Transfer RNA Modification Enzymes from Thermophiles and Their Modified Nucleosides in tRNA

 ,
,
Abstract
:1. Introduction
2. Sequencing of tRNA from Thermophiles
3. Modified Nucleosides in tRNA from Moderate Thermophiles Are Common to Those from Mesophiles
4. Strategies of tRNA Stabilization by Modified Nucleosides in Extreme-Thermophiles and Hyper-Thermophiles
4.1. m5s2U54 Is a Typical Thermophile-Specific Modified Nucleoside in tRNA
4.2. The Network Between Modified Nucleosides in tRNA and tRNA Modification Enzymes in T. thermophilus Adapts Protein Synthesis at Low and High Temperatures
4.3. Stabilization of tRNA Structure by 2′-O-Methylation
4.4. Other tRNA Stabilization Factors
5. tRNA Modifications and Environmental Stresses at High Temperatures
5.1. Oxidative Stress
5.2. Other Environmental Stresses
6. Utilization of tRNA Modification Enzymes from Thermophiles
7. Perspective
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Crick, F.H. On protein synthesis. Symp. Soc. Exp. Biol. 1958, 12, 138–163. [Google Scholar] [PubMed]
- Grunberger, D.; Weinstein, I.B.; Jacobson, K.B. Codon recognition by enzymatically mischarged valine transfer ribonucleic acid. Science 1969, 166, 1635–1637. [Google Scholar] [CrossRef] [PubMed]
- Sprinzl, M.; Horn, C.; Brown, M.; Ioudovitch, A.; Steinberg, S. Compilation of tRNA sequences and sequences of tRNA genes. Nucleic Acids Res. 1998, 26, 148–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertus, J.D.; Ladner, J.E.; Finch, J.T.; Rhodes, D.; Brown, R.S.; Clark, B.F.C.; Klug, A. Structure of yeast phenylalanine tRNA at 3 Å resolution. Nature 1974, 250, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Suddath, F.L.; Quigley, G.J.; McPherson, A.; Sussan, J.L.; Wang, A.H.J.; Seeman, N.C.; Rich, A. Three-dimensional tertiary structure of yeast phenylalanine transfer RNA. Science 1974, 185, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Moore, P.B. The crystal structure of yeast phenylalanine tRNA at 1.93 A resolution: A classic structure revisited. RNA 2000, 6, 1091–1105. [Google Scholar] [CrossRef] [PubMed]
- Boccaletto, P.; Machnicka, M.A.; Purta, E.; Piatkowski, P.; Baginski, B.; Wirecki, T.K.; de Crécy-Lagard, V.; Ross, R.; Limbach, P.A.; Kotter, A.; et al. MODOMICS: A database of RNA modification pathways. 2017 update. Nucleic Acids Res 2018, 46, D303–D307. [Google Scholar] [CrossRef] [PubMed]
- Juhling, F.; Morl, M.; Hartmann, R.K.; Sprinzl, M.; Stadler, P.F.; Putz, J. tRNAdb 2009: Compilation of tRNA sequences and tRNA genes. Nucleic Acids Res. 2009, 37, D159–D162. [Google Scholar] [CrossRef] [PubMed]
- Constantinesco, F.; Motorin, Y.; Grosjean, H. Transfer RNA modification enzymes from Pyrococcus furiosus: Detection of the enzymatic activities in vitro. Nucleic Acids Res. 1999, 27, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Grawunder, U.; Schön, A.; Sprintzl, M. Sequence and base modifications of two phenylalanine-tRNAs from Thermus thermophilus HB8. Nucleic Acids Res. 1992, 20, 137. [Google Scholar] [CrossRef] [PubMed]
- Tomikawa, C.; Yokogawa, T.; Kanai, T.; Hori, H. N7-Methylguanine at position 46 (m7G46) in tRNA from Thermus thermophilus is required for cell viability through a tRNA modification network. Nucleic Acids Res. 2010, 38, 942–957. [Google Scholar] [CrossRef] [PubMed]
- Menezes, S.; Gaston, K.W.; Krivos, K.L.; Apolinario, E.E.; Reich, N.O.; Sowers, K.R.; Limbach, P.A.; Perona, J.J. Formation of m2G6 in Methanocaldococcus jannaschii tRNA catalyzed by the novel methyltransferase Trm14. Nucleic Acids Res. 2011, 39, 7641–7655. [Google Scholar] [CrossRef] [PubMed]
- Fislage, M.; Roovers, M.; Münnich, S.; Droogmans, L.; Versées, W. Crystallization and preliminary X-ray crystallographic analysis of putative tRNA-modification enzymes from Pyrococcus furiosus and Thermus thermophilus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 1432–1435. [Google Scholar] [CrossRef] [PubMed]
