Rapid and Highly Sensitive Non-Competitive Immunoassay for Specific Detection of Nodularin

Abstract
:1. Introduction
2. Materials and Methods
2.1. Common Materials and Instruments
2.2. Toxin Standards
2.3. Affinity Selection of Anti-Immunocomplex NOD-Specific Phage Antibodies
2.4. Cloning, Expression, Screening and Purification
2.5. The Non-Competitive NOD-Specific Assay
2.6. Effect of Incubation Time and Total Assay Volume
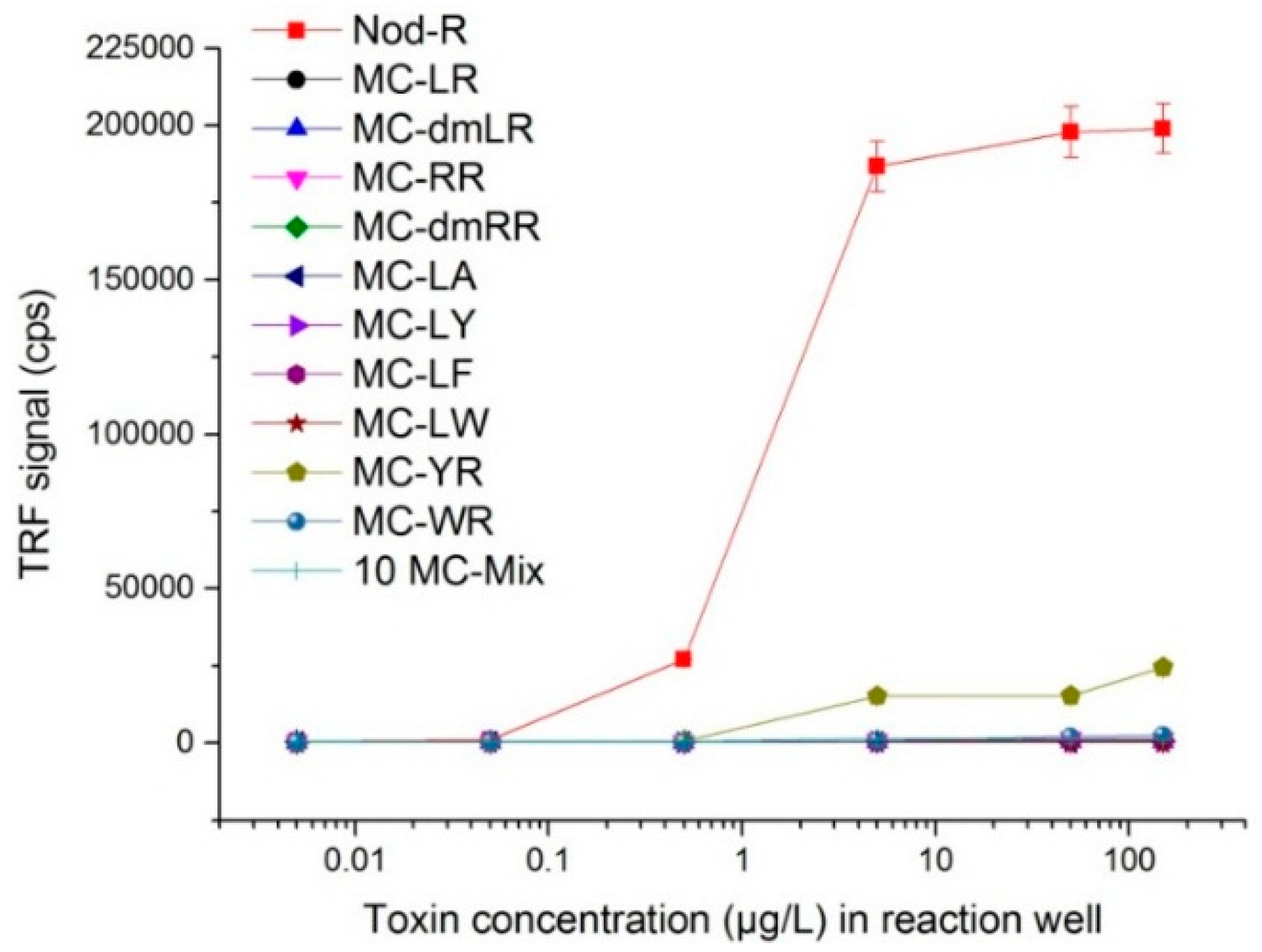
2.7. Analysis of the Specificity Profile of SA32C11
2.8. Sample Analysis
2.8.1. Recovery of NOD from Spiked Water Samples
2.8.2. Raw Water Samples with Known Toxin Content
2.8.3. Sea and Coastal Inlet Water
2.8.4. Bloom Samples from the Sample Archive
2.8.5. Tissue Samples
2.8.6. Extraction of Hepatotoxins
3. Results
3.1. Immunocomplex Panning
3.2. Screening Summary
3.3. Specificity Profile of SA32C11 scFv-AP
3.4. Optimization of the Assay Time
3.5. Assay Performance at Different Volumes
3.6. Sample Analysis
3.6.1. Recovery of NOD from Spiked Water Samples
3.6.2. Water Samples with Known Toxin Content
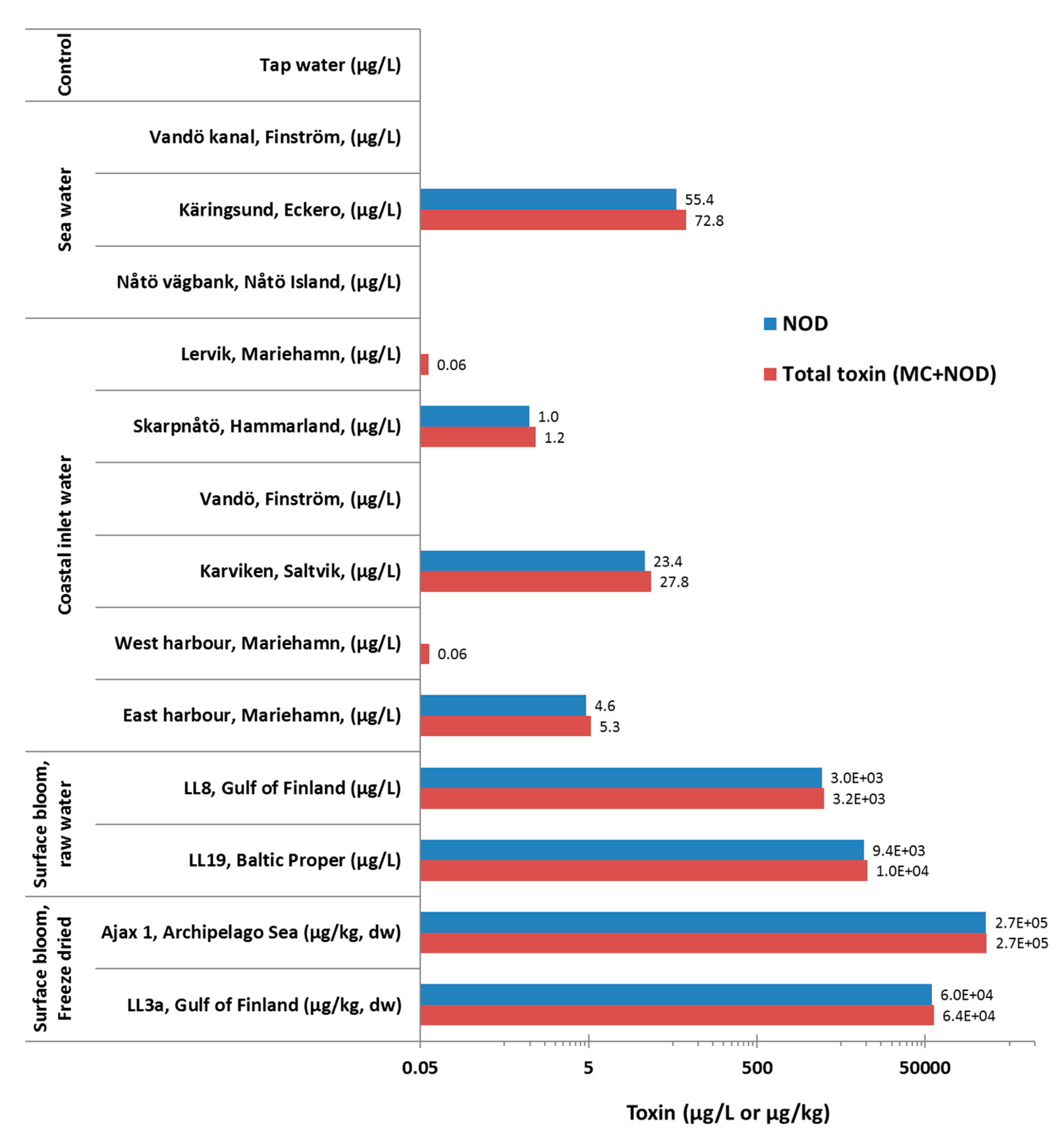
3.6.3. Samples with Unknown Toxin Content
Sea and Coastal Water Samples
Surface Bloom Samples
Tissue Sample
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Sipiä, V.; Kankaanpää, H.; Lahti, K.; Carmichael, W.W.; Meriluoto, J. Detection of nodularin in flounders and cod from the baltic sea. Environ. Toxicol. 2001, 16, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Sipiä, V.; Kankaanpää, H.; Peltonen, H.; Vinni, M.; Meriluoto, J. Transfer of nodularin to three-spined stickleback (Gasterosteus aculeatus L.), herring (Clupea harengus L.), and salmon (Salmo salar L.) in the northern baltic sea. Ecotoxicol. Environ. Saf. 2007, 66, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Sipiä, V.O.; Kankaanpää, H.T.; Flinkman, J.; Lahti, K.; Meriluoto, J.A.O. Time-dependent accumulation of cyanobacterial hepatotoxins in flounders (Platichthys flesus) and mussels (Mytilus edulis) from the northern baltic sea. Environ. Toxicol. 2001, 16, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Sipiä, V.O.; Kankaanpää, H.T.; Pflugmacher, S.; Flinkman, J.; Furey, A.; James, K.J. Bioaccumulation and detoxication of nodularin in tissues of flounder (Platichthys flesus), mussels (Mytilus edulis, Dreissena polymorpha), and clams (Macoma balthica) from the northern baltic sea. Ecotoxicol. Environ. Saf. 2002, 53, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Francis, G. Poisonous australia lake. Nature 1878, 18, 11–12. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Harada, K.; Namikoshi, M.; Chen, C.; Harvis, C.A.; Munro, M.H.G.; Blunt, J.W.; Mulligan, P.E.; Beasley, V.R.; Dahlem, A.M.; et al. Nodularin, microcystin, and the configuration of adda. J. Am. Chem. Soc. 1988, 110, 8557–8558. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Eschedor, J.T.; Patterson, G.M.; Moore, R.E. Toxicity and partial structure of a hepatotoxic peptide produced by the cyanobacterium nodularia spumigena mertens emend. L575 from New Zealand. Appl. Environ. Microbiol. 1988, 54, 2257–2263. [Google Scholar] [PubMed]
- Honkanen, R.E.; Dukelow, M.; Zwiller, J.; Moore, R.E.; Khatra, B.S.; Boynton, A.L. Cyanobacterial nodularin is a potent inhibitor of type 1 and type 2a protein phosphatases. Mol. Pharmacol. 1991, 40, 577–583. [Google Scholar] [PubMed]
- Eriksson, J.E.; Meriluoto, J.A.; Kujari, H.P.; Osterlund, K.; Fagerlund, K.; Hallbom, L. Preliminary characterization of a toxin isolated from the cyanobacterium nodularia spumigena. Toxicon 1988, 26, 161–166. [Google Scholar] [CrossRef]
- Yoshizawa, S.; Matsushima, R.; Watanabe, M.F.; Harada, K.; Ichihara, A.; Carmichael, W.W.; Fujiki, H. Inhibition of protein phosphatases by microcystis and nodularin associated with hepatotoxicity. J. Cancer Res. Clin. Oncol. 1990, 116, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Harding, W.R.; Rowe, N.; Wessels, J.C.; Beattie, K.A.; Codd, G.A. Death of a dog attributed to the cyanobacterial (blue-green algal) hepatotoxin nodularin in south africa. J. S. Afr. Vet. Assoc. 1995, 66, 256–259. [Google Scholar] [PubMed]
- Sueoka, E.; Sueoka, N.; Okabe, S.; Kozu, T.; Komori, A.; Ohta, T.; Suganuma, M.; Kim, S.J.; Lim, I.K.; Fujiki, H. Expression of the tumor necrosis factorα gene and early response genes by nodularin, a liver tumor promoter, in primary cultured rat hepatocytes. J. Cancer Res. Clin. Oncol. 1997, 123, 413–419. [Google Scholar] [PubMed]
- Žegura, B.; Štraser, A.; Filipič, M. Genotoxicity and potential carcinogenicity of cyanobacterial toxins—A review. Mutat. Res. 2011, 727, 16–41. [Google Scholar] [CrossRef] [PubMed]
