
Insights on the Horizontal Gene Transfer of Carbapenemase Determinants in the Opportunistic Pathogen Acinetobacter baumannii


Abstract
:
1. Introduction
2. Clinical Importance of Acinetobacter baumannii
3. Evolution of Resistance towards Large Spectrum Antibiotics
4. Carbapenem Resistance in Acinetobacter baumannii
5. Production of Carbapenemases
5.1. Intrinsic β-Lactamases
5.2. Acquired β-Lactamases
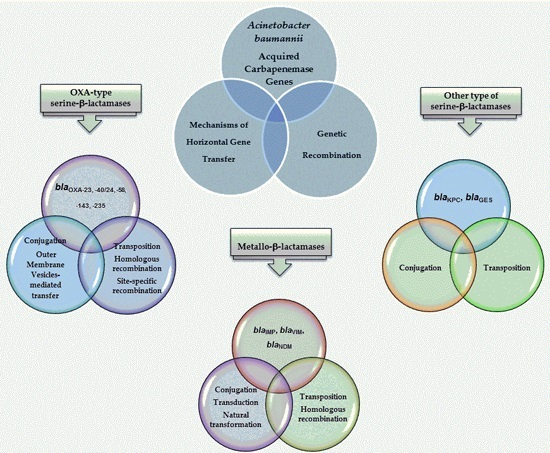
6. Mechanisms of Acquisition of Carbapenemase Determinants in Acinetobacter baumannii
6.1. Mechanisms Involved in the Movement and Dissemination of CHDLs
6.1.1. OXA-23 (and OXA-23-Like)
6.1.2. OXA-40/24 (and OXA-40-Like)
6.1.3. OXA-58 (and OXA-58-Like)
6.1.4. OXA-143 (and OXA-143-Like)
6.1.5. OXA-235 (and OXA-235-Like)
6.2. Mechanisms Involved in the Movement and Dissemination of MBL Genes
6.2.1. IMP-Type
6.2.2. VIM-Type
6.2.3. NDM-Type
6.3. Mechanisms Involved in the Movement and Dissemination of Class A β-Lactamases
6.3.1. KPC-Type
6.3.2. GES-Type
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interests
References
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.H.; Hu, Z.Q. Acinetobacter: A potential reservoir and dispenser for beta-lactamases. Crit. Rev. Microbiol. 2012, 38, 30–51. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.A.; Amyes, S.G. OXA beta-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Naas, T.; Nordmann, P. Diversity, epidemiology, and genetics of class D beta-lactamases. Antimicrob. Agents Chemother. 2010, 54, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Gombac, F.; Riccio, M.L.; Rossolini, G.M.; Lagatolla, C.; Tonin, E.; Monti-Bragadin, C.; Lavenia, A.; Dolzani, L. Molecular characterization of integrons in epidemiologically unrelated clinical isolates of Acinetobacter baumannii from Italian hospitals reveals a limited diversity of gene cassette arrays. Antimicrob. Agents Chemother. 2002, 46, 3665–3668. [Google Scholar] [CrossRef] [PubMed]
- Dijkshoorn, L.; Nemec, A.; Seifert, H. An increasing threat in hospitals: Multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 2007, 5, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Wendt, C.; Dietze, B.; Dietz, E.; Ruden, H. Survival of Acinetobacter baumannii on dry surfaces. J. Clin. Microbiol. 1997, 35, 1394–1397. [Google Scholar] [PubMed]
- Jawad, A.; Seifert, H.; Snelling, A.M.; Heritage, J.; Hawkey, P.M. Survival of Acinetobacter baumannii on dry surfaces: Comparison of outbreak and sporadic isolates. J. Clin. Microbiol. 1998, 36, 1938–1941. [Google Scholar] [PubMed]
- Da Silva, G.J. Resistência aos Antibióticos Beta-Lactâmicos em Isolados Clínicos de Acinetobacter spp. Caracterização Molecular de Novas Carbapenemases, IMP-5 e OXA-33, e Estudo Clonal Entre os Isolados Resistentes ao Imipenemo. Ph.D. Thesis, University of Coimbra, Coimbra, Portugal, December 2002. [Google Scholar]
- Da Silva, G.; Dijkshoorn, L.; van der Reijden, T.; van Strijen, B.; Duarte, A. Identification of widespread, closely related Acinetobacter baumannii isolates in Portugal as a subgroup of European clone II. Clin. Microbiol. Infect. 2007, 13, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, G.J.; Mendonca, N.; Batista, G.; Duarte, A. Sequence types of Portuguese carbapenem-resistant Acinetobacter baumannii isolates collected over 10 years. J. Antimicrob. Chemother. 2010, 65, 2254–2256. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, G.J.; Quinteira, S.; Bertolo, E.; Sousa, J.C.; Gallego, L.; Duarte, A.; Peixe, L. Long-term dissemination of an OXA-40 carbapenemase-producing Acinetobacter baumannii clone in the Iberian Peninsula. J. Antimicrob. Chemother. 2004, 54, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Visca, P.; Seifert, H.; Towner, K.J. Acinetobacter infection—An emerging threat to human health. IUBMB Life 2011, 63, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Kempf, M.; Rolain, J.M. Emergence of resistance to carbapenems in Acinetobacter baumannii in Europe: Clinical impact and therapeutic options. Int. J. Antimicrob. Agents 2012, 39, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeannot, K.; Diancourt, L.; Vaux, S.; Thouverez, M.; Ribeiro, A.; Coignard, B.; Courvalin, P.; Brisse, S. Molecular epidemiology of carbapenem non-susceptible Acinetobacter baumannii in France. PLoS ONE 2014, 9, e115452. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, G.J.; van der Reijden, T.; Domingues, S.; Mendonca, N.; Petersen, K.; Dijkshoorn, L. Characterization of a novel international clonal complex (CC32) of Acinetobacter baumannii with epidemic potential. Epidemiol. Infect. 2014, 142, 1554–1558. [Google Scholar] [CrossRef] [PubMed]
- Sunenshine, R.H.; Wright, M.O.; Maragakis, L.L.; Harris, A.D.; Song, X.; Hebden, J.; Cosgrove, S.E.; Anderson, A.; Carnell, J.; Jernigan, D.B.; et al. Multidrug-resistant Acinetobacter infection mortality rate and length of hospitalization. Emerg. Infect. Dis. 2007, 13, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Choo, J.W.; Kwon, S.H.; Yu, S.N.; Lee, E.J.; Kim, T.H.; Choo, E.J.; Jeon, M.H. Risk factors for mortality in patients with Acinetobacter baumannii bacteremia. Infect. Chemother. 2013, 45, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Kasiakou, S.K. Colistin: The revival of polymyxins for the management of multidrug-resistant Gram-negative bacterial infections. Clin. Infect. Dis. 2005, 40, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Doi, Y.; Murray, G.L.; Peleg, A.Y. Acinetobacter baumannii: Evolution of antimicrobial resistance-treatment options. Semin. Respir. Crit. Care Med. 2015, 36, 85–98. [Google Scholar] [PubMed]
- Bonnin, R.A.; Nordmann, P.; Poirel, L. Screening and deciphering antibiotic resistance in Acinetobacter baumannii: A state of the art. Expert Rev. Anti Infect. Ther. 2013, 11, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Dortet, L.; Poirel, L. Carbapenem resistance in Enterobacteriaceae: Here is the storm! Trends Mol. Med. 2012, 18, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Limansky, A.S.; Mussi, M.A.; Viale, A.M. Loss of a 29-kilodalton outer membrane protein in Acinetobacter baumannii is associated with imipenem resistance. J. Clin. Microbiol. 2002, 40, 4776–4778. [Google Scholar] [CrossRef] [PubMed]
