
First Report of Phytophthora mengei Causing Root Rot and Canker in Avocado (Persea americana) in Michoacán, Mexico

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation
2.2. Morphological Characterization
2.3. Molecular Characterization
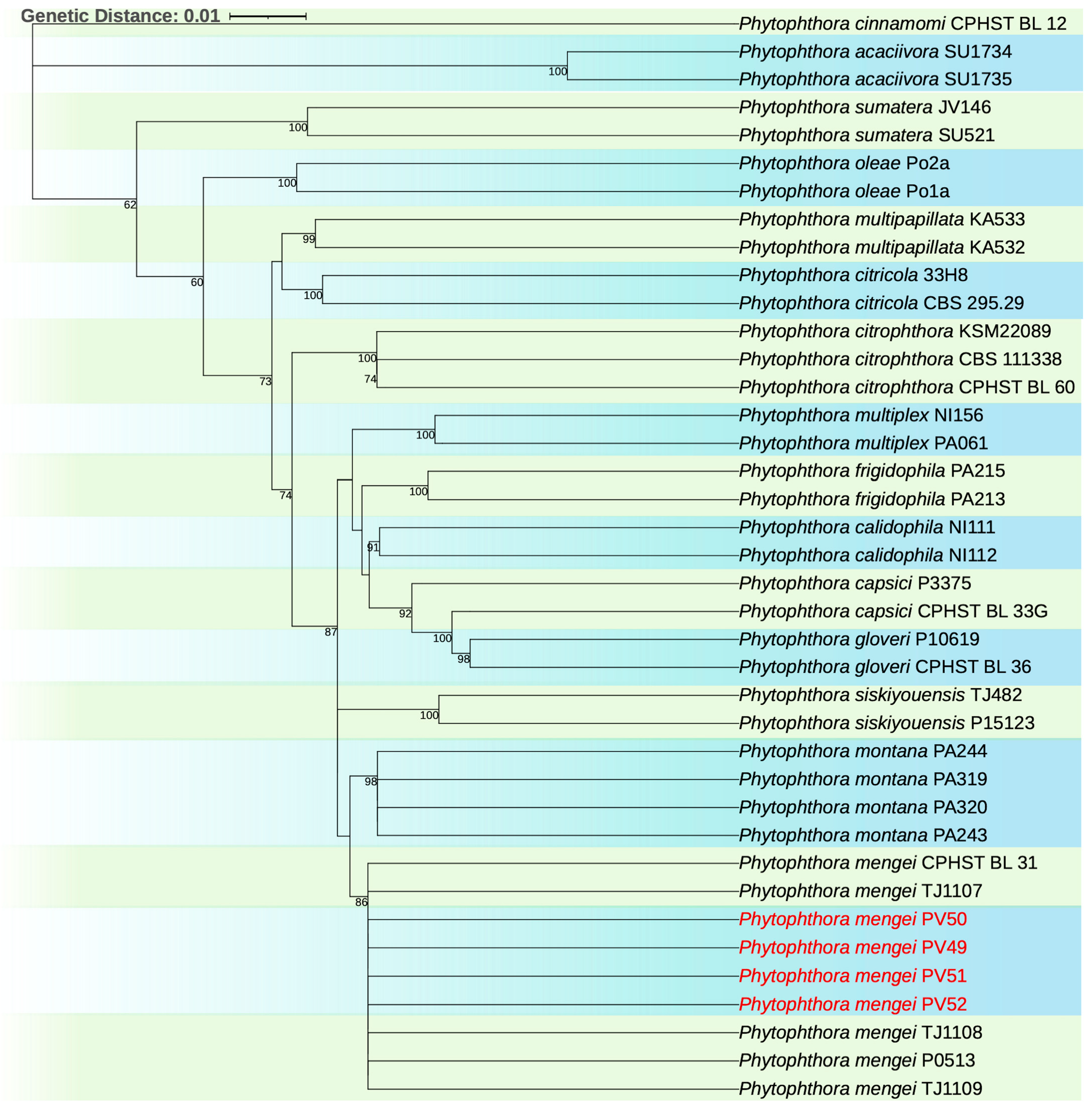
2.4. Phylogenetic Analysis
2.5. Pathogenicity Tests on Avocado Plants with Root Isolate
2.6. Pathogenicity Tests with Canker Isolate on Rootstocks
2.7. Pathogenicity Tests on Avocado Fruits
2.8. Fungicide Sensitivity
3. Results
3.1. Sampling and Isolation
3.2. Morphological Characterization
3.3. Molecular Characterization
3.4. Phylogenetic Analysis
3.5. Pathogenicity Tests on Avocado Plants with Root Isolate
3.6. Pathogenicity Tests with Canker Isolate on Rootstocks
3.7. Pathogenicity Tests on Avocado Fruits
3.8. Fungicide Sensitivity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arauéjo, R.G.; Rodriéguez-Jasso, R.M.; Ruiz, H.A.; Pintado, M.M.; Aguilar, C. Avocado by-products: Nutritional and functional properties. Trends Food Sci. Technol. 2018, 80, 51–60. [Google Scholar] [CrossRef]
- SIAP (Servicio de Información Alimentaria y Pesquera). Available online: https://nube.agricultura.gob.mx/avance_agricola/ (accessed on 13 November 2024).