- Roovers, M.; Oudjama, Y.; Fislage, M.; Bujnicki, J.M.; Versées, W.; Droogmans, L. The open reading frame TTC1157 of Thermus thermophilus HB27 encodes the methyltransferase forming N2-methylguanosine at position 6 in tRNA. RNA 2012, 18, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Fislage, M.; Roovers, M.; Tuszynska, I.; Bujnicki, J.M.; Droogmans, L.; Versées, W. Crystal structures of the tRNA:m2G6 methyltransferase Trm14/TrmN from two domains of life. Nucleic Acids Res. 2012, 40, 5149–5161. [Google Scholar] [CrossRef] [PubMed]
- Randau, L.; Stanley, B.J.; Kohlway, A.; Mechta, S.; Xiong, Y.; Söll, D. A cytidine deaminase edits C to U in transfer RNAs in Archaea. Science 2009, 324, 657–659. [Google Scholar] [CrossRef] [PubMed]
- Su, A.A.; Tripp, V.; Randau, L. RNA-Seq analyses reveal the order of tRNA processing events and the maturation of C/D box and CRISPR RNAs in the hyperthermophile Methanopyrus kandleri. Nucleic Acids Res. 2013, 41, 6250–6258. [Google Scholar] [CrossRef] [PubMed]
- Pixa, G.; Dirheimer, G.; Keith, G. Sequence of tRNALeu CmAA from Bacillus stearothermophilus. Biochem. Biophys. Res. Commun. 1983, 112, 578–585. [Google Scholar] [CrossRef]
- Keith, G.; Guerrier-Takada, C.; Grosjean, H.; Dirheimer, G. A revised sequence for Bacillus stearothermophilus phenylalanine tRNA. FEBS Lett. 1977, 84, 241–243. [Google Scholar] [CrossRef]
- Brown, R.S.; Rubin, J.R.; Rhodes, D.; Guilley, H.; Simoncsits, A.; Brownlee, G.G. The nucleoside sequence of tyrosine tRNA from Bacillus stearothermophilus. Nucleic Acids Res. 1978, 5, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Kuchino, Y.; Yamaizumi, Z.; Kato, M.; Oshima, T.; Nishimura, S. Nucleotide sequence of formylmethionine tRNA from an extreme thermophile, Thermus thermophilus HB8. J. Biochem. 1979, 86, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Oshima, T.; Hansske, F.; Ohta, T. Separation and comparison of 2-thioribothymidine-containing transfer ribonucleic acid and the ribothymidine-containing counterpart from cells of Thermus thermophilus HB 8. Biochemistry 1983, 22, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Keith, G.; Yusupov, M.; Briand, C.; Moras, D.; Kern, D.; Brion, C. Sequence of tRNAAsp from Thermus thermophilus HB8. Nucleic Acids Res. 1993, 21, 4399. [Google Scholar] [CrossRef] [PubMed]
- Awai, T.; Kimura, S.; Tomikawa, C.; Ochi, A.; Ihsanawati, I.S.; Bessho, Y.; Yokoyama, S.; Ohno, S.; Nishikawa, K.; Yokogawa, T.; et al. Aquifex aeolicus tRNA (N2, N2-guanine)-dimethyltransferase (Trm1) catalyzes transfer of methyl groups not only to guanine 26 but also to guanine 27 in tRNA. J. Biol. Chem. 2009, 284, 20467–20478. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, M.W.; Walker, R.T. The nucleotide sequence of the tRNAMMet from the archaebacterium Thermoplasma acidophilum. Nucleic Acids Res. 1981, 9, 4387–4390. [Google Scholar] [CrossRef] [PubMed]
- Caldeira de Araujo, A.; Favre, A. Induction of size reduction in Escherichia coli by near-ultraviolet light. Eur. J. Biochem. 1985, 146, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Kramer, G.F.; Baker, J.C.; Ames, B.N. Near-UV stress in Salmonella typhimurium: 4-thiouridine in tRNA, ppGpp, and ApppGpp as components of an adaptive response. J. Bacteriol. 1988, 170, 2344–2351. [Google Scholar] [CrossRef] [PubMed]
- Mueller, E.G.; Buck, C.J.; Palenchar, P.M.; Barnhart, L.E.; Paulson, J.L. Identification of a gene involved in the generation of 4-thiouridine in tRNA. Nucleic Acids Res. 1998, 26, 2060–2610. [Google Scholar] [CrossRef]