- Gehrlein, M.; Leying, H.; Cullmann, W.; Wendt, S.; Opferkuch, W. Imipenem resistance in Acinetobacter baumanii is due to altered penicillin-binding proteins. Chemotherapy 1991, 37, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Poole, K. Efflux pumps as antimicrobial resistance mechanisms. Ann. Med. 2007, 39, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Magnet, S.; Courvalin, P.; Lambert, T. Resistance-nodulation-cell division-type efflux pump involved in aminoglycoside resistance in Acinetobacter baumannii strain BM4454. Antimicrob. Agents Chemother. 2001, 45, 3375–3380. [Google Scholar] [CrossRef] [PubMed]
- Marchand, I.; Damier-Piolle, L.; Courvalin, P.; Lambert, T. Expression of the RND-type efflux pump AdeABC in Acinetobacter baumannii is regulated by the AdeRS two-component system. Antimicrob. Agents Chemother. 2004, 48, 3298–3304. [Google Scholar] [CrossRef] [PubMed]
- Heritier, C.; Poirel, L.; Lambert, T.; Nordmann, P. Contribution of acquired carbapenem-hydrolyzing oxacillinases to carbapenem resistance in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2005, 49, 3198–3202. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.S.; Yao, S.M.; Fung, C.P.; Hsieh, Y.P.; Liu, C.P.; Lin, J.F. An OXA-66/OXA-51-like carbapenemase and possibly an efflux pump are associated with resistance to imipenem in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2007, 51, 3844–3852. [Google Scholar] [CrossRef] [PubMed]
- Heritier, C.; Poirel, L.; Fournier, P.E.; Claverie, J.M.; Raoult, D.; Nordmann, P. Characterization of the naturally occurring oxacillinase of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2005, 49, 4174–4179. [Google Scholar] [CrossRef] [PubMed]
- Heritier, C.; Poirel, L.; Nordmann, P. Cephalosporinase over-expression resulting from insertion of ISAba1 in Acinetobacter baumannii. Clin. Microbiol. Infect. 2006, 12, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Turton, J.F.; Ward, M.E.; Woodford, N.; Kaufmann, M.E.; Pike, R.; Livermore, D.M.; Pitt, T.L. The role of ISAba1 in expression of OXA carbapenemase genes in Acinetobacter baumannii. FEMS Microbiol. Lett. 2006, 258, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.L.; Lee, Y.T.; Kuo, S.C.; Hsueh, P.R.; Chang, F.Y.; Siu, L.K.; Ko, W.C.; Fung, C.P. Emergence and distribution of plasmids bearing the blaOXA-51-like gene with an upstream ISAba1 in carbapenem-resistant Acinetobacter baumannii isolates in Taiwan. Antimicrob. Agents Chemother. 2010, 54, 4575–4581. [Google Scholar] [CrossRef] [PubMed]
- Turton, J.F.; Woodford, N.; Glover, J.; Yarde, S.; Kaufmann, M.E.; Pitt, T.L. Identification of Acinetobacter baumannii by detection of the blaOXA-51-like carbapenemase gene intrinsic to this species. J. Clin. Microbiol. 2006, 44, 2974–2976. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.T.; Turton, J.F.; Chen, T.L.; Wu, R.C.; Chang, W.C.; Fung, C.P.; Chen, C.P.; Cho, W.L.; Huang, L.Y.; Siu, L.K. First identification of blaOXA-51-like in non-baumannii Acinetobacter spp. J. Chemother. 2009, 21, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.T.; Kuo, S.C.; Chiang, M.C.; Yang, S.P.; Chen, C.P.; Chen, T.L.; Fung, C.P. Emergence of carbapenem-resistant non-baumannii species of Acinetobacter harboring a blaOXA-51-like gene that is intrinsic to A. baumannii. Antimicrob. Agents Chemother. 2012, 56, 1124–1127. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Poirel, L. Emerging carbapenemases in Gram-negative aerobes. Clin. Microbiol. Infect. 2002, 8, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.S.; Lee, W.S.; Lam, C.; Hsu, C.W.; Chen, R.J.; Hsueh, P.R. Carbapenemase-producing Gram-negative bacteria: Current epidemics, antimicrobial susceptibility and treatment options. Future Microbiol. 2015, 10, 407–425. [Google Scholar] [CrossRef] [PubMed]
- Meletis, G. Carbapenem resistance: Overview of the problem and future perspectives. Ther. Adv. Infect. Dis. 2016, 3, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Ambler, R.P. The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1980, 289, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Yum, J.H.; Yong, D.; Lee, H.M.; Kim, H.D.; Docquier, J.D.; Rossolini, G.M.; Chong, Y. Novel acquired metallo-beta-lactamase gene, blaSIM-1, in a class 1 integron from Acinetobacter baumannii clinical isolates from Korea. Antimicrob. Agents Chemother. 2005, 49, 4485–4491. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Roh, K.H.; Lee, Y.; Chung, H.S.; Yum, J.H.; Yong, D.; Lee, K.; Chong, Y. Clonal change of blaSIM-1-carrying Acinetobacter spp. from 2003 to 2008 in the hospital where it was initially discovered. Microb. Drug Resist. 2013, 19, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Bush, K. The ABCD’s of beta-lactamase nomenclature. J. Infect. Chemother. 2013, 19, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Higgins, P.G.; Poirel, L.; Lehmann, M.; Nordmann, P.; Seifert, H. OXA-143, a novel carbapenem-hydrolyzing class D beta-lactamase in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 5035–5038. [Google Scholar] [CrossRef] [PubMed]
- Higgins, P.G.; Perez-Llarena, F.J.; Zander, E.; Fernandez, A.; Bou, G.; Seifert, H. OXA-235, a novel class D beta-lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2013, 57, 2121–2126. [Google Scholar] [CrossRef] [PubMed]
- Elsherif, R.; Ismail, D.; Elawady, S.; Jastaniah, S.; Al-Masaudi, S.; Harakeh, S.; Karrouf, G. Boronic acid disk diffusion for the phenotypic detection of polymerase chain reaction-confirmed, carbapenem-resistant, gram-negative bacilli isolates. BMC Microbiol. 2016, 16, 135. [Google Scholar] [CrossRef] [PubMed]
- Von Wintersdorff, C.J.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.; Wolffs, P.F. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Bergogne-Berezin, E.; Towner, K.J. Acinetobacter spp. as nosocomial pathogens: Microbiological, clinical, and epidemiological features. Clin. Microbiol. Rev. 1996, 9, 148–165. [Google Scholar] [PubMed]
- Fu, Y.; Jiang, J.; Zhou, H.; Jiang, Y.; Fu, Y.; Yu, Y.; Zhou, J. Characterization of a novel plasmid type and various genetic contexts of blaOXA-58 in Acinetobacter spp. from multiple cities in China. PLoS ONE 2014, 9, e84680. [Google Scholar] [CrossRef] [PubMed]