- Lankoff, A.; Banasik, A.; Nowak, M. Protective effect of melatonin against nodularin-induced oxidative stress in mouse liver. Arch. Toxicol. 2002, 76, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Maatouk, I.; Bouaïcha, N.; Plessis, M.J.; Périn, F. Detection by 32p-postlabelling of 8-oxo-7,8-dihydro-2′-deoxyguanosine in DNA as biomarker of microcystin-LR- and nodularin-induced DNA damage in vitro in primary cultured rat hepatocytes and in vivo in rat liver. Mutat. Res. 2004, 564, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Pflugmacher, S.; Olin, M.; Kankaanpää, H. Nodularin induces oxidative stress in the baltic sea brown alga fucus vesiculosus (phaeophyceae). Mar. Environ. Res. 2007, 64, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Lehtimäki, N.; Shunmugam, S.; Jokela, J.; Wahlsten, M.; Carmel, D.; Keränen, M.; Sivonen, K.; Aro, E.-M.; Allahverdiyeva, Y.; Mulo, P. Nodularin uptake and induction of oxidative stress in spinach (Spinachia oleracea). J. Plant Physiol. 2011, 168, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Persson, K.-J.; Legrand, C.; Olsson, T. Detection of nodularin in european flounder (Platichthys flesus) in the west coast of sweden: Evidence of nodularin mediated oxidative stress. Harmful Algae 2009, 8, 832–838. [Google Scholar] [CrossRef]
- Zhang, H.; Shao, D.; Wu, Y.; Cai, C.; Hu, C.; Shou, X.; Dai, B.; Ye, B.; Wang, M.; Jia, X. Apoptotic responses of carassius auratus lymphocytes to nodularin exposure in vitro. Fish Shellfish Immunol. 2012, 33, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shao, D.; Wu, Y.; Dai, B.; Cai, C.; Fang, W.; Ye, B.; Zhang, Y.; liu, J.; Jia, X. Regulation of nodularin-induced apoptosis by epigallocatechin-3-gallate on fish lymphocytes in vitro. Fish Shellfish Immunol. 2013, 34, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Spoof, L.; Catherine, A. Appendix 3. Tables of microcystins and nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; Wiley & Sons: Chichester, UK, 2017; pp. 526–537. [Google Scholar]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef] [PubMed]
- Karjalainen, M.; Pääkkönen, J.-P.; Peltonen, H.; Sipiä, V.; Valtonen, T.; Viitasalo, M. Nodularin concentrations in baltic sea zooplankton and fish during a cyanobacterial bloom. Mar. Biol. 2008, 155, 483–491. [Google Scholar] [CrossRef]
- Karjalainen, M.; Reinikainen, M.; Spoof, L.; Meriluoto, J.A.O.; Sivonen, K.; Viitasalo, M. Trophic transfer of cyanobacterial toxins from zooplankton to planktivores: Consequences for pike larvae and mysid shrimps. Environ. Toxicol. 2005, 20, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Akcaalan, R.; Mazur-Marzec, H.; Zalewska, A.; Albay, M. Phenotypic and toxicological characterization of toxic nodularia spumigena from a freshwater lake in turkey. Harmful Algae 2009, 8, 273–278. [Google Scholar] [CrossRef]