- Kambampati, R.; Lauhon, C.T. IscS is a sulfurtransferase for the in vitro biosynthesis of 4-thiouridine in Escherichia coli tRNA. Biochemistry 1999, 38, 16561–16568. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Saneyoshi, M.; Kumagai, I.; Miura, K.; Watanabe, K. Effects of modification of 4-thiouridine in E. coli tRNAMetf on its methyl acceptor activity by thermostable Gm-methylases. J. Biochem. 1989, 106, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Naumann, P.T.; Lauhon, C.T.; Ficner, R. Purification, crystallization and preliminary crystallographic analysis of a 4-thiouridine synthetase-RNA complex. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Neumann, P.; Lakomek, K.; Naumann, P.T.; Erwin, W.M.; Lauhon, C.T.; Ficner, R. Crystal structure of a 4-thiouridine synthetase-RNA complex reveals specificity of tRNA U8 modification. Nucleic Acids Res. 2014, 42, 6673–6685. [Google Scholar] [CrossRef] [PubMed]
- Nomura, Y.; Ohno, S.; Nishikawa, K.; Yokogawa, T. Correlation between the stability of tRNA tertiary structure and the catalytic efficiency of a tRNA-modifying enzyme, archaeal tRNA-guanine transglycosylase. Genes Cells 2016, 21, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Vinyard, D.J.; Reesbeck, M.E.; Suzuki, T.; Manakongtreecheep, K.; Holland, P.L.; Brudvig, G.W.; Söll, D. A [3Fe-4S] cluster is required for tRNA thiolation in archaea and eukaryotes. Proc. Natl. Acad. Sci. USA 2016, 113, 12703–12708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čavužić, M.; Liu, Y. Biosynthesis of sulfur-containing tRNA modifications: A comparison of bacterial, archaeal, and eukaryotic pathways. Biomolecules 2017, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Tomikawa, C.; Ohira, T.; Inoue, Y.; Kawamura, T.; Yamagishi, A.; Suzuki, T.; Hori, H. Distinct tRNA modifications in the thermo-acidophilic archaeon, Thermoplasma acidophilum. FEBS Lett. 2013, 587, 3537–3580. [Google Scholar] [CrossRef] [PubMed]
- Kempenaers, M.; Roovers, M.; Oudjama, Y.; Tkaczuk, K.L.; Bujnicki, J.M.; Droogmans, L. New archaeal methyltransferases forming 1-methyladenosine or 1-methyladenosine and 1-methylguanosine at position 9 of tRNA. Nucleic Acids Res. 2010, 38, 6533–6543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Laer, B.; Roovers, M.; Wauters, L.; Kasprzak, J.M.; Dyzma, M.; Deyaert, E.; Kumar Singh, R.; Feller, A.; Bujnicki, J.M.; Droogmans, L.; et al. Structural and functional insights into tRNA binding and adenosine N1-methylation by an archaeal Trm10 homologue. Nucleic Acids Res. 2016, 44, 940–953. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Feller, A.; Roovers, M.; Van Elder, D.; Wauters, L.; Droogmans, L.; Versées, W. Structural and biochemical analysis of the dual-specificity Trm10 enzyme from Thermococcus kodakaraensis prompts reconsideration of its catalytic mechanism. RNA 2018, 24, 1080–1092. [Google Scholar] [CrossRef] [PubMed]
- Armengaud, J.; Urbonavicius, J.; Fernandez, B.; Chaussinand, G.; Bujnicki, J.M.; Grosjean, H. N2-methylation of guanosine at position 10 in tRNA is catalyzed by a THUMP domain-containing, S-adenosylmethionine-dependent methyltransferase, conserved in Archaea and Eukaryota. J. Biol. Chem. 2004, 279, 37142–37152. [Google Scholar] [CrossRef] [PubMed]
- Gabant, G.; Auxilien, S.; Tuszynska, I.; Locard, M.; Gajda, M.J.; Chaussinand, G.; Fernandez, B.; Dedieu, A.; Grosjean, H.; Golinelli-Pimpaneau, B.; et al. THUMP from archaeal tRNA:m22G10 methyltransferase, a genuine autonomously folding domain. Nucleic Acids Res. 2006, 34, 2483–2494. [Google Scholar] [CrossRef] [PubMed]
- Urbonavicius, J.; Armengaud, J.; Grosjean, H. Identity elements required for enzymatic formation of N2,N2-dimethylguanosine from N2-monomethylated derivative and its possible role in avoiding alternative conformations in archaeal tRNA. J. Mol. Biol. 2006, 357, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Hirata, A.; Nishiyama, S.; Tamura, T.; Yamauchi, A.; Hori, H. Structural and functional analyses of the archaeal tRNA m2G/m22G10 methyltransferase aTrm11 provide mechanistic insights into site specificity of a tRNA methyltransferase that contains common RNA-binding modules. Nucleic Acids Res. 2016, 44, 6377–6390. [Google Scholar] [CrossRef] [PubMed]