- Heritier, C.; Dubouix, A.; Poirel, L.; Marty, N.; Nordmann, P. A nosocomial outbreak of Acinetobacter baumannii isolates expressing the carbapenem-hydrolysing oxacillinase OXA-58. J. Antimicrob. Chemother. 2005, 55, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Towner, K.J.; Evans, B.; Villa, L.; Levi, K.; Hamouda, A.; Amyes, S.G.; Carattoli, A. Distribution of intrinsic plasmid replicase genes and their association with carbapenem-hydrolyzing class D beta-lactamase genes in European clinical isolates of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 2154–2159. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Bercot, B.; Millemann, Y.; Bonnin, R.A.; Pannaux, G.; Nordmann, P. Carbapenemase-producing Acinetobacter spp. in Cattle, France. Emerg. Infect. Dis. 2012, 18, 523–525. [Google Scholar] [CrossRef] [PubMed]
- Merabishvili, M.; Vandenheuvel, D.; Kropinski, A.M.; Mast, J.; de Vos, D.; Verbeken, G.; Noben, J.P.; Lavigne, R.; Vaneechoutte, M.; Pirnay, J.P. Characterization of newly isolated lytic bacteriophages active against Acinetobacter baumannii. PLoS ONE 2014, 9, e104853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, G.H.; Wang, J.L.; Wen, F.S.; Chang, K.M.; Kuo, C.F.; Lin, C.H.; Luo, H.R.; Hung, C.H. Isolation and characterization of phikm18p, a novel lytic phage with therapeutic potential against extensively drug resistant Acinetobacter baumannii. PLoS ONE 2012, 7, e46537. [Google Scholar] [CrossRef] [PubMed]
- Vaneechoutte, M.; Young, D.M.; Ornston, L.N.; de Baere, T.; Nemec, A.; van der Reijden, T.; Carr, E.; Tjernberg, I.; Dijkshoorn, L. Naturally transformable Acinetobacter sp. strain ADP1 belongs to the newly described species Acinetobacter baylyi. Appl. Environ. Microbiol. 2006, 72, 932–936. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.S.; Don, M.; Merkier, A.K.; Bistue, A.J.; Zorreguieta, A.; Centron, D.; Tolmasky, M.E. Naturally competent Acinetobacter baumannii clinical isolate as a convenient model for genetic studies. J. Clin. Microbiol. 2010, 48, 1488–1490. [Google Scholar] [CrossRef] [PubMed]
- Wilharm, G.; Piesker, J.; Laue, M.; Skiebe, E. DNA uptake by the nosocomial pathogen Acinetobacter baumannii occurs during movement along wet surfaces. J. Bacteriol. 2013, 195, 4146–4153. [Google Scholar] [CrossRef] [PubMed]
- Rumbo, C.; Fernandez-Moreira, E.; Merino, M.; Poza, M.; Mendez, J.A.; Soares, N.C.; Mosquera, A.; Chaves, F.; Bou, G. Horizontal transfer of the OXA-24 carbapenemase gene via outer membrane vesicles: A new mechanism of dissemination of carbapenem resistance genes in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 3084–3090. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Amyes, S. OXA beta-lactamases in Acinetobacter: The story so far. J. Antimicrob. Chemother. 2006, 57, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Pitout, J.D.; Nordmann, P. Carbapenemases: Molecular diversity and clinical consequences. Future Microbiol. 2007, 2, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Paton, R.; Miles, R.S.; Hood, J.; Amyes, S.G.; Miles, R.S.; Amyes, S.G. ARI 1: Beta-lactamase-mediated imipenem resistance in Acinetobacter baumannii. Int. J. Antimicrob. Agents 1993, 2, 81–87. [Google Scholar] [CrossRef]
- Donald, H.M.; Scaife, W.; Amyes, S.G.; Young, H.K. Sequence analysis of ARI-1, a novel OXA beta-lactamase, responsible for imipenem resistance in Acinetobacter baumannii 6B92. Antimicrob. Agents Chemother. 2000, 44, 196–169. [Google Scholar] [CrossRef] [PubMed]
- Mugnier, P.D.; Poirel, L.; Naas, T.; Nordmann, P. Worldwide dissemination of the blaOXA-23 carbapenemase gene of Acinetobacter baumannii. Emerg. Infect. Dis. 2010, 16, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Turton, J.F.; Baddal, B.; Perry, C. Use of the accessory genome for characterization and typing of Acinetobacter baumannii. J. Clin. Microbiol. 2011, 49, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Nigro, S.J.; Hall, R.M. Structure and context of Acinetobacter transposons carrying the oxa23 carbapenemase gene. J. Antimicrob. Chemother. 2016, 71, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Yan, Z.; Zhang, Z.; Zhou, Q.; Zhou, J.; Wakeland, E.K.; Fang, X.; Xuan, Z.; Shen, D.; Li, Q.Z. Complete genome analysis of three Acinetobacter baumannii clinical isolates in China for insight into the diversification of drug resistance elements. PLoS ONE 2013, 8, e66584. [Google Scholar] [CrossRef] [PubMed]
- Pagano, M.; Martins, A.F.; Machado, A.B.; Barin, J.; Barth, A.L. Carbapenem-susceptible Acinetobacter baumannii carrying the ISAba1 upstream blaOXA-51-like gene in Porto Alegre, southern Brazil. Epidemiol. Infect. 2013, 141, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Corvec, S.; Poirel, L.; Naas, T.; Drugeon, H.; Nordmann, P. Genetics and expression of the carbapenem-hydrolyzing oxacillinase gene blaOXA-23 in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2007, 51, 1530–1533. [Google Scholar] [CrossRef] [PubMed]
- Mugnier, P.D.; Poirel, L.; Nordmann, P. Functional analysis of insertion sequence ISAba1, responsible for genomic plasticity of Acinetobacter baumannii. J. Bacteriol. 2009, 191, 2414–2418. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, I.; Novais, A.; Grosso, F.; Rodrigues, C.; Chifiriuc, M.C.; Lazar, V.; Peixe, L. Snapshot on carbapenemase-producing Pseudomonas aeruginosa and Acinetobacter baumannii in Bucharest hospitals reveals unusual clones and novel genetic surroundings for blaOXA-23. J. Antimicrob. Chemother. 2015, 70, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Hamidian, M.; Kenyon, J.J.; Holt, K.E.; Pickard, D.; Hall, R.M. A conjugative plasmid carrying the carbapenem resistance gene blaOXA-23 in AbaR4 in an extensively resistant GC1 Acinetobacter baumannii isolate. J. Antimicrob. Chemother. 2014, 69, 2625–2628. [Google Scholar] [CrossRef] [PubMed]
- Nigro, S.; Hall, R.M. Distribution of the blaOXA-23-containing transposons Tn2006 and Tn2008 in Australian carbapenem-resistant Acinetobacter baumannii isolates. J. Antimicrob. Chemother. 2015, 70, 2409–2411. [Google Scholar] [CrossRef] [PubMed]
- Bou, G.; Oliver, A.; Martinez-Beltran, J. OXA-24, a novel class D beta-lactamase with carbapenemase activity in an Acinetobacter baumannii clinical strain. Antimicrob. Agents Chemother. 2000, 44, 1556–1561. [Google Scholar] [CrossRef] [PubMed]
- Heritier, C.; Poirel, L.; Aubert, D.; Nordmann, P. Genetic and functional analysis of the chromosome-encoded carbapenem-hydrolyzing oxacillinase OXA-40 of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2003, 47, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Lolans, K.; Rice, T.W.; Munoz-Price, L.S.; Quinn, J.P. Multicity outbreak of carbapenem-resistant Acinetobacter baumannii isolates producing the carbapenemase OXA-40. Antimicrob. Agents Chemother. 2006, 50, 2941–2945. [Google Scholar] [CrossRef] [PubMed]