- Gehringer, M.M.; Adler, L.; Roberts, A.A.; Moffitt, M.C.; Mihali, T.K.; Mills, T.J.; Fieker, C.; Neilan, B.A. Nodularin, a cyanobacterial toxin, is synthesized in planta by symbiotic nostoc sp. ISME J. 2012, 6, 1834–1847. [Google Scholar] [CrossRef] [PubMed]
- Beattie, K.A.; Kaya, K.; Codd, G.A. The cyanobacterium nodularia pcc 7804, of freshwater origin, produces [L-Har2] nodularin. Phytochemistry 2000, 54, 57–61. [Google Scholar] [CrossRef]
- Moffitt, M.C.; Neilan, B.A. On the presence of peptide synthetase and polyketide synthase genes in the cyanobacterial genus nodularia. FEMS Microbiol. Lett. 2001, 196, 207–214. [Google Scholar] [CrossRef] [PubMed]
- McGregor, G.B.; Sendall, B.C. Iningainema pulvinus gen nov., sp nov. (Cyanobacteria, Scytonemataceae) a new nodularin producer from Edgbaston Reserve, north-eastern Australia. Harmful Algae 2017, 62, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Falconer, I.R.; Smith, J.V.; Jackson, A.R.B.; Jones, A.; Runnegar, M.T.C. Oral toxicity of a bloom of the cyanobacterium microcystis aeruginosa administered to mice over periods up to 1 year. J. Toxicol. Environ. Health 1988, 24, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Hiskia, A.; Kaloudis, T. Protein phosphatase inhibition assays. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 267–271. [Google Scholar]
- Caixach, J.; Flores, C.; Spoof, L.; Meriluoto, J.; Schmidt, W.; Mazur-Marzec, H.; Hiskia, A.; Kaloudis, T.; Furey, A. Liquid chromatography-mass spectrometry. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 218–257. [Google Scholar]
- Baier, W.; Loleit, M.; Fischer, B.; Jung, G.; Neumann, U.; Weiss, M.; Weckesser, J.; Hoffmann, P.; Bessler, W.G.; Mittenbühler, K. Generation of antibodies directed against the low-immunogenic peptide-toxins microcystin-LR/RR and nodularin. Int. J. Immunopharmacol. 2000, 22, 339–353. [Google Scholar] [CrossRef]
- Mikhailov, A.; Härmälä-Braskén, A.-S.; Polosukhina, E.; Hanski, A.; Wahlsten, M.; Sivonen, K.; Eriksson, J.E. Production and specificity of monoclonal antibodies against nodularin conjugated through n-methyldehydrobutyrine. Toxicon 2001, 39, 1453–1459. [Google Scholar] [CrossRef]