- Zentmyer, G.A. Phytophthora cinnamomi and the Diseases It Causes; Phytopathological Monograph No 10; The American Phytopathological Society (APS Press): St. Paul, MN, USA, 1980; p. 96. [Google Scholar]
- Ramiérez-Gil, J.G.; Castanéeda-Saénchez, D.A.; Morales-Osorio, J.G. Estudios etioloégicos de la marchitez del aguacate en Antioquia-Colombia. Rev. Ceres 2014, 61, 50–61. [Google Scholar] [CrossRef]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide, 1st ed.; The American Phytopathological Society (APS Press): St. Paul, MN, USA, 1996. [Google Scholar]
- Hardham, A.R.; Blackman, L.M. Phytophthora cinnamomi . Mol. Plant Pathol. 2018, 19, 260–285. [Google Scholar] [CrossRef]
- Zentmyer, G.A.; Jefferson, L.; Hickman, C.J.; Chang-Ho, Y. Studies of Phytophthora citricola, isolated from Persea americana. Mycologia 1974, 66, 830–845. [Google Scholar] [CrossRef]
- Coffey, M.D.; Oudemans, P.; Ouimette, D. Phytophthora citricola: Another cause of avocado decline. CAS Yearb. 1988, 72, 127–131. [Google Scholar]
- Hong, C.X.; Gallegly, M.E.; Browne, G.T.; Bhat, R.G.; Richardson, P.A.; Kong, P. The avocado subgroup of Phytophthora citricola constitutes a distinct species, Phytophthora mengei sp. nov. Mycologia 2009, 101, 833–840. [Google Scholar] [CrossRef]
- EI-Hamalawi, Z.A.; Menge, J.A. Avocado trunk canker disease caused by Phytophthora citricola: Investigation of factors affecting infection and disease development. Plant Dis. 1994, 78, 260–264. [Google Scholar] [CrossRef]
- Abad, Z.G.; Burgess, T.I.; Redford, A.J.; Bienapfl, J.C.; Srivastava, S.; Mathew, R.; Jennings, K. IDphy: An International Online Resource for Molecular and Morphological Identification of Phytophthora. Plant Dis. 2023, 107, 987–998. [Google Scholar] [CrossRef]
- Zentmyer, G.A.; Jefferson, L.; Hickman, C.J. Another species of Phytophthora on avocado in California. CAS Yearb. 1973, 56, 125–129. [Google Scholar]
- Oudemans, P.; Forster, H.; Coffey, M.D. Evidence for distinct isozymes subgroups within Phytophthora citricola and close relationships with P. capsici and P. citrophthora. Mycol. Res. 1994, 98, 189–199. [Google Scholar] [CrossRef]
- Fucikovsky, L.; Luna, I. Avocado fruit diseases and their control in Mexico. Yearb. South Afr. Avocado Grow. Assoc. 1987, 10, 119–121. [Google Scholar]
- Bezuidenhout, C.M.; Denman, S.; Kirk, S.A.; Botha, W.J.; Mostert, L.; McLeod, A. Phytophthora taxa associated with cultivated Agathosma, with emphasis on the P. citricola complex and P. capensis sp. nov. Persoonia 2010, 25, 32–49. [Google Scholar] [CrossRef] [PubMed]
- Henricot, B.; Perez, S.A.; Jung, T. Phytophthora pachypleura sp. nov., a new species causing root rot of Aucuba japonica and other ornamentals in the United Kingdom. Plant Pathol. 2014, 63, 1095–1109. [Google Scholar] [CrossRef]
- Martin, F.N.; Blair, J.E.; Coffey, M.D. A combined mitochondrial and nuclear multilocus phylogeny of the genus Phytophthora. Fungal Genet. Biol. 2014, 66, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.; Milenković, I.; Balci, Y.; Janoušek, J.; Kudláček, T.; Nagy, Z.; Baharuddin, B.; Bakonyi, J.; Broders, K.; Cacciola, S.; et al. Worldwide forest surveys reveal forty-three new species in Phytophthora major clade 2 with fundamental implications for the evolution and biogeography of the genus and global plant biosecurity. Studies Mycol. 2024, 107, 251–388. [Google Scholar] [CrossRef]
