. [Google Scholar]
- Shi, Y.; Lu, S.; Zhou, X.; Wang, X.; Zhang, C.; Wu, N.; Dong, T.; Xing, S.; Wang, Y.; Xiao, W.; et al. Systematic metabolic engineering enables highly efficient production of vitamin A in Saccharomyces cerevisiae. Synth. Syst. Biotechnol. 2025, 10, 58–67. [Google Scholar] [CrossRef]
- Ledesma-Amaro, R.; Serrano-Amatriain, C.; Jiménez, A.; Revuelta, J.L. Metabolic Engineering of Riboflavin Production in Ashbya gossypii Through Pathway Optimization. Microb. Cell Fact. 2015, 14, 1. [Google Scholar] [CrossRef]
- Bhushan, B.; Kumkum, C.R.; Kumari, M.; Ahire, J.J.; Singh, P.; Kumar, S.; Patel, S. Soymilk Bio-Enrichment by Indigenously Isolated Riboflavin-Producing Strains of Lactobacillus plantarum. LWT 2020, 119, 108871. [Google Scholar] [CrossRef]
- Zhang, Q.; Lyu, S. Microbial interactions in a vitamin C industrial fermentation system: Novel insights and perspectives. Appl. Environ. Microbiol. 2022, 88, e01212-22. [Google Scholar] [CrossRef]
- Zhou, M.; Bi, Y.; Ding, M.; Yuan, Y. One-step biosynthesis of vitamin C in Saccharomyces cerevisiae. Front. Microbiol. 2021, 12, 643472. [Google Scholar] [CrossRef]
- Lee, H.-M.; Vo, P.N.L.; Na, D. Advancement of Metabolic Engineering Assisted by Synthetic Biology. Catalysts 2018, 8, 619. [Google Scholar] [CrossRef]
- Yuan, S.F.; Alper, H.S. Metabolic Engineering of Microbial Cell Factories for Production of Nutraceuticals. Microb. Cell Fact. 2019, 18, 46. [Google Scholar] [CrossRef]
- Cheng, Y.R.; Sun, Z.J.; Cui, G.Z.; Song, X.; Cui, Q. A New Strategy for Strain Improvement of Aurantiochytrium sp. Based on Heavy-Ions Mutagenesis and Synergistic Effects of Cold Stress and Inhibitors of Enoyl-ACP Reductase. Enzym. Microb. Technol. 2016, 93–94, 182–190. [Google Scholar] [CrossRef]
- Yuan, X.; Zhong, M.; Huang, X.; Hussain, Z.; Ren, M.; Xie, X. Industrial Production of Functional Foods for Human Health and Sustainability. Foods 2024, 13, 3546. [Google Scholar] [CrossRef]
- Ma, D.; Du, G.; Fang, H.; Zhang, J.; Liu, Y.; Li, X.; Li, W.; Wang, Z. Advances and prospects in microbial production of biotin. Microb. Cell Fact. 2024, 23, 135. [Google Scholar] [CrossRef]
- Wei, P.P.; Zhu, F.C.; Chen, C.W.; Li, G.S. Engineering a heterologous synthetic pathway in Escherichia coli for efficient production of biotin. Biotechnol. Lett. 2021, 43, 1221–1228. [Google Scholar] [CrossRef]
- Miranda, B.C.J.; Tustumi, F.; Nakamura, E.T.; Shimanoe, V.H.; Kikawa, D.; Waisberg, J. Obesity and Colorectal Cancer: A Narrative Review. Medicina 2024, 60, 1218. [Google Scholar] [CrossRef]
- Martinez, F.G.; Moreno-Martin, G.; Pescuma, M.; Madrid-Albarrán, Y.; Mozzi, F. Biotransformation of Selenium by Lactic Acid Bacteria: Formation of Seleno-Nanoparticles and Seleno-Amino Acids. Front. Bioeng. Biotechnol. 2020, 8, 17. [Google Scholar] [CrossRef]
- Mahara, F.A.; Nuraida, L.; Lioe, H.N. Fermentation of Milk Using Folate-Producing Lactic Acid Bacteria to Increase Natural Folate Content: A Review. J. Appl. Biotechnol. Rep. 2019, 6, 129–136. [Google Scholar] [CrossRef]
- Idrees, M.; Imran, M.; Atiq, N.; Zahra, R.; Abid, R.; Alreshidi, M.; Roberts, T.; Abdelgadir, A.; Tipu, M.K.; Farid, A.; et al. Probiotics, their action modality and the use of multi-omics in metamorphosis of commensal microbiota into target-based probiotics. Front. Nutr. 2022, 9, 959941. [Google Scholar] [CrossRef]
- Maeda, H.; Sakuragi, Y.; Bryant, D.A.; Dellapenna, D. Tocopherols protect Synechocystis sp. strain PCC 6803 from lipid peroxidation. Plant Physiol. 2005, 138, 1422–1435. [Google Scholar] [CrossRef]
- Ye, Z.; Shi, B.; Huang, Y.; Ma, T.; Xiang, Z.; Hu, B.; Kuang, Z.; Huang, M.; Lin, X.; Tian, Z.; et al. Revolution of vitamin E production by starting from microbial fermented farnesene to isophytol. Innovation 2022, 3, 100228. [Google Scholar] [CrossRef]