- Merino, M.; Acosta, J.; Poza, M.; Sanz, F.; Beceiro, A.; Chaves, F.; Bou, G. OXA-24 carbapenemase gene flanked by XerC/XerD-like recombination sites in different plasmids from different Acinetobacter species isolated during a nosocomial outbreak. Antimicrob. Agents Chemother. 2010, 54, 2724–2727. [Google Scholar] [CrossRef] [PubMed]
- Grosso, F.; Quinteira, S.; Poirel, L.; Novais, A.; Peixe, L. Role of common blaOXA-24/OXA-40-carrying platforms and plasmids in the spread of OXA-24/OXA-40 among Acinetobacter species clinical isolates. Antimicrob. Agents Chemother. 2012, 56, 3969–3972. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, M.M.; Giani, T.; D’Arezzo, S.; Capone, A.; Petrosillo, N.; Visca, P.; Luzzaro, F.; Rossolini, G.M. Characterization of pABVA01, a plasmid encoding the OXA-24 carbapenemase from Italian isolates of Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 3528–3533. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.B.; Adams-Haduch, J.M.; Bogdanovich, T.; Pasculle, A.W.; Quinn, J.P.; Wang, H.N.; Doi, Y. Identification of diverse OXA-40 group carbapenemases, including a novel variant, OXA-160, from Acinetobacter baumannii in Pennsylvania. Antimicrob. Agents Chemother. 2011, 55, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Marque, S.; Poirel, L.; Heritier, C.; Brisse, S.; Blasco, M.D.; Filip, R.; Coman, G.; Naas, T.; Nordmann, P. Regional occurrence of plasmid-mediated carbapenem-hydrolyzing oxacillinase OXA-58 in Acinetobacter spp. in Europe. J. Clin. Microbiol. 2005, 43, 4885–4888. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.L.; Wu, R.C.; Shaio, M.F.; Fung, C.P.; Cho, W.L. Acquisition of a plasmid-borne blaOXA-58 gene with an upstream IS1008 insertion conferring a high level of carbapenem resistance to Acinetobacter baumannii. Antimicrob. Agents Chemother. 2008, 52, 2573–2580. [Google Scholar] [CrossRef] [PubMed]
- Bertini, A.; Poirel, L.; Mugnier, P.D.; Villa, L.; Nordmann, P.; Carattoli, A. Characterization and PCR-based replicon typing of resistance plasmids in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2010, 54, 4168–4177. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Nordmann, P. Genetic structures at the origin of acquisition and expression of the carbapenem-hydrolyzing oxacillinase gene blaOXA-58 in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2006, 50, 1442–1448. [Google Scholar] [CrossRef] [PubMed]
- Ravasi, P.; Limansky, A.S.; Rodriguez, R.E.; Viale, A.M.; Mussi, M.A. ISAba825, a functional insertion sequence modulating genomic plasticity and blaOXA-58 expression in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 917–920. [Google Scholar] [CrossRef] [PubMed]
- Bertini, A.; Poirel, L.; Bernabeu, S.; Fortini, D.; Villa, L.; Nordmann, P.; Carattoli, A. Multicopy blaOXA-58 gene as a source of high-level resistance to carbapenems in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2007, 51, 2324–2328. [Google Scholar] [CrossRef] [PubMed]
- Girlich, D.; Damaceno, Q.S.; Oliveira, A.C.; Nordmann, P. OXA-253, a variant of the carbapenem-hydrolyzing class D beta-lactamase OXA-143 in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2014, 58, 2976–2978. [Google Scholar] [CrossRef] [PubMed]
- Cayo, R.; Rodrigues-Costa, F.; Matos, A.P.; Carvalhaes, C.G.; Jove, T.; Gales, A.C. Identification of a new integron harboring blaIMP-10 in carbapenem-resistant Acinetobacter baumannii clinical isolates. Antimicrob. Agents Chemother. 2015, 59, 3687–3689. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, G.J.; Correia, M.; Vital, C.; Ribeiro, G.; Sousa, J.C.; Leitão, R.; Peixe, L.; Duarte, A. Molecular characterization of blaIMP-5, a new integron-borne metallo-β-lactamase gene from Acinetobacter baumannii nosocomial isolate in Portugal. FEMS Microbiol. Lett. 2002, 215, 33–39. [Google Scholar] [CrossRef]
- Riccio, M.L.; Franceschini, N.; Boschi, L.; Caravelli, B.; Cornaglia, G.; Fontana, R.; Amicosante, G.; Rossolini, G.M. Characterization of the metallo-beta-lactamase determinant of Acinetobacter baumannii AC-54/97 reveals the existence of blaIMP allelic variants carried by gene cassettes of different phylogeny. Antimicrob. Agents Chemother. 2000, 44, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.W.; Afzal-Shah, M.; Houang, E.T.; Palepou, M.I.; Lyon, D.J.; Woodford, N.; Livermore, D.M. IMP-4, a novel metallo-beta-lactamase from nosocomial Acinetobacter spp. collected in Hong Kong between 1994 and 1998. Antimicrob. Agents Chemother. 2001, 45, 710–714. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.E.; Castanheira, M.; Toleman, M.A.; Sader, H.S.; Jones, R.N.; Walsh, T.R. Characterization of an integron carrying blaIMP-1 and a new aminoglycoside resistance gene, aac(6')-31, and its dissemination among genetically unrelated clinical isolates in a Brazilian hospital. Antimicrob. Agents Chemother. 2007, 51, 2611–2614. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Lin, J.Y.; Chu, C.; Su, L.H.; Lin, T.Y.; Chiu, C.H. Integron-associated imipenem resistance in Acinetobacter baumannii isolated from a regional hospital in Taiwan. Int. J. Antimicrob. Agents 2006, 27, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.L.; Ma, L.; Chang, J.C.; Su, L.H.; Chu, C.; Leu, H.S.; Siu, L.K. Variable resistance patterns of integron-associated multidrug-resistant Acinetobacter baumannii isolates in a surgical intensive care unit. Microb. Drug Resist. 2004, 10, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.; Nielsen, K.M.; da Silva, G.J. The blaIMP-5-carrying integron in a clinical Acinetobacter baumannii strain is flanked by miniature inverted-repeat transposable elements (MITEs). J. Antimicrob. Chemother. 2011, 66, 2667–2668. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.; Toleman, M.A.; Nielsen, K.M.; da Silva, G.J. Identical miniature inverted repeat transposable elements (MITE) flanks class 1 integrons in clinical isolates of Acinetobacter spp. J. Clin. Microbiol. 2013, 51, 2382–2384. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.; Harms, K.; Fricke, W.F.; Johnsen, P.J.; da Silva, G.J.; Nielsen, K.M. Natural transformation facilitates transfer of transposons, integrons and gene cassettes between bacterial species. PLoS Pathog. 2012, 8, e1002837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsakris, A.; Ikonomidis, A.; Pournaras, S.; Tzouvelekis, L.S.; Sofianou, D.; Legakis, N.J.; Maniatis, A.N. VIM-1 metallo-beta-lactamase in Acinetobacter baumannii. Emerg. Infect. Dis. 2006, 12, 981–983. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.Y.; Chen, T.L.; Lu, P.L.; Tsai, C.A.; Cho, W.L.; Chang, F.Y.; Fung, C.P.; Siu, L.K. Dissemination of multidrug-resistant, class 1 integron-carrying Acinetobacter baumannii isolates in Taiwan. Clin. Microbiol. Infect. 2008, 14, 1010–1019. [Google Scholar] [CrossRef] [PubMed]