- Burgess, T.I.; Edwards, J.; Drenth, A.; Massenbauer, T.; Cunnington, J.; Mostowfizadeh-Ghalamfarsa, R.; Dinh, Q.; Liew, E.C.Y.; White, D.; Scott, P.; et al. Current status of Phytophthora in Australia. Persoonia 2021, 47, 151–177. [Google Scholar] [CrossRef]
- Ceja-Torres, L.F.; Teéliz-Ortiz, D.; Osada-Kawasone, S.; Morales-García, J.L. Etiologiéa, distribucioén e incidencia del cancro del aguacate Persea americana Mill. en cuatro municipios del Estado de Michoacaén, Meéxico. Rev. Mex. Fitopatol. 2000, 18, 79–86. [Google Scholar]
- Ochoa-Ascencio, S.; Santacruz-Ulibarri, H.; Salazar-Garciéa, S. Phytophthora heveae causing basal rot of avocado fruit in Mexico. In Proceedings of the VII World Avocado Congress, Cairns, QLD, Australia, 5–9 September 2011. [Google Scholar]
- Romero-Cova, S.; Solís-Aragón, M.G. Nuevos hospedantes de algunas especies del género Phytophthora (Phycomycetes: Pythiaceae) en México. Agrociencia 1996, 30, 241–247. [Google Scholar]
- Menge, J.A.; Ploetz, R.C. Diseases of avocado. In Diseases of Tropical Fruit Crops; Ploetz, R.C., Ed.; CABI Publishing: Cambridge, MA, USA, 2003; pp. 35–71. [Google Scholar]
- EPPO. Phytophthora boehmeriae (PHYTBM) World Distribution. Available online: https://gd.eppo.int/taxon/PHYTBM/distribution (accessed on 10 February 2025).
- Mondragoén-Flores, A.; Manosalva, P.; Ochoa-Ascencio, S.; Diéaz-Celaya, M.; Rodriéguez-Alvarado, G.; Fernaéndez-Pavía, S.P. Characterization and fungicides sensitivity of Phytophthora cinnamomi isolates causing avocado root rot in Zitacuaro, Michoacan. Rev. Mex. Fitopatol. 2022, 40, 1–23. [Google Scholar] [CrossRef]
- Tuite, J. Plant Pathological Methods. Fungi and Bacteria; Burgess Publishing Co.: Minneapolis, MN, USA, 1969. [Google Scholar]
- Fernández-Pavía, S.P.; Díaz-Celaya, M.; Rodríguez-Alvarado, G.; Gómez-Dorantes, N.; Abad, Z.G.; Félix-Gastélum, R.; Ochoa-Ascencio, S.; Garay-Serrano, E.; Mondragón-Flores, A.; Soto-Plancarte, A.; et al. Laboratory Manual of Phytophthora, 1st ed.; Mexican Society of Phytopathology: Mexico City, Mexico, 2020. [Google Scholar]
- Soto-Plancarte, A.; Díaz-Celaya, M.; Rodríguez-Alvarado, G.; Fernández-Pavía, Y.L.; Silva-Rojas, H.V.; Pedraza-Santos, M.E.; Salgado-Garciglia, R.; Bourret, T.B.; Fernández-Pavía, S.P. The heterogeneity of ornamental plants in nurseries increases the chance of finding new hosts for Phytophthora. J. Fungi 2025, 11, 187. [Google Scholar] [CrossRef]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A molecular phylogeny of Phytophthora and related oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Robideau, G.P.; De Cock, A.W.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Désaulniers, N.; Eggertson, Q.A.; Gachon, C.M.M.; et al. DNA barcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Bonfield, J.K.; Whitwham, A. Gap5—Editing the billion fragment sequence assembly. Bioinformatics 2010, 26, 1699–1703. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop, New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef]
- iTOL: Interactive Tree of Life. Available online: https://itol.embl.de/ (accessed on 18 October 2024).