- Bi, H.; Xv, C.; Su, C.; Feng, P.; Zhang, C.; Wang, M.; Fang, Y.; Tan, T. β-Farnesene Production from Low-Cost Glucose in Lignocellulosic Hydrolysate by Engineered Yarrowia lipolytica. Fermentation 2022, 8, 532. [Google Scholar] [CrossRef]
- Liu, Y.; Van Bennekom, E.O.; Zhang, Y.; Abee, T.; Smid, E.J. Long-Chain Vitamin K2 Production in Lactococcus lactis Is Influenced by Temperature, Carbon Source, Aeration, and Mode of Energy Metabolism. Microb. Cell Fact. 2019, 18, 129. [Google Scholar] [CrossRef]
- Staudinger, J.L.; Mahroke, A.; Patel, G.; Dattel, C.; Reddy, S. Pregnane X receptor signaling pathway and vitamin K: Molecular mechanisms and clinical relevance in human health. Cells 2024, 13, 681. [Google Scholar] [CrossRef]
- Jach, M.E.; Malm, A. Yarrowia lipolytica as an alternative and valuable source of nutritional and bioactive compounds for humans. Molecules 2022, 27, 2300. [Google Scholar] [CrossRef]
- Tomaszewska-Hetman, L.; Rywińska, A.; Lazar, Z.; Rymowicz, W. Enhancement of α-Ketoglutaric Acid Production by Yarrowia lipolytica Grown on Mixed Renewable Carbon Sources through Adjustment of Culture Conditions. Catalysts 2023, 13, 14. [Google Scholar] [CrossRef]
- Icer, M.A.; Sarikaya, B.; Kocyigit, E.; Atabilen, B.; Çelik, M.N.; Capasso, R.; Ağagündüz, D.; Budán, F. Contributions of Gamma-Aminobutyric Acid (GABA) Produced by Lactic Acid Bacteria on Food Quality and Human Health: Current Applications and Future Prospects. Foods 2024, 13, 2437. [Google Scholar] [CrossRef]
- Han, J.; Zhao, X.; Zhao, X.; Wang, Q.; Li, P.; Gu, Q. Microbial-Derived γ-Aminobutyric Acid: Synthesis, Purification, Physiological Function, and Applications. J. Agric. Food Chem. 2023, 71, 14931–14946. [Google Scholar] [CrossRef]
- Milon, R.B.; Hu, P.; Zhang, X.; Hu, X.; Ren, L. Recent Advances in the Biosynthesis and Industrial Biotechnology of Gamma-Amino Butyric Acid. Bioresour. Bioprocess. 2024, 11, 32. [Google Scholar] [CrossRef]
- Abarquero, D.; Bodelón, R.; Flórez, A.B.; Fresno, J.M.; Renes, E.; Mayo, B.; Tornadijo, M.E. Technological and Safety Assessment of Selected Lactic Acid Bacteria for Cheese Starter Cultures Design: Enzymatic and Antimicrobial Activity, Antibiotic Resistance and Biogenic Amine Production. LWT 2023, 180, 114709. [Google Scholar] [CrossRef]
- Kiki, M.J. Biopigments of Microbial Origin and Their Application in the Cosmetic Industry. Cosmetics 2023, 10, 47. [Google Scholar] [CrossRef]
- Loh, W.L.C.; Huang, K.C.; Ng, H.S.; Lan, J.C.W. Exploring the Fermentation Characteristics of a Newly Isolated Marine Bacteria Strain, Gordonia terrae TWRH01 for Carotenoids Production. J. Biosci. Bioeng. 2020, 130, 187–194. [Google Scholar] [CrossRef]
- Anshi; Kapil, S.; Goswami, L.; Sharma, V. Unveiling the Intricacies of Microbial Pigments as Sustainable Alternatives to Synthetic Colorants: Recent Trends and Advancements. Micro 2024, 4, 621–640. [Google Scholar] [CrossRef]
- Venil, C.K.; Dufossé, L.; Renuka Devi, P. Bacterial Pigments: Sustainable Compounds with Market Potential for Pharma and Food Industry. Front. Sustain. Food Syst. 2020, 4, 100. [Google Scholar] [CrossRef]
- Bidram, M.; Ganjalikhany, M.R. Bioactive Peptides from Food Science to Pharmaceutical Industries: Their Mechanism of Action, Potential Role in Cancer Treatment and Available Resources. Heliyon 2024, 10, e40563. [Google Scholar] [CrossRef]
- Peres Fabbri, L.; Cavallero, A.; Vidotto, F.; Gabriele, M. Bioactive Peptides from Fermented Foods: Production Approaches, Sources, and Potential Health Benefits. Foods 2024, 13, 3369. [Google Scholar] [CrossRef]