- Yum, J.H.; Yi, K.; Lee, H.; Yong, D.; Lee, K.; Kim, J.M.; Rossolini, G.M.; Chong, Y. Molecular characterization of metallo-beta-lactamase-producing Acinetobacter baumannii and Acinetobacter genomospecies 3 from Korea: Identification of two new integrons carrying the blaVIM-2 gene cassettes. J. Antimicrob. Chemother. 2002, 49, 837–840. [Google Scholar] [CrossRef] [PubMed]
- Loli, A.; Tzouvelekis, L.S.; Gianneli, D.; Tzelepi, E.; Miriagou, V. Outbreak of Acinetobacter baumannii with chromosomally encoded VIM-1 undetectable by imipenem-EDTA synergy tests. Antimicrob. Agents Chemother. 2008, 52, 1894–1896. [Google Scholar] [CrossRef] [PubMed]
- Tsakris, A.; Ikonomidis, A.; Poulou, A.; Spanakis, N.; Vrizas, D.; Diomidous, M.; Pournaras, S.; Markou, F. Clusters of imipenem-resistant Acinetobacter baumannii clones producing different carbapenemases in an intensive care unit. Clin. Microbiol. Infect. 2008, 14, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Wailan, A.M.; Sidjabat, H.E.; Yam, W.K.; Alikhan, N.F.; Petty, N.K.; Sartor, A.L.; Williamson, D.A.; Forde, B.M.; Schembri, M.A.; Beatson, S.A.; et al. Mechanisms involved in acquisition of blaNDM genes by IncA/C2 and IncFIIY plasmids. Antimicrob. Agents Chemother. 2016, 60, 4082–4088. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.M.; Zhong, L.L.; Zhang, X.F.; Hu, H.T.; Li, Y.Q.; Yang, X.R.; Feng, L.Q.; Huang, X.; Tian, G.B. NDM-1-producing Citrobacter freundii, Escherichia coli, and Acinetobacter baumannii identified from a single patient in China. Antimicrob. Agents Chemother. 2015, 59, 5073–5077. [Google Scholar] [CrossRef] [PubMed]
- Pillonetto, M.; Arend, L.; Vespero, E.C.; Pelisson, M.; Chagas, T.P.; Carvalho-Assef, A.P.; Asensi, M.D. First report of NDM-1-producing Acinetobacter baumannii sequence type 25 in Brazil. Antimicrob. Agents Chemother. 2014, 58, 7592–7594. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Schrenzel, J.; Cherkaoui, A.; Bernabeu, S.; Renzi, G.; Nordmann, P. Molecular analysis of NDM-1-producing enterobacterial isolates from Geneva, Switzerland. J. Antimicrob. Chemother. 2011, 66, 1730–1733. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Lu, Z.; Schwarz, S.; Zhang, R.M.; Wang, X.M.; Si, W.; Yu, S.; Chen, L.; Liu, S. Complete sequence of the blaNDM-1-carrying plasmid pNDM-AB from Acinetobacter baumannii of food animal origin. J. Antimicrob. Chemother. 2013, 68, 1681–1682. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, R.A.; Poirel, L.; Naas, T.; Pirs, M.; Seme, K.; Schrenzel, J.; Nordmann, P. Dissemination of New Delhi metallo-beta-lactamase-1-producing Acinetobacter baumannii in Europe. Clin. Microbiol. Infect. 2012, 18, E362–E365. [Google Scholar] [CrossRef] [PubMed]
- Decousser, J.W.; Jansen, C.; Nordmann, P.; Emirian, A.; Bonnin, R.A.; Anais, L.; Merle, J.C.; Poirel, L. Outbreak of NDM-1-producing Acinetobacter baumannii in France, January to May 2013. Euro Surveill. 2013, 18, 2–5. [Google Scholar] [CrossRef]
- Poirel, L.; Bonnin, R.A.; Boulanger, A.; Schrenzel, J.; Kaase, M.; Nordmann, P. Tn125-related acquisition of blaNDM-like genes in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2012, 56, 1087–1089. [Google Scholar] [CrossRef] [PubMed]
- Krahn, T.; Wibberg, D.; Maus, I.; Winkler, A.; Bontron, S.; Sczyrba, A.; Nordmann, P.; Pühler, A.; Poirel, L.; Schlüter, A. Intraspecies transfer of the chromosomally encoded Acinetobacter baumannii blaNDM-1 carbapenemase gene. Antimicrob. Agents Chemother. 2016, 60, 3032–3040. [Google Scholar] [CrossRef] [PubMed]
- Martinez, T.; Vazquez, G.J.; Aquino, E.E.; Martinez, I.; Robledo, I.E. ISEcp1-mediated transposition of blaKPC into the chromosome of a clinical isolate of Acinetobacter baumannii from Puerto Rico. J. Med. Microbiol. 2014, 63, 1644–1648. [Google Scholar] [CrossRef] [PubMed]
- Martinez, T.; Martinez, I.; Vazquez, G.J.; Aquino, E.E.; Robledo, I.E. Genetic environment of the KPC gene in Acinetobacter baumannii ST2 clone from Puerto Rico and genomic insights into its drug resistance. J. Med. Microbiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Moubareck, C.; Bremont, S.; Conroy, M.C.; Courvalin, P.; Lambert, T. GES-11, a novel integron-associated GES variant in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 3579–3581. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, R.A.; Rotimi, V.O.; Al Hubail, M.; Gasiorowski, E.; Al Sweih, N.; Nordmann, P.; Poirel, L. Wide dissemination of GES-type carbapenemases in Acinetobacter baumannii isolates in Kuwait. Antimicrob. Agents Chemother. 2013, 57, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Bogaerts, P.; Naas, T.; El Garch, F.; Cuzon, G.; Deplano, A.; Delaire, T.; Huang, T.D.; Lissoir, B.; Nordmann, P.; Glupczynski, Y. GES extended-spectrum beta-lactamases in Acinetobacter baumannii isolates in Belgium. Antimicrob. Agents Chemother. 2010, 54, 4872–4878. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, R.A.; Nordmann, P.; Potron, A.; Lecuyer, H.; Zahar, J.R.; Poirel, L. Carbapenem-hydrolyzing GES-type extended-spectrum beta-lactamase in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2011, 55, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Karah, N.; Giske, C.G.; Sundsfjord, A.; Samuelsen, O. A diversity of OXA-carbapenemases and class 1 integrons among carbapenem-resistant Acinetobacter baumannii clinical isolates from Sweden belonging to different international clonal lineages. Microb. Drug Resist. 2011, 17, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Charfi-Kessis, K.; Mansour, W.; Ben Haj Khalifa, A.; Mastouri, M.; Nordmann, P.; Aouni, M.; Poirel, L. Multidrug-resistant Acinetobacter baumannii strains carrying the blaoxa-23 and the blaGES-11 genes in a neonatology center in Tunisia. Microb. Pathog. 2014, 74, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Marque, S.; Heritier, C.; Segonds, C.; Chabanon, G.; Nordmann, P. OXA-58, a novel class D beta-lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2005, 49, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Figueiredo, S.; Cattoir, V.; Carattoli, A.; Nordmann, P. Acinetobacter radioresistens as a silent source of carbapenem resistance for Acinetobacter spp. Antimicrob. Agents Chemother. 2008, 52, 1252–1256. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Du, X.; Wang, L.; Yang, Q.; Fu, Y.; Yu, Y. Clinical carbapenem-resistant Acinetobacter baylyi strain coharboring blaSIM-1 and blaOXA-23 from China. Antimicrob. Agents Chemother. 2011, 55, 5347–5349. [Google Scholar] [CrossRef] [PubMed]