- Chen, D.W.; Zentmyer, G.A. Production of sporangia by Phytophthora cinnamomi in axenic culture. Mycologia 1970, 62, 397–402. [Google Scholar] [CrossRef]
- Gray, M.A.; Hao, W.; Forster, H.; Adaskaveg, J.E. Baseline sensitivities of new fungicides and their toxicity to selected life stages of Phytophthora species from citrus in California. Plant Dis. 2018, 102, 734–742. [Google Scholar] [CrossRef]
- Adaskaveg, J.E.; Hao, W.; Forster, H. Postharvest strategies for managing Phytophthora brown rot of citrus using potassium phosphite in combination with heat treatments. Plant Dis. 2015, 99, 1477–1482. [Google Scholar] [CrossRef]
- Hu, J.; Hong, C.; Stromberg, E.; Moorman, G. Mefenoxam sensitivity in Phytophthora cinnamomi isolates. Plant Dis. 2010, 94, 39–44. [Google Scholar] [CrossRef]
- Fucikovsky, L. Enfermedades del Aguacate en el Estado de México. Circ. DAGEM 1968, 45, 16. [Google Scholar]
- Téliz, O.D. Enfermedades del aguacate. In El aguacate y su Manejo Integrado; Téliz, O.D., Ed.; Mundi-Prensa: Mexico City, México, 2000; pp. 137–182. [Google Scholar]
- Coffey, M.D. Citricola: Avocado killer. Calif. Grower 1989, 13, 8. [Google Scholar]
- Coffey, M.D.; Klure, L.J.; Bower, L.A. Variability in sensitivity to metalaxyl of isolates of Phytophthora cinnamomi and Phytophthora citricola. Phytopathology 1984, 74, 417–422. [Google Scholar] [CrossRef]
- Bhat, R.G.; Browne, G.T. Genetic diversity in populations of Phytophthora citricola associated with horticultural crops in California. Plant Dis. 2007, 91, 1556–1563. [Google Scholar] [CrossRef]
- Belisle, R.J.; McKee, B.; Hao, W.; Crowley, M.; Arpaia, M.L.; Miles, T.D.; Adaskaveg, J.E.; Manosalva, P. Phenotypic characterization of genetically distinct Phytophthora cinnamomi isolates from avocado. Phytopathology 2019, 109, 384–394. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potassium Phosphite | Metalaxyl-M | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Concentration (μg/mL−1) | Growth (mm) | Inhibition (%) | Concentration (μg/mL−1) | Growth (mm) | Inhibition (%) | ||||
| 0 | 21.7 | a * | 0 | a | 0 | 19.8 | a | 0 | a |
| 5 | 8.5 | b | 61.0 | b | 0.05 | 19.5 | ab | 1.7 | a |
| 10 | 5.7 | b | 73.9 | c | 0.15 | 17.3 | b | 12.5 | b |
| 25 | 3.2 | d | 85.4 | d | 0.5 | 7.8 | c | 60.4 | d |
| 50 | 1.0 | e | 95.4 | e | 1 | 6.3 | cd | 68.1 | dc |
| 100 | 0 | f | 100 | f | 3 | 5.16 | d | 73.9 | c |
| 300 | 0 | f | 100 | f | 5 | 5.0 | d | 74.9 | c |
| 600 | 0 | f | 100 | f | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mondragón-Flores, A.; Soto-Plancarte, A.; Rodríguez-Alvarado, G.; Manosalva, P.; Ochoa-Ascencio, S.; Hoyt, B.; Gómez-Dorantes, N.; Fernández-Pavía, S.P. First Report of Phytophthora mengei Causing Root Rot and Canker in Avocado (Persea americana) in Michoacán, Mexico. Microorganisms 2025, 13, 1471. https://doi.org/10.3390/microorganisms13071471
Mondragón-Flores A, Soto-Plancarte A, Rodríguez-Alvarado G, Manosalva P, Ochoa-Ascencio S, Hoyt B, Gómez-Dorantes N, Fernández-Pavía SP. First Report of Phytophthora mengei Causing Root Rot and Canker in Avocado (Persea americana) in Michoacán, Mexico. Microorganisms. 2025; 13(7):1471. https://doi.org/10.3390/microorganisms13071471
Chicago/Turabian StyleMondragón-Flores, Alejandra, Alejandro Soto-Plancarte, Gerardo Rodríguez-Alvarado, Patricia Manosalva, Salvador Ochoa-Ascencio, Benjamin Hoyt, Nuria Gómez-Dorantes, and Sylvia Patricia Fernández-Pavía. 2025. "First Report of Phytophthora mengei Causing Root Rot and Canker in Avocado (Persea americana) in Michoacán, Mexico" Microorganisms 13, no. 7: 1471. https://doi.org/10.3390/microorganisms13071471
APA StyleMondragón-Flores, A., Soto-Plancarte, A., Rodríguez-Alvarado, G., Manosalva, P., Ochoa-Ascencio, S., Hoyt, B., Gómez-Dorantes, N., & Fernández-Pavía, S. P. (2025). First Report of Phytophthora mengei Causing Root Rot and Canker in Avocado (Persea americana) in Michoacán, Mexico. Microorganisms, 13(7), 1471. https://doi.org/10.3390/microorganisms13071471