- Cruz-Casas, D.E.; Aguilar, C.N.; Ascacio-Valdés, J.A.; Rodríguez-Herrera, R.; Chávez-González, M.L.; Flores-Gallegos, A.C. Enzymatic Hydrolysis and Microbial Fermentation: The Most Favorable Biotechnological Methods for the Release of Bioactive Peptides. Food Chem. Mol. Sci. 2021, 3, 100047. [Google Scholar] [CrossRef]
- Purohit, K.; Reddy, N.; Sunna, A. Exploring the Potential of Bioactive Peptides: From Natural Sources to Therapeutics. Int. J. Mol. Sci. 2024, 25, 1391. [Google Scholar] [CrossRef]
- Bommakanti, V.; Puthenparambil Ajikumar, A.; Sivi, C.M.; Prakash, G.; Mundanat, A.S.; Ahmad, F.; Haque, S.; Prieto, M.A.; Rana, S.S. An Overview of Herbal Nutraceuticals, Their Extraction, Formulation, Therapeutic Effects and Potential Toxicity. Separations 2023, 10, 177. [Google Scholar] [CrossRef]
- Chaudhary, R.; Nawaz, A.; Fouillaud, M.; Dufossé, L.; Haq, I.u.; Mukhtar, H. Microbial Cell Factories: Biodiversity, Pathway Construction, Robustness, and Industrial Applicability. Microbiol. Res. 2024, 15, 247–272. [Google Scholar] [CrossRef]
- Gast, V.; Sandegren, A.; Dunås, F.; Eriksson, A.; Berntsson, O.; Wernersson, K.; Rosenqvist, L. Engineering Saccharomyces cerevisiae for the Production and Secretion of Affibody Molecules. Microb. Cell Fact. 2022, 21, 36. [Google Scholar] [CrossRef]
- Goold, H.D.; Moseley, J.L.; Lauersen, K.J. The synthetic future of algal genomes. Cell Genom. 2024, 4, 100505. [Google Scholar] [CrossRef]
- Henry, L.P.; Bruijning, M.; Forsberg, S.K.; Ayroles, J.F. The Microbiome Extends Host Evolutionary Potential. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Chen, L.; Li, Y.; Han, P.; Zhang, Y.; Xu, H.; Wei, X.; Zhong, W.; Li, J.; Guo, X.; Song, Y.; et al. Integrative Multiomics Analysis Reveals Host-Microbe-Metabolite Interplays Associated with the Aging Process in Singaporeans. Gut Microbes 2022, 14, 2070392. [Google Scholar] [CrossRef]
- Salvetti, E.; O’Toole, P.W. The Genomic Basis of Lactobacilli as Health-Promoting Organisms. Microbiol. Spectr. 2017, 5, 22. [Google Scholar] [CrossRef]
- Gueimonde, M.; Collado, M. Metagenomics and Probiotics. Clin. Microbiol. Infect. 2012, 18, 32–34. [Google Scholar] [CrossRef]
- Bottacini, F.; van Sinderen, D.; Ventura, M. Omics of Bifidobacteria: Research and Insights into Their Health-Promoting Activities. Biochem. J. 2017, 474, 4137–4152. [Google Scholar] [CrossRef]
- Kiousi, D.E.; Rathosi, M.; Tsifintaris, M.; Chondrou, P.; Galanis, A. Probiomics: Omics Technologies to Unravel the Role of Probiotics in Health and Disease. Adv. Nutr. 2021, 12, 1802–1820. [Google Scholar] [CrossRef]
- Ngashangva, N.; Mukherjee, P.K.; Sharma, C.; Kalita, M.C.; Sarangthem, I. Integrated Genomics and Proteomics Analysis of Paenibacillus peoriae IBSD35 and Insights into Its Antimicrobial Characteristics. Sci. Rep. 2022, 12, 18861. [Google Scholar] [CrossRef]
- Perkins, D.N.; Pappin, D.J.; Creasy, D.M.; Cottrell, J.S. Probability-Based Protein Identification by Searching Sequence Databases Using Mass Spectrometry Data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, K.; Hendrie, C.; Liang, C.; Li, M.; Doherty-Kirby, A.; Lajoie, G. PEAKS: Powerful Software for Peptide De Novo Sequencing by Tandem Mass Spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 2337–2342. [Google Scholar] [CrossRef]
- Kwoji, I.D.; Aiyegoro, O.A.; Okpeku, M.; Adeleke, M.A. ‘Multi-Omics’ Data Integration: Applications in Probiotics Studies. NPJ Sci. Food 2023, 7, 25. [Google Scholar] [CrossRef]
- Chemla, Y.; Sweeney, C.J.; Wozniak, C.A.; Patel, S.; Lee, R.; Roberts, S.; Johnson, M.; Cohen, P.; Zhang, X.; Tang, Y. Design and Regulation of Engineered Bacteria for Environmental Release. Nat. Microbiol. 2025, 10, 281–300. [Google Scholar] [CrossRef]
- EFSA Panel on Genetically Modified Organisms (GMO); Georgieva, V.V.; Mariani, M.L.; Vervent, M.S.; Bolognese, F.; Gerber, A.; Danilevich, D.; Jenkins, R.; Nowak, K.; Angelov, I. New Developments in Biotechnology Applied to Microorganisms. EFSA J. 2024, 22, e8895. [Google Scholar]