- Boo, T.W.; Walsh, F.; Crowley, B. Molecular characterization of carbapenem-resistant Acinetobacter species in an Irish university hospital: Predominance of Acinetobacter genomic species 3. J. Med. Microbiol. 2009, 58, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Smet, A.; Cools, P.; Krizova, L.; Maixnerova, M.; Sedo, O.; Haesebrouck, F.; Kempf, M.; Nemec, A.; Vaneechoutte, M. Acinetobacter gandensis sp. nov. isolated from horse and cattle. Int. J. Syst. Evol. Microbiol. 2014, 64, 4007–4015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zander, E.; Fernandez-Gonzalez, A.; Schleicher, X.; Dammhayn, C.; Kamolvit, W.; Seifert, H.; Higgins, P.G. Worldwide dissemination of acquired carbapenem-hydrolysing class D beta-lactamases in Acinetobacter spp. other than Acinetobacter baumannii. Int. J. Antimicrob. Agents 2014, 43, 375–377. [Google Scholar] [CrossRef] [PubMed]
- Adams-Haduch, J.M.; Paterson, D.L.; Sidjabat, H.E.; Pasculle, A.W.; Potoski, B.A.; Muto, C.A.; Harrison, L.H.; Doi, Y. Genetic basis of multidrug resistance in Acinetobacter baumannii clinical isolates at a tertiary medical center in Pennsylvania. Antimicrob. Agents Chemother. 2008, 52, 3837–3843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La, M.V.; Jureen, R.; Lin, R.T.; Teo, J.W. Unusual detection of an Acinetobacter class D carbapenemase gene, blaOXA-23, in a clinical Escherichia coli isolate. J. Clin. Microbiol. 2014, 52, 3822–3823. [Google Scholar] [CrossRef] [PubMed]
- Osterblad, M.; Karah, N.; Halkilahti, J.; Sarkkinen, H.; Uhlin, B.E.; Jalava, J. Rare detection of the Acinetobacter class D carbapenemase blaOXA-23 gene in Proteus mirabilis. Antimicrob. Agents Chemother. 2016, 60, 3243–3245. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, R.; Marchandin, H.; Chanal, C.; Sirot, D.; Labia, R.; de Champs, C.; Jumas-Bilak, E.; Sirot, J. Chromosome-encoded class D beta-lactamase OXA-23 in Proteus mirabilis. Antimicrob. Agents Chemother. 2002, 46, 2004–2006. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.L.; Ji, S.J.; Ruan, Z.; Fu, Y.; Fu, Y.Q.; Wang, Y.F.; Yu, Y.S. Dissemination of blaOXA-23 in Acinetobacter spp. in China: Main roles of conjugative plasmid pAZJ221 and transposon Tn2009. Antimicrob. Agents Chemother. 2015, 59, 1998–2005. [Google Scholar] [CrossRef] [PubMed]
- Bou, G.; Oliver, A.; Martínez-Beltrán, J. OXA-24, a novel class D β-lactamase with carbapenemase activity in an Acinetobacter baumannii clinical strain. Antimicrob. Agents Chemother. 2006, 50, 2280. [Google Scholar] [CrossRef]
- Mosqueda, N.; Gato, E.; Roca, I.; Lopez, M.; de Alegria, C.R.; Fernandez Cuenca, F.; Martinez-Martinez, L.; Pachon, J.; Cisneros, J.M.; Rodriguez-Bano, J.; et al. Characterization of plasmids carrying the blaOXA-24/40 carbapenemase gene and the genes encoding the AbkA/AbkB proteins of a toxin/antitoxin system. J. Antimicrob. Chemother. 2014, 69, 2629–2633. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Kim, M.N.; Choi, T.Y.; Cho, S.E.; Lee, S.; Whang, D.H.; Yong, D.; Chong, Y.; Woodford, N.; Livermore, D.M.; et al. Wide dissemination of OXA-type carbapenemases in clinical Acinetobacter spp. isolates from South Korea. Int. J. Antimicrob. Agents 2009, 33, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Quinteira, S.; Grosso, F.; Ramos, H.; Peixe, L. Molecular epidemiology of imipenem-resistant Acinetobacter haemolyticus and Acinetobacter baumannii isolates carrying plasmid-mediated OXA-40 from a Portuguese hospital. Antimicrob. Agents Chemother. 2007, 51, 3465–3466. [Google Scholar] [CrossRef] [PubMed]
- Sevillano, E.; Gallego, L.; Garcia-Lobo, J.M. First detection of the OXA-40 carbapenemase in P. aeruginosa isolates, located on a plasmid also found in A. baumannii. Pathol. Biol. (Paris) 2009, 57, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, M.J.; Kesty, N.C. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes Dev. 2005, 19, 2645–2655. [Google Scholar] [CrossRef] [PubMed]
- Boo, T.W.; Crowley, B. Detection of blaOXA-58 and blaOXA-23-like genes in carbapenem-susceptible Acinetobacter clinical isolates: Should we be concerned? J. Med. Microbiol. 2009, 58, 839–841. [Google Scholar] [CrossRef] [PubMed]
- Bosnjak, Z.; Plecko, V.; Budimir, A.; Marekovic, I.; Bedenic, B. First report of NDM-1-producing Acinetobacter guillouiae. Chemotherapy 2014, 60, 250–252. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Yang, P.; Wang, X.; Zong, Z. Characterization of Acinetobacter johnsonii isolate XBB1 carrying nine plasmids and encoding NDM-1, OXA-58 and PER-1 by genome sequencing. J. Antimicrob. Chemother. 2016, 71, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Marti, S.; Sanchez-Cespedes, J.; Blasco, M.D.; Espinal, P.; Ruiz, M.; Alba, V.; Vila, J. Characterization of the carbapenem-hydrolyzing oxacillinase OXA-58 in an Acinetobacter phenon 6/ct13TU clinical isolate. Diagn. Microbiol. Infect. Dis. 2008, 61, 468–470. [Google Scholar] [CrossRef] [PubMed]
- Marti, S.; Sanchez-Cespedes, J.; Blasco, M.D.; Ruiz, M.; Espinal, P.; Alba, V.; Fernandez-Cuenca, F.; Pascual, A.; Vila, J. Characterization of the carbapenem-hydrolyzing oxacillinase OXA-58 in an Acinetobacter genospecies 3 clinical isolate. Antimicrob. Agents Chemother. 2008, 52, 2955–2958. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.E.; Bell, J.M.; Turnidge, J.D.; Castanheira, M.; Deshpande, L.M.; Jones, R.N. Codetection of blaOXA-23-like gene (blaOXA-133) and blaOXA-58 in Acinetobacter radioresistens: Report from the SENTRY antimicrobial surveillance program. Antimicrob. Agents Chemother. 2009, 53, 843–844. [Google Scholar] [CrossRef] [PubMed]