- Ueno, Y.; Nagata, S.; Tsutsumi, T.; Hasegawa, A.; Yoshida, F.; Suttajit, M.; Mebs, D.; Putsch, M.; Vasconcelos, V. Survey of microcystins in environmental water by a highly sensitive immunoassay based on monoclonal antibody. Nat. Toxins 1996, 4, 271–276. [Google Scholar] [CrossRef]
- Nagata, S.; Soutome, H.; Tsutsumi, T.; Hasegawa, A.; Sekijima, M.; Sugamata, M.; Harada, K.; Suganuma, M.; Ueno, Y. Novel monoclonal antibodies against microcystin and their protective activity for hepatotoxicity. Nat. Toxins 1995, 3, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, T.; Nagata, S.; Yoshida, F.; Ueno, Y. Anti-idiotype monoclonal antibodies against anti-microcystin antibody and their use in enzyme immunoassay. Toxicon 1998, 36, 235–245. [Google Scholar] [CrossRef]
- Chu, F.; Huang, X.; Wei, R. Enzyme-linked immunosorbent assay for microcystins in blue-green algal blooms. J. Assoc. Off. Anal. Chem. 1989, 73, 451–456. [Google Scholar]
- Zhou, Y.; Li, Y.-S.; Zhi, B.-H.; Lu, S.-Y.; Ren, H.-L.; Zhang, Y.-Y.; Li, Z.-H.; Shen, Q.-F.; Meng, X.-M.; Liu, Z.-S.; et al. Detection of nodularin based on a monoclonal antibody in water and aquatic fish samples. Food Control 2011, 22, 797–800. [Google Scholar] [CrossRef]
- Akter, S.; Vehniäinen, M.; Spoof, L.; Nybom, S.; Meriluoto, J.; Lamminmäki, U. Broad-spectrum noncompetitive immunocomplex immunoassay for cyanobacterial peptide hepatotoxins (microcystins and nodularins). Anal. Chem. 2016, 88, 10080–10087. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W.; Sambrook, J. The Condensed Protocols from Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2006; 800p. [Google Scholar]
- Meriluoto, J.; Codd, G.A. Toxic: Cyanobacterial Monitoring and Cyanotoxin Analysis; Åbo Akademi University Press: Turku, Finland, 2005. [Google Scholar]
- Brockmann, E.C.; Akter, S.; Savukoski, T.; Huovinen, T.; Lehmusvuori, A.; Leivo, J.; Saavalainen, O.; Azhayev, A.; Lovgren, T.; Hellman, J.; et al. Synthetic single-framework antibody library integrated with rapid affinity maturation by vl shuffling. Protein Eng. Des. Sel. 2011, 24, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Huovinen, T.; Syrjanpaa, M.; Sanmark, H.; Brockmann, E.C.; Azhayev, A.; Wang, Q.; Vehniainen, M.; Lamminmaki, U. Two scfv antibody libraries derived from identical VL-VH framework with different binding site designs display distinct binding profiles. Protein Eng. Des. Sel. 2013, 26, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Savela, H.; Spoof, L.; Perälä, N.; Preede, M.; Lamminmäki, U.; Nybom, S.; Häggqvist, K.; Meriluoto, J.; Vehniäinen, M. Detection of cyanobacterial sxt genes and paralytic shellfish toxins in freshwater lakes and brackish waters on Åland islands, Finland. Harmful Algae 2015, 46, 1–10. [Google Scholar] [CrossRef]