- Hanlon, P.; Sewalt, V. GEMs: Genetically Engineered Microorganisms and the Regulatory Oversight of Their Uses in Modern Food Production. Crit. Rev. Food Sci. Nutr. 2020, 61, 959–970. [Google Scholar] [CrossRef]
- Wesseler, J.; Scatasta, S.; Banse, M.; Benning, S.; Bessou, C.; Blanchet, C.; Goh, Y.; Hansen, B.; Laube, W.; Morel, J. EU Regulation of Genetically Modified Microorganisms in Light of New Policy Developments: Possible Implications for EU Bioeconomy Investments. Appl. Econ. Perspect. Policy 2023, 45, 839–859. [Google Scholar] [CrossRef]
- Santos-Beneit, F. What is the role of microbial biotechnology and genetic engineering in medicine? MicrobiologyOpen 2024, 13, e1406. [Google Scholar] [CrossRef]
- Park, J.Y.; Lim, J.H.; Ahn, J.H.; Kim, Y.; Lee, S.; Jeong, H.; Park, S.; Kang, J. Biosynthesis of Resveratrol Using Metabolically Engineered Escherichia coli. Appl. Biol. Chem. 2021, 64, 20. [Google Scholar] [CrossRef]
- Park, Y.-K.; Nicaud, J.-M. Metabolic Engineering for Unusual Lipid Production in Yarrowia lipolytica. Microorganisms 2020, 8, 1937. [Google Scholar] [CrossRef]
- Paddon, C.J.; Westfall, P.J.; Pitera, D.J.; Benjamin, K.; Fisher, K.; McPhee, J.; Harker, M.; Palsson, B.Ø.; Keasling, J.D. High-Level Semi-Synthetic Production of the Potent Antimalarial Artemisinin. Nature 2013, 496, 528–532. [Google Scholar] [CrossRef]
- Mendes, A.R.; Spínola, M.P.; Lordelo, M.; Prates, J.A.M. Advances in Bioprocess Engineering for Optimising Chlorella vulgaris Fermentation: Biotechnological Innovations and Applications. Foods 2024, 13, 4154. [Google Scholar] [CrossRef]
- Rainha, J.; Gomes, D.; Rodrigues, L.R.; Rodrigues, J.L. Synthetic Biology Approaches to Engineer Saccharomyces cerevisiae Towards the Industrial Production of Valuable Polyphenolic Compounds. Life 2020, 10, 56. [Google Scholar] [CrossRef]
- Meng, G.; Zhao, W.; Liu, Q.; Li, X.; Zhang, H.; Liu, W.; Wang, S.; Zhang, Y.; Yang, X. Strategy for the Design of a Bioreactor for L-Lysine Immobilized Fermentation Using Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2022, 106, 5449–5458, Erratum in: Appl. Microbiol. Biotechnol. 2023, 107, 4987. [Google Scholar] [CrossRef]
- Khattab, S.M.R.; Watanabe, T. Efficient Conversion of Glycerol to Ethanol by an Engineered Saccharomyces cerevisiae Strain. Appl. Environ. Microbiol. 2021, 87, e00268-21. [Google Scholar] [CrossRef]
- Chotirotsukon, C.; Khattab, S.M.R.; Kobayashi, N.; Katahira, M.; Laosiripojana, N.; Champreda, V.; Watanabe, T. Microwave-Accelerated Glycerolysis of Sugarcane Trash Using Lewis Acid, AlK(SO4)2, for Bioethanol Production. Ind. Crops Prod. 2022, 190, 115849. [Google Scholar] [CrossRef]
- Khattab, S.M.R.; Okano, H.; Kimura, C.; Fujita, T.; Watanabe, T. Efficient Integrated Production of Bioethanol and Antiviral Glycerolysis Lignin from Sugarcane Trash. Biotechnol. Biofuels 2023, 16, 82. [Google Scholar] [CrossRef]
- Khattab, S.M.R.; Abdel-Rahman, M.A.; Katahira, M.; Watanabe, T. Engineering Saccharomyces cerevisiae and Controlling Conditions for 2,3-Butanediol Production from Glycerol. Sustain. Energy Fuels 2024, 8, 4297–4310. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Lunina, J.N. The Citric Acid Production from Raw Glycerol by Yarrowia lipolytica Yeast and Its Regulation. Appl. Microbiol. Biotechnol. 2013, 97, 7387–7397. [Google Scholar] [CrossRef]
- Yang, S.; Pan, X.; Wang, Q.; Liu, Y.; Zhang, J.; Li, W.; Zhang, Y.; Chen, L.; Li, Z. Enhancing Erythritol Production from Crude Glycerol in a Wild-Type Yarrowia lipolytica by Metabolic Engineering. Front. Microbiol. 2022, 13, 1054243. [Google Scholar] [CrossRef]