- Cayo, R.; Rodrigues-Costa, F.; Pereira Matos, A.; Godoy Carvalhaes, C.; Dijkshoorn, L.; Gales, A.C. Old clinical isolates of Acinetobacter seifertii in Brazil producing OXA-58. Antimicrob. Agents Chemother. 2016, 60, 2589–2591. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Chen, X.; Meng, X.; Zhang, G.; Wang, J.; Zhou, D.; Guo, X. “Roar” of blaNDM-1 and “silence” of blaOXA-58 co-exist in Acinetobacter pittii. Sci. Rep. 2015, 5, 8976. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.T.; Kuo, S.C.; Chiang, M.H.; Lee, Y.T.; Sung, W.C.; Chen, Y.H.; Chen, T.L.; Fung, C.P. Acinetobacter baumannii extracellular OXA-58 is primarily and selectively released via outer membrane vesicles after Sec-dependent periplasmic translocation. Antimicrob. Agents Chemother. 2015, 59, 7346–7354. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.T.; Kuo, S.C.; Lee, Y.T.; Chen, C.P.; Lin, S.W.; Shen, L.J.; Fung, C.P.; Cho, W.L.; Chen, T.L. Sheltering effect and indirect pathogenesis of carbapenem-resistant Acinetobacter baumannii in polymicrobial infection. Antimicrob. Agents Chemother. 2014, 58, 3983–3990. [Google Scholar] [CrossRef] [PubMed]
- Antonio, C.S.; Neves, P.R.; Medeiros, M.; Mamizuka, E.M.; Elmor de Araujo, M.R.; Lincopan, N. High prevalence of carbapenem-resistant Acinetobacter baumannii carrying the blaOXA-143 gene in Brazilian hospitals. Antimicrob. Agents Chemother. 2011, 55, 1322–1323. [Google Scholar] [CrossRef] [PubMed]
- Carvalhaes, C.G.; Cayo, R.; Assis, D.M.; Martins, E.R.; Juliano, L.; Juliano, M.A.; Gales, A.C. Detection of SPM-1-producing Pseudomonas aeruginosa and class D beta-lactamase-producing Acinetobacter baumannii isolates by use of liquid chromatography-mass spectrometry and matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2013, 51, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Leite, G.C.; Oliveira, M.S.; Perdigao-Neto, L.V.; Rocha, C.K.; Guimaraes, T.; Rizek, C.; Levin, A.S.; Costa, S.F. Antimicrobial combinations against pan-resistant Acinetobacter baumannii isolates with different resistance mechanisms. PLoS ONE 2016, 11, e0151270. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Nagao, M.; Matsumura, Y.; Hotta, G.; Matsushima, A.; Ito, Y.; Takakura, S.; Ichiyama, S. Regional dissemination of Acinetobacter species harbouring metallo-beta-lactamase genes in Japan. Clin. Microbiol. Infect. 2013, 19, 729–736. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information—Beta-Lactamase Data Resources. Available online: http://www.ncbi.nlm.nih.gov/pathogens/beta-lactamase-data-resources/ (accessed on 23 May 2016).
- Espedido, B.A.; Partridge, S.R.; Iredell, J.R. blaIMP-4 in different genetic contexts in Enterobacteriaceae isolates from Australia. Antimicrob. Agents Chemother. 2008, 52, 2984–2987. [Google Scholar] [CrossRef] [PubMed]
- Grosso, F.; Silva, L.; Sousa, C.; Ramos, H.; Quinteira, S.; Peixe, L. Extending the reservoir of blaIMP-5: The emerging pathogen Acinetobacter bereziniae. Future Microbiol. 2015, 10, 1609–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, A.; Yomoda, S.; Kobayashi, I.; Okubo, T.; Tsunoda, M.; Iyobe, S. Detection of carbapenemase-producing Acinetobacter baumannii in a hospital. J. Clin. Microbiol. 2000, 38, 526–529. [Google Scholar] [PubMed]
- Zhao, W.H.; Hu, Z.Q. Epidemiology and genetics of VIM-type metallo-beta-lactamases in Gram-negative bacilli. Future Microbiol. 2011, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- INTEGRALL—The Integron Database. Available online: http://integrall.bio.ua.pt/ (accessed on 23 May 2016).
- Oh, E.J.; Lee, S.; Park, Y.J.; Park, J.J.; Park, K.; Kim, S.I.; Kang, M.W.; Kim, B.K. Prevalence of metallo-beta-lactamase among Pseudomonas aeruginosa and Acinetobacter baumannii in a Korean university hospital and comparison of screening methods for detecting metallo-beta-lactamase. J. Microbiol. Methods 2003, 54, 411–418. [Google Scholar] [CrossRef]
- Lee, M.F.; Peng, C.F.; Hsu, H.J.; Chen, Y.H. Molecular characterisation of the metallo-beta-lactamase genes in imipenem-resistant Gram-negative bacteria from a university hospital in southern Taiwan. Int. J. Antimicrob. Agents 2008, 32, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, Y.; Wilharm, G.; Zander, E.; Wichelhaus, T.A.; Gottig, S.; Hunfeld, K.P.; Seifert, H.; Witte, W.; Higgins, P.G. Molecular characterization of blaNDM-1 in an Acinetobacter baumannii strain isolated in Germany in 2007. J. Antimicrob. Chemother. 2011, 66, 1998–2001. [Google Scholar] [CrossRef] [PubMed]
- Espinal, P.; Poirel, L.; Carmeli, Y.; Kaase, M.; Pal, T.; Nordmann, P.; Vila, J. Spread of NDM-2-producing Acinetobacter baumannii in the Middle East. J. Antimicrob. Chemother. 2013, 68, 1928–1930. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; D’Souza, R.; Pinto, N.; Ryu, C.M.; Park, J.; Yong, D.; Lee, K. Characterization and complete genome sequence analysis of two Myoviral bacteriophages infecting clinical carbapenem-resistant Acinetobacter baumannii isolates. J. Appl. Microbiol. 2016, 121, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Robledo, I.E.; Aquino, E.E.; Sante, M.I.; Santana, J.L.; Otero, D.M.; Leon, C.F.; Vazquez, G.J. Detection of KPC in Acinetobacter spp. in Puerto Rico. Antimicrob. Agents Chemother. 2010, 54, 1354–1357. [Google Scholar] [CrossRef] [PubMed]
- Robledo, I.E.; Aquino, E.E.; Vazquez, G.J. Detection of the KPC gene in Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Acinetobacter baumannii during a PCR-based nosocomial surveillance study in Puerto Rico. Antimicrob. Agents Chemother. 2011, 55, 2968–2970. [Google Scholar] [CrossRef] [PubMed]
- Azimi, L.; Talebi, M.; Pourshafie, M.R.; Owlia, P.; Rastegar Lari, A. Characterization of carbapenemases in extensively drug resistance Acinetobacter baumannii in a Burn Care Center in Iran. Int. J. Mol. Cell. Med. 2015, 4, 46–53. [Google Scholar] [PubMed]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile beta-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [PubMed]