- Savela, H.; Vehniäinen, M.; Spoof, L.; Nybom, S.; Meriluoto, J.; Lamminmäki, U. Rapid quantification of mcyb copy numbers on dry chemistry PCR chips and predictability of microcystin concentrations in freshwater environments. Harmful Algae 2014, 39, 280–286. [Google Scholar] [CrossRef]
- Akter, S.; Vehniäinen, M.; Meriluoto, J.; Spoof, L.; Lamminmäki, U. Non-competitive elisa with broad specificity for microcystins and nodularins. Adv. Oceanogr. Limnol. 2017, 8. [Google Scholar] [CrossRef]
- Kankaanpää, H.T.; Sjövall, O.; Huttunen, M.; Olin, M.; Karlsson, K.; Hyvärinen, K.; Sneitz, L.; Härkönen, J.; Sipiä, V.O.; Meriluoto, J.A.O. Production and sedimentation of peptide toxins nodularin-r and microcystin-LR in the northern baltic sea. Environ. Pollut. 2009, 157, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Lehtimaki, J.; Moisander, P.; Sivonen, K.; Kononen, K. Growth, nitrogen fixation, and nodularin production by two baltic sea cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 1647–1656. [Google Scholar] [PubMed]
- Mazur-Marzec, H.; Krezel, A.; Kobos, J.; Plinski, M. Toxic nodularia spumigena blooms in the coastal waters of the Gulf of Gdańsk: A ten-year survey. Oceanologia 2006, 48, 255–273. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Collection Place | Year | Previous Analysis/Comments | Pretreatment/Sample Processing | |
|---|---|---|---|---|---|
| 1 | Spiked sea waters (n = 2) | Åland Islands, Finland | 2010 | Samples were previously analyzed for cyanobacterial saxitoxin biosynthesis (sxt) genes and paralytic shellfish toxins [46]. No previous analysis of MC/NOD for these samples. | Two seawaters with no detectable toxin were spiked with known amount of NOD before analysis. |
| 2 | Lake waters (n = 13)(MC positive, NOD negative) | Finland and Estonia | 2009 | Toxin content and toxin analogues were detected earlier by LC-MS analysis [41,47,48]. No detectable nodularin, most lakewater contain MCs. | No pretreatment, stored at −20 °C, used in the assay as raw. |
| 3 | Sea waters (n = 2) (NOD positive) | Finland and Estonia | 2009 | Toxin content and toxin analogues were detected earlier by LC-MS analysis [41,47,48]. Both samples contained NOD. | As above |