- Mavrommati, M.; Papanikolaou, S.; Aggelis, G. Improving Ethanol Tolerance of Saccharomyces cerevisiae Through Adaptive Laboratory Evolution Using High Ethanol Concentrations as a Selective Pressure. Process Biochem. 2023, 124, 280–289. [Google Scholar] [CrossRef]
- Dragosits, M.; Mattanovich, D. Adaptive Laboratory Evolution—Principles and Applications for Biotechnology. Microb. Cell Fact. 2013, 12, 64. [Google Scholar] [CrossRef]
- Malvido, M.C.; González, E.A.; Bazán Tantaleán, D.L.; Bendaña Jácome, R.J.; Guerra, N.P. Batch and Fed-Batch Production of Probiotic Biomass and Nisin in Nutrient-Supplemented Whey Media. Braz. J. Microbiol. 2019, 50, 915–925. [Google Scholar] [CrossRef]
- Li, Y.P.; Ahmadi, F.; Kariman, K.; Zhang, X.; Wang, Y.; Li, W.; Zhang, L.; Zhao, S.; Chen, X. Recent advances and challenges in single cell protein (SCP) technologies for food and feed production. NPJ Sci Food 2024, 8, 66. [Google Scholar] [CrossRef]
- Naveira-Pazos, C.; Veiga, M.C.; Mussagy, C.U.; Farias, F.O.; Kennes, C.; Pereira, J.F.B. Carotenoids production and extraction from Yarrowia lipolytica cells: A biocompatible approach using biosolvents. Sep. Purif. Technol. 2024, 343, 127136. [Google Scholar] [CrossRef]
- Ravi, G.; Venkata Dasu, V.; Pakshirajan, K. A comparative evaluation of batch and fed-batch cultures for enhanced lipid, carotenoid, and β-carotene production by Rhodotorula mucilaginosa. Prep. Biochem. Biotechnol. 2024, 1–14, Online ahead of print. [Google Scholar] [CrossRef]
- López, J.; Cataldo, V.F.; Peña, M.; Saa, P.A.; Saitua, F.; Ibaceta, M.; Agosin, E. Build Your Bioprocess on a Solid Strain—β-Carotene Production in Recombinant Saccharomyces cerevisiae. Front. Bioeng. Biotechnol. 2019, 7, 171. [Google Scholar] [CrossRef]
- Abouelela, M.E.; Helmy, Y.A. Next-generation probiotics as novel therapeutics for improving human health: Current trends and future perspectives. Microorganisms 2024, 12, 430. [Google Scholar] [CrossRef]
- Punia, S.; Sandhu, K.S.; Grasso, S.; Singh Purewal, S.; Kaur, M.; Kumar Siroha, A.; Kumar, K.; Kumar, V.; Kumar, M. Aspergillus oryzae Fermented Rice Bran: A Byproduct with Enhanced Bioactive Compounds and Antioxidant Potential. Foods 2020, 10, 70. [Google Scholar] [CrossRef]
- Cui, S.; Sadiq, F.A.; Mao, B.; Xu, Z.; Liu, Z.; Zhao, J.; Zhang, H.; Chen, W. High-Density Cultivation of Lactobacillus and Bifidobacterium Using an Automatic Feedback Feeding Method. LWT 2019, 112, 108232. [Google Scholar] [CrossRef]
- Wan, S.; Liu, X.; Sun, W.; Zhang, Y.; Li, Q.; Wang, L.; Xu, X. Current Advances for Omics-Guided Process Optimization of Microbial Manufacturing. Bioresour. Bioprocess. 2023, 10, 30. [Google Scholar] [CrossRef]
- Dzurendova, S.; Olsen, P.M.; Byrtusová, D.; Nováková, M.; Li, H.; Zhang, Y.; Wang, Z.; Li, T. Raman Spectroscopy Online Monitoring of Biomass Production, Intracellular Metabolites and Carbon Substrates During Submerged Fermentation of Oleaginous and Carotenogenic Microorganisms. Microb. Cell Fact. 2023, 22, 261. [Google Scholar] [CrossRef]
- Sahin, D.; Tas, E.; Altindag, U.H. Enhancement of Docosahexaenoic Acid (DHA) Production from Schizochytrium sp. S31 Using Different Growth Medium Conditions. AMB Express 2018, 8, 7. [Google Scholar] [CrossRef]
- Badreldin, N.; Cheng, X.; Youssef, A. An Overview of Software Sensor Applications in Biosystem Monitoring and Control. Sensors 2024, 24, 6738. [Google Scholar] [CrossRef]
- Eifert Antosz, K.; Knapčíková, L.; Husár, J. Evaluation and Application of Machine Learning Techniques for Quality Improvement in Metal Product Manufacturing. Appl. Sci. 2024, 14, 10450. [Google Scholar] [CrossRef]
- Gargalo, C.L.; Udugama, I.; Pontius, K.; Palsson, B.O.; Mendes, P.; Rother, M.; De Mey, M. Towards Smart Biomanufacturing: A Perspective on Recent Developments in Industrial Measurement and Monitoring Technologies for Bio-Based Production Processes. J. Ind. Microbiol. Biotechnol. 2020, 47, 947–964. [Google Scholar] [CrossRef]