- Al-Agamy, M.H.; Khalaf, N.G.; Tawfick, M.M.; Shibl, A.M.; El Kholy, A. Molecular characterization of carbapenem-insensitive Acinetobacter baumannii in Egypt. Int. J. Infect. Dis. 2014, 22, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Cicek, A.C.; Saral, A.; Iraz, M.; Ceylan, A.; Duzgun, A.O.; Peleg, A.Y.; Sandalli, C. OXA- and GES-type beta-lactamases predominate in extensively drug-resistant Acinetobacter baumannii isolates from a Turkish University Hospital. Clin. Microbiol. Infect. 2014, 20, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Hammoudi, D.; Moubareck, C.A.; Hakime, N.; Houmani, M.; Barakat, A.; Najjar, Z.; Suleiman, M.; Fayad, N.; Sarraf, R.; Sarkis, D.K. Spread of imipenem-resistant Acinetobacter baumannii co-expressing OXA-23 and GES-11 carbapenemases in Lebanon. Int. J. Infect. Dis. 2015, 36, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Nordmann, P. Carbapenem resistance in Acinetobacter baumannii: Mechanisms and epidemiology. Clin. Microbiol. Infect. 2006, 12, 826–836. [Google Scholar] [CrossRef] [PubMed]
- Canchaya, C.; Fournous, G.; Brussow, H. The impact of prophages on bacterial chromosomes. Mol. Microbiol. 2004, 53, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Brussow, H.; Canchaya, C.; Hardt, W.D. Phages and the evolution of bacterial pathogens: From genomic rearrangements to lysogenic conversion. Microbiol. Mol. Biol. Rev. 2004, 68, 560–602. [Google Scholar] [CrossRef] [PubMed]
- Brabban, A.D.; Hite, E.; Callaway, T.R. Evolution of foodborne pathogens via temperate bacteriophage-mediated gene transfer. Foodb. Pathog. Dis. 2005, 2, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Novick, R.P. Phage-mediated intergeneric transfer of toxin genes. Science 2009, 323, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.M.; Ray, J.L.; Johnsen, P.J. Horizontal gene transfer: Uptake of extracellular DNA by bacteria. In Encyclopedia of Microbiology; Shaechter, M., Ed.; Elsevier: Oxford, UK, 2009; pp. 587–596. [Google Scholar]
- Stokes, H.W.; Gillings, M.R. Gene flow, mobile genetic elements and the recruitment of antibiotic resistance genes into Gram-negative pathogens. FEMS Microbiol. Rev. 2011, 35, 790–819. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.; da Silva, G.J.; Nielsen, K.M. Global dissemination patterns of common gene cassette arrays in class 1 integrons. Microbiology 2015, 161, 1313–1337. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, M.D.; Harding, C.M.; Baker, B.D.; Bonomo, R.A.; Hujer, K.M.; Rather, P.N.; Munson, R.S., Jr. Draft genome sequence of the clinical isolate Acinetobacter nosocomialis strain M2. Genome Announc. 2013, 1, e00906-13. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.M.; Tracy, E.N.; Carruthers, M.D.; Rather, P.N.; Actis, L.A.; Munson, R.S., Jr. Acinetobacter baumannii strain M2 produces type IV pili which play a role in natural transformation and twitching motility but not surface-associated motility. MBio 2013, 4, e00360–e00313. [Google Scholar] [CrossRef] [PubMed]
- Johnsborg, O.; Eldholm, V.; Havarstein, L.S. Natural genetic transformation: Prevalence, mechanisms and function. Res. Microbiol. 2007, 158, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Fulsundar, S.; Harms, K.; Flaten, G.E.; Johnsen, P.J.; Chopade, B.A.; Nielsen, K.M. Gene transfer potential of outer membrane vesicles of Acinetobacter baylyi and effects of stress on vesiculation. Appl. Environ. Microbiol. 2014, 80, 3469–3483. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Carbapenemases | bla Gene Type | Resistance Origin | Location | Intercelullar Transfer (HGT) 1 | Genetic Context | Intracellular Movement | References |
|---|---|---|---|---|---|---|---|
| CHDLs 2 | OXA-51 | Intrinsic | Mainly chromosomal; | Sometimes, ISAba1-blaoxa-51-like | [33,35] | ||
| plasmid | Tn6080 | Transposition | [34] | ||||
| OXA-23 | Acquired | Chromosome and plasmid | Conjugation | Genomic islands; Transposons; Insertion sequences | Transposition | [62,63,64,65,66,67,68,69,70,71,72,73] | |
| OXA-40/24 | Acquired | Chromosome and plasmid | Conjugation? 5 | Self-transmissible plasmid belonging to replicon group GR6 | [45,74,75,76,77] | ||
| and in plasmids containing mob genes | [52,78] | ||||||
| Flanked by conserved inverted repeats homologous to XerC/XerD, recombinases’ targets | Site-specific recombination | [77,78,79,80] | |||||
| OMVs 4-mediated transfer | [59] | ||||||
| OXA-58 | Acquired | Mostly plasmidic | Conjugation? 5 | Self-conjugative plasmid belonging to replicon group GR6 | [52,67,81,82,83] | ||
| Insertion sequences | [3,50,84,85] | ||||||
| Flanked by two repeated sequences, Re27 | Homologous recombination | [84] | |||||
| Flanked by two IS26 leading to duplication of ISAba2/ISAba3-blaOXA-58-ISAba3 | Transposition | [86] | |||||
| OXA-143 | Acquired | Plasmid | Flanked by two rep genes | Homologous recombination | [45] | ||
| XerC/XerD recognition site | Site-specific recombination | [87] | |||||
| OXA-235 | Acquired | Chromosome and plasmid | Flanked by two ISAba1 | Transposition | [46] | ||
| MBLs 3 | IMP | Acquired | Chromosome and plasmid | Conjugation | Many inserted in class 1 integrons | [88,89,90,91,92,93] | |
| Gene not embedded into integrons | [94] | ||||||
| Class 1 integron flanked by MITEs 6 | Transposition | [95,96] | |||||
| Natural transformation | Chromosomal class 1 integron | Homologous recombination | [89,97] | ||||
| VIM | Acquired | Chromosome | Class 1 and 2 integrons | [98,99,100,101,102], GenBank Ac. nr. LC107606 | |||
| NDM | Acquired | Mostly plasmidic; | Conjugation | Associated to composite transposon Tn125 | Transposition | [103,104,105,106,107,108,109,110] | |
| chromosome | Transduction | Flanked by ISAba125 and ISCR21 | [111] | ||||
| Class A β-lactamases | KPC | Acquired | Chromosome | Tn4401b embedded into KQ-like element | Transposition | [112] | |
| (Novel truncated version of) Tn4401e | [113] | ||||||
| GES | Acquired | Chromosome and plasmid | Conjugation | Class 1 integrons | [114,115,116,117,118,119] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Da Silva, G.J.; Domingues, S. Insights on the Horizontal Gene Transfer of Carbapenemase Determinants in the Opportunistic Pathogen Acinetobacter baumannii. Microorganisms 2016, 4, 29. https://doi.org/10.3390/microorganisms4030029
Da Silva GJ, Domingues S. Insights on the Horizontal Gene Transfer of Carbapenemase Determinants in the Opportunistic Pathogen Acinetobacter baumannii. Microorganisms. 2016; 4(3):29. https://doi.org/10.3390/microorganisms4030029
Chicago/Turabian StyleDa Silva, Gabriela Jorge, and Sara Domingues. 2016. "Insights on the Horizontal Gene Transfer of Carbapenemase Determinants in the Opportunistic Pathogen Acinetobacter baumannii" Microorganisms 4, no. 3: 29. https://doi.org/10.3390/microorganisms4030029