| 4 | Sea (n = 3) and coastal inlet waters (n = 6) | Åland Islands, Finland | 2010 | Samples were previously analyzed for cyanobacterial saxitoxin biosynthesis (sxt) genes and paralytic shellfish toxins [46]. No previous analysis of MC/NOD for these samples. | As above |
| 5 | Surface bloom containing seawater (n = 2) | Monitoring station LL19 (Baltic Proper) | 2003 | None Cyanobacteria blooms occur in the area during late summer every year | Concentration on a 50 µm plankton net, stored at −20 °C, analyzed after thawing at RT. |
| Monitoring station LL8 (Gulf of Finland) | 2007 | None Sporadic cyanobacteria blooms occur in the area. | No pretreatment. Stored at −20 °C, analyzed after thawing at RT. | ||
| 6 | Surface phytoplankton bloom sample (n = 2) | Monitoring station Ajax 1 (Archipelago Sea) [49] | 2002 | Sporadic cyanobacteria blooms occur in the area. | Freeze-dried and was stored at −20 °C. 25 mg sample was mixed with 5 mL of 70% MeOH. |
| Monitoring station LL3a (Gulf of Finland) | 2009 | As above | Sample was collected using a pump and filtration. Freeze-dried and was stored at −20 °C. 11 mg sample was mixed with 2.2 mL of 70% MeOH. |
| Origin of Water Sample and Date of Collection | NOD-R Added to the Sample (µg/L) | NOD-R Determined by Non-Competitive Assay (µg/L) | CV (%) of the Measurement | Recovery (%) | |
|---|---|---|---|---|---|
| 1 | Reagent water 27.5.2017 | 0 | 0 | - | - |
| 0.1 | 0.08 | 1.68 | 84 | ||
| 0.3 | 0.27 | 2.83 | 91 | ||
| 1 | 0.92 | 2.40 | 92 | ||
| 3 | 2.38 | 4.51 | 79 | ||
| 2 | Drinking tap water 27.5.2017 | 0 | 0 | - | - |
| 0.1 | 0.09 | 4.62 | 90 | ||
| 0.3 | 0.29 | 1.70 | 98 | ||
| 1 | 0.95 | 1.49 | 95 | ||
| 3 | 2.86 | 8.45 | 95 | ||
| 3 | Nåtö vägbank, Nåtö Island (sea) 2.8.2010 | 0 | 0 | - | - |
| 0.1 | 0.11 | 2.22 | 105 | ||
| 0.3 | 0.32 | 1.83 | 107 | ||
| 1 | 1.06 | 1.92 | 106 | ||
| 3 | 3.26 | 5.66 | 109 | ||
| 4 | Vandö, Finström, Coastal inlet 3.8.2010 | 0 | 0 | - | - |
| 0.1 | 0.10 | 1.81 | 98 | ||
| 0.3 | 0.30 | 4.37 | 102 | ||
| 1 | 1.06 | 3.60 | 106 | ||
| 3 | 3.25 | 3.52 | 108 | ||
| 5 | Vandö kanal, Finström, Sea 3.8.2010 | 0 | 0 | - | - |
| 0.1 | 0.11 | 2.16 | 115 | ||
| 0.3 | 0.33 | 1.32 | 109 | ||
| 1 | 1.18 | 1.84 | 118 | ||
| 3 | 3.81 | 1.36 | 127 |
| Place | Date | NOD-Specific Assay | LC-MS Analysis Adapted from [41,47,48] | ||
|---|---|---|---|---|---|
| Toxin Concentration (µg/L) | Toxin Concentration (µg/L) | Toxin Analogues | |||