- Refai, M.Y.; Elazzazy, A.M.; Abu-Elghait, M.; El-Refai, M.; Abdel-Raouf, U.; El-Sayed, H.; Farag, M.A. Ziziphus spina-christi (L.) Willd. Leaves Extract Affecting agr Quorum Sensing System in Staphylococcus aureus. Arab. J. Sci. Eng. 2024, 49, 97–105. [Google Scholar] [CrossRef]
- Wu, S.; Liu, J.; Liu, C.; Xiao, M.; Zeng, W.; Tan, Z.; Jiang, Y.; Zhang, Y.; Zhang, T. Quorum Sensing for Population-Level Control of Bacteria and Potential Therapeutic Applications. Cell. Mol. Life Sci. 2020, 77, 1319–1343. [Google Scholar] [CrossRef]
- Bashor, C.J.; Patel, N.; Choubey, S.; Beyzavi, A.; Kondev, J.; Collins, J.J.; Khalil, A.S. Complex Signal Processing in Synthetic Gene Circuits Using Cooperative Regulatory Assemblies. Science 2019, 364, 593–597. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, J.; Kalimuthu, S.; Li, Z.; Yin, L.; Zhao, M.; Xu, C. A systematically biosynthetic investigation of lactic acid bacteria reveals diverse antagonistic bacteriocins that potentially shape the human microbiome. Microbiome 2023, 11, 91. [Google Scholar] [CrossRef]
- Refai, M.Y.; Elazzazy, A.M.; Desouky, S.E.; Al-Hindi, R.; Abdel-Raouf, U.; El-Sayed, H. Interception of Epoxide Ring to Quorum Sensing System in Enterococcus faecalis and Staphylococcus aureus. AMB Express 2023, 13, 126. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.; Wang, J.; Chen, Y.; Liu, L.; Zhao, Q. Quorum Sensing of Bifidobacteria: Research and Progress. Microbiol. Res. 2025, 294, 128102. [Google Scholar] [CrossRef]
- Li, Y.; Sun, W.; Tang, H.; Zhou, X.; Zhang, M. Metabolomics-Based Insights into the Mechanism of Quorum Sensing-Enhanced Conjugated Linoleic Acid Production in Limosilactobacillus fermentum L1. LWT 2025, 218, 117477. [Google Scholar] [CrossRef]
- Du, R.; Feng, L.; Yang, Y.; Li, Z.; Huang, C.; Chen, T. Enhanced Dextran Production by Weissella confusa in Co-Culture with Candida shehatae and Its Quorum Sensing Regulation Mechanism. Int. J. Biol. Macromol. 2025, 295, 139662. [Google Scholar] [CrossRef]
- Muniz, E.d.N.; Montenegro, R.T.d.Q.; da Silva, D.N.; D’Almeida, A.P.; Gonçalves, L.R.B.; de Albuquerque, T.L. Advances in Biotechnological Strategies for Sustainable Production of Non-Animal Proteins: Challenges, Innovations, and Applications. Fermentation 2024, 10, 638. [Google Scholar] [CrossRef]
- Pashang, R.; Gilbride, K.A.; Wenk, J. Microbial Dynamics and Quality Monitoring in Biopharmaceutical Production. CBEN 2024, 11, e202400022. [Google Scholar] [CrossRef]
- Wang, S.; Ju, D.; Zeng, X. Mechanisms and Clinical Implications of Human Gut Microbiota-Drug Interactions in the Precision Medicine Era. Biomedicines 2024, 12, 194. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism Type | Species | Biomass (g/L) | Lipid Content (%) | Lipid Titer (g/L) | Overall Yield (g/g) | Lipid Productivity (g/L/h) | Feedstock/Substrate | Lipid Composition Majority | Culture Mode | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Yeast | Yarrowia lipolytica NS432 | 115.75 | 73 | 84.5 | 0.2 | 0.73 | Glucose | (C16:0), (C16:1), (C18:0), (C18:1), and (C18:2) | Fed-batch bioreactor | [39] |
| 40 | 77 | 30.8 | 0.21 | 0.27 | Batch bioreactor | |||||
| Yarrowia lipolytica | 89.3 | 81.4 | 72.7 | 0.252 | 0.97 | Glucose | (C18:1) 50% and (C18:2) 10% | Stepwise exponential feeding bioreactor | [40] | |
| Cryptococcus curvatus | 4.32 | 41.2 | 1.78 | 0.11 | 0.037 | Waste office paper | (C18:1) almost around 50%, (C16:0), (C18:0), and (C18:2) | Batch shake flask | [41] | |
| Lipomyces starkeyi | 85.4 | 49 | 41.8 | 0.1 | 0.176 | Xylose, glucose | (C14:0), (C16:0), (C18:0), (C18:1), (C18:2), and (C18:3) | Repeated fed-batch bioreactor | [42] | |
| Lipomyces starkeyi | 13.30 | 60.47 | 8.04 | 0.11 | 0.0558 | Hemicellulose hydrolysate | (C16:0), (C18:1), (C18:0), (C18:2), and (C18:3) | pH-regulated fed-batch cultivation, bioreactor | [43] | |
| Rhodosporidium toruloides | 26.8 | 61.1 | 16.4 | 0.23 | 0.08 | Glucose | (C18:1) and (C16:0) | Batch shake flask | [44] | |
| Rhodotorula glutinis | 70.8 | 47.2 | 33.5 | 0.159 | 0.17 | Undetoxified corncob hydrolysate | (C16:0), (C18:1), and (C18:2) | Fed-batch culture with two-stage nitrogen feeding strategy in bioreactor | [45] | |
| Candida tropicalis ASY2 | 2.49 | 48.59 | 1.21 | 0.135 | 0.01 | Starch Wastewater (SWW) | (C18:1) and (C20:1) | Batch-flask | [46] | |
| Candida viswanathii Y-E4 | 26.2 | 51.9 | 13.6 | 13.6 | 0.08 | Crude glycerol | (C18:1) and (C18:2) | Batch-flask | [47] | |
| Pichia kudriavzevii | 33.2 | 57.9 | 19.2 | 0.162 | 0.296 | Acetic and propionic acid | (C16:0), (C18:0), (C18:2), and (C17:0) | Fed-batch bioreactor | [48] |
| Exopolysaccharide | Microbial Source | Key Biosynthetic Enzymes | Industrial Applications | Yield (g/L) | Bioactivity and Mechanism | Reference |
|---|---|---|---|---|---|---|
| Xanthan | Xanthomonas campestris | UDP-glucose pyrophosphorylase, glycosyltransferases | Food, cosmetics, biomedicine | 50+ | Antioxidant (ROS scavenging, Nrf2 activation) | [50] |
| Hyaluronic Acid | Streptococcus zooepidemicus | Hyaluronan synthase | Pharmaceuticals, skincare, tissue engineering | 15 | Immunomodulation (TLR2, cytokine activation) | [56] |
| Alginate | Pseudomonas aeruginosa | Alginate polymerase | Food, wound healing, biofilms | 30 | Prebiotic (SCFA production, gut microbiota) | [51] |
| Levan | Bacillus subtilis | Levansucrase | Prebiotics, drug delivery | 40 | Antimicrobial (biofilm inhibition) | [50] |
| Scleroglucan | Sclerotium rolfsii | Beta-glucan synthases | Pharmaceuticals, antiviral agents | 66.6 | Immunostimulatory (TLR4 activation, TNF-α secretion) | [55] |
| Bioactive Peptide | Microbial Source | Bioactive Mechanism | Industrial Applications | Reference |
|---|---|---|---|---|
| Lactotripeptides (VPP, IPP) | Lactobacillus helveticus | ACE inhibition (antihypertensive) | Functional dairy, cardiovascular health | [115] |
| Casein-derived peptides | Lactobacillus casei | Antioxidant (ROS scavenging, Nrf2 activation) | Nutraceuticals, anti-aging formulations | [116] |
| Soy-derived peptides | Bifidobacterium longum | Immunomodulation (TLR2-mediated cytokine activation) | Functional foods, gut health | [117] |
| Antimicrobial peptides (AMPs) | Bacillus subtilis | Membrane disruption, quorum sensing inhibition | Food preservatives, antibiotics | [118] |
| Lactotripeptides (VPP, IPP) | Lactobacillus helveticus | ACE inhibition (antihypertensive) | Functional dairy, cardiovascular health | [115] |
| Casein-derived peptides | Lactobacillus casei | Antioxidant (ROS scavenging, Nrf2 activation) | Nutraceuticals, anti-aging formulations | [116] |
| Soy-derived peptides | Bifidobacterium longum | Immunomodulation (TLR2-mediated cytokine activation) | Functional foods, gut health | [117] |
| Antimicrobial peptides (AMPs) | Bacillus subtilis | Membrane disruption, quorum sensing inhibition | Food preservatives, antibiotics | [118] |
| Criteria | Traditional Production (Plant/Animal-Based) | Microbial Production (Fermentation-Based) | Effectiveness (Microbial vs. Traditional) |
|---|---|---|---|
| Source | Extracted from plants or animal tissues (e.g., bovine cartilage, fish collagen, plant-derived polysaccharides, chemically synthesized lactic acid) | Produced via microbial fermentation (Lactobacillus, Bacillus, Streptococcus, Xanthomonas, Sclerotium rolfsii) | Microbial production is more sustainable and scalable, avoiding seasonal/raw material fluctuations |