| 1 | Nåtö vägbank, (Sea), Åland, Finland. | 29.7.2009 | 1.26 | 1.50 | NOD |
| 2 | Stroomi rand (Sea), Estonia. | 18.8.2009 | 0.19 | 0.25 | MC-dmRR, NOD |
| 3 | Brantsböle, Åland, Finland. | 27.7.2009 | 0.08 | 21.40 | MC-dmLR, MC-LR, MC-LY, MC-LW, MC-LF |
| 4 | Hauninen reservoir, Raisio, Finland. | 17.8.2009 | <dl | 0.39 | MC-dmRR, MC-RR, MC-dmLR, 1031,5 |
| 5 | Lemböte byträsk, Lemböte, Åland Islands, Finland. | 29.7.2009 | <dl | 0.32 | MC-YR, MC-dmLR |
| 6 | Lake Peipus, Rannapungerja beach, Estonia. | 25.8.2009 | 0.08 | 0.60 | MC-dmRR, MC-RR, MC-YR, MC-dmLR, MC-LR |
| 7 | Lake Peipus, Mustvee beach, Estonia. | 14.8.2009 | <dl | 0.20 | MC-dmRR, MC-RR, MC-dmLR, MC-LR |
| 8 | Littoistenjärvi, Kaarina, Finland. | 3.9.2013 | <dl | 0.50 | MC-dmRR, MC-RR, MC-YR, MC-dmLR, MC-LR |
| 9 | Littoistenjärvi, Kaarina, Finland. | 11.9.2009 | 0.09 | 3.70 | MC-dmRR, MC-RR, MC-YR, MC-dmLR, MC-LR |
| 10 | Hauninen reservoir, Raisio, Finland. | 14.7.2009 | 0.09 | 0.27 | MC-dmRR |
| 11 | Hauninen reservoir, Raisio, Finland. | 15.9.2009 | <dl | 0.86 | MC-dmRR, MC-RR, MC-dmLR, 1031,5 |
| 12 | Hauninen reservoir, Raisio, Finland. | 29.9.2009 | <dl | 1.90 | MC-dmRR, MC-dmLR, 1031,5 |
| 13 | Hauninen reservoir, Raisio, Finland. | 29.10.2009 | <dl | 0.68 | MC-dmRR, MC-dmLR, 1031,5 |
| 14 | Savojärvi, Pöytyä, Finland. | 7.8.2009 | 0.46 | 40.90 | MC-didmRR, MC-dmRR, MC-didmLR, MC-dmLR |
| 15 | Littoistenjärvi, Kaarina, Finland. | 4.8.2009 | <dl | nd | nd |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akter, S.; Vehniäinen, M.; Kankaanpää, H.T.; Lamminmäki, U. Rapid and Highly Sensitive Non-Competitive Immunoassay for Specific Detection of Nodularin. Microorganisms 2017, 5, 58. https://doi.org/10.3390/microorganisms5030058
Akter S, Vehniäinen M, Kankaanpää HT, Lamminmäki U. Rapid and Highly Sensitive Non-Competitive Immunoassay for Specific Detection of Nodularin. Microorganisms. 2017; 5(3):58. https://doi.org/10.3390/microorganisms5030058
Chicago/Turabian StyleAkter, Sultana, Markus Vehniäinen, Harri T. Kankaanpää, and Urpo Lamminmäki. 2017. "Rapid and Highly Sensitive Non-Competitive Immunoassay for Specific Detection of Nodularin" Microorganisms 5, no. 3: 58. https://doi.org/10.3390/microorganisms5030058
APA StyleAkter, S., Vehniäinen, M., Kankaanpää, H. T., & Lamminmäki, U. (2017). Rapid and Highly Sensitive Non-Competitive Immunoassay for Specific Detection of Nodularin. Microorganisms, 5(3), 58. https://doi.org/10.3390/microorganisms5030058