| Yield | Variable, seasonal limitations (e.g., 0.2–2 g/L for plant-extracted polysaccharides, 0.5–1 g/L for lactic acid from petrochemical sources) | High, stable, and scalable (e.g., 50–80 g/L for microbial EPS, 10–20 g/L for bioactive peptides, 66.6 g/L for scleroglucan, 100–150 g/L for lactic acid fermentation) | Microbial yields are significantly higher (10–50× improvement), making production more efficient |
| Purification Process | High cost, low efficiency—contamination risk, batch-to-batch variation, difficult to remove unwanted plant/animal residues, high refining cost for petrochemical lactic acid | Lower cost, higher efficiency—controlled microbial expression reduces unwanted byproducts, enabling cost-effective downstream processing | Microbial production is more cost-effective due to controlled biosynthesis, reducing unwanted impurities |
| Purification Cost and Challenges | High cost, low efficiency—contamination risk, batch-to-batch variation, difficult to remove unwanted plant/animal residues, high refining cost for petrochemical lactic acid | Lower cost, higher efficiency—controlled microbial expression reduces unwanted byproducts, enabling cost-effective downstream processing | Microbial production is more cost-effective due to controlled biosynthesis, reducing unwanted impurities |
| Bioactivity Retention | Can degrade due to processing conditions (e.g., heat, solvent exposure) | Controlled fermentation preserves bioactivity, optimizing peptide/enzyme activity | Microbial processes offer higher bioactivity retention |
| Purity and Consistency | Variable purity—batch-to-batch variation, presence of contaminants (e.g., heavy metals, allergens, chemical residues in lactic acid synthesis) | Higher purity and consistency—controlled biosynthesis minimizes contaminants | Microbial systems provide higher reproducibility and purity |
| Sustainability | High land, water, and energy consumption, reliance on petrochemical sources (lactic acid) | Lower environmental footprint, sustainable production | Microbial processes are greener, reducing resource use |
| Cost | Dependent on agricultural fluctuations, higher raw material costs, chemical synthesis cost for lactic acid | Lower long-term cost after process optimization | Microbial production is more cost-effective long-term |
| Industrial Examples |
|
| Microbial production dominates high-value functional ingredients, especially for scleroglucan, lactic acid, xanthan, and bioactive peptides |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elazzazy, A.M.; Baeshen, M.N.; Alasmi, K.M.; Alqurashi, S.I.; Desouky, S.E.; Khattab, S.M.R. Where Biology Meets Engineering: Scaling Up Microbial Nutraceuticals to Bridge Nutrition, Therapeutics, and Global Impact. Microorganisms 2025, 13, 566. https://doi.org/10.3390/microorganisms13030566
Elazzazy AM, Baeshen MN, Alasmi KM, Alqurashi SI, Desouky SE, Khattab SMR. Where Biology Meets Engineering: Scaling Up Microbial Nutraceuticals to Bridge Nutrition, Therapeutics, and Global Impact. Microorganisms. 2025; 13(3):566. https://doi.org/10.3390/microorganisms13030566
Chicago/Turabian StyleElazzazy, Ahmed M., Mohammed N. Baeshen, Khalid M. Alasmi, Shatha I. Alqurashi, Said E. Desouky, and Sadat M. R. Khattab. 2025. "Where Biology Meets Engineering: Scaling Up Microbial Nutraceuticals to Bridge Nutrition, Therapeutics, and Global Impact" Microorganisms 13, no. 3: 566. https://doi.org/10.3390/microorganisms13030566
APA StyleElazzazy, A. M., Baeshen, M. N., Alasmi, K. M., Alqurashi, S. I., Desouky, S. E., & Khattab, S. M. R. (2025). Where Biology Meets Engineering: Scaling Up Microbial Nutraceuticals to Bridge Nutrition, Therapeutics, and Global Impact. Microorganisms, 13(3), 566. https://doi.org/10.3390/microorganisms13030566