- Kuchino, Y.; Ihara, M.; Yabusaki, Y.; Nishimura, S. Initiator tRNAs from archaebacteria show common unique sequence characteristics. Nature 1982, 298, 684–685. [Google Scholar] [CrossRef] [PubMed]
- Kaya, Y.; Ofengand, J. A novel unanticipated type of pseudouridine synthase with homologs in bacteria, archaea, and eukarya. RNA 2003, 9, 711–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, S.; Urban, A.; Hecker, A.; Leclerc, F.; Branlant, C.; Motorin, Y. Deficiency of the tRNATyr: Psi 35-synthase aPus7 in Archaea of the Sulfolobales order might be rescued by the H/ACA sRNA-guided machinery. Nucleic Acids Res. 2009, 37, 1308–1322. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Hirata, A.; Ohno, S.; Nomura, Y.; Nagano, T.; Nameki, N.; Yokogawa, T.; Hori, H. Multisite-specific archaeosine tRNA-guanine transglycosylase (ArcTGT) from Thermoplasma acidophilum, a thermo-acidophilic archaeon. Nucleic Acids Res. 2016, 44, 1894–1908. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R. Halobacterium volcanii tRNAs. Identification of 41 tRNAs covering all amino acids, and the sequences of 33 class I tRNAs. J. Biol. Chem. 1984, 259, 9461–9471. [Google Scholar] [PubMed]
- Gregson, J.M.; Crain, P.F.; Edmonds, C.G.; Gupta, R.; Hashizume, T.; Phillipson, D.W.; McCloskey, J.A. Structure of the archaeal transfer RNA nucleoside G*-15 (2-amino-4,7-dihydro-4-oxo-7-beta-D-ribofuranosyl-1H-pyrrolo[2,3-d]pyrimidine-5-carboximi dam ide (archaeosine)). J. Biol. Chem. 1993, 268, 10076–10086. [Google Scholar] [PubMed]
- Watanabe, M.; Matsuo, M.; Tanaka, S.; Akimoto, H.; Asahi, S.; Nishimura, S.; Katze, J.R.; Hashizume, T.; Crain, P.F.; McCloskey, J.A.; et al. Biosynthesis of archaeosine, a novel derivative of 7-deazaguanosine specific to archaeal tRNA, proceeds via a pathway involving base replacement on the tRNA polynucleotide chain. J. Biol. Chem. 1997, 272, 20146–20151. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Fox, D.T.; Lacy, J.A.; Van Lanen, S.G.; Iwata-Reuyl, D. Hypermodification of tRNA in Thermophilic archaea. Cloning, overexpression, and characterization of tRNA-guanine transglycosylase from Methanococcus jannaschii. J. Biol. Chem. 2000, 275, 28731–28738. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Nameki, N.; Matsuo-Takasaki, M.; Nishimura, S.; Okada, N. tRNA recognition of tRNA-guanine transglycosylase from a hyperthermophilic archaeon, Pyrococcus horikoshii. J. Biol. Chem. 2001, 276, 2387–2394. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, R.; Nureki, O.; Kijimoto, T.; Watanabe, M.; Kondo, H.; Nameki, N.; Okada, N.; Nishimura, S.; Yokoyama, S. Crystallization and preliminary X-ray analysis of the archaeosine tRNA-guanine transglycosylase from Pyrococcus horikoshii. Acta Crystallogr. D Biol. Crystallogr. 2001, 57, 1659–1662. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, R.; Nureki, O.; Fukai, S.; Kijimoto, T.; Nameki, N.; Watanabe, M.; Kondo, H.; Sekine, M.; Okada, N.; Nishimura, S.; et al. Crystal structure of archaeosine tRNA-guanine transglycosylase. J. Mol. Biol. 2002, 318, 665–677. [Google Scholar] [CrossRef]
- Ishitani, R.; Nureki, O.; Fukai, S.; Kijimoto, T.; Nameki, N.; Watanabe, M.; Kondo, H.; Sekine, M.; Okada, N.; Nishimura, S.; et al. Alternative tertiary structure of tRNA for recognition by a posttranscriptional modification enzyme. Cell 2003, 113, 383–394. [Google Scholar] [CrossRef]
- Noon, K.R.; Guymon, R.; Crain, P.F.; McCloskey, J.A.; Thomm, M.; Lim, J.; Cavicchioli, R. Influence of temperature on tRNA modification in archaea: Methanococcoides burtonii (optimum growth temperature [Topt], 23 degrees C) and Stetteria hydrogenophila (Topt, 95 degrees C). J. Bacteriol. 2003, 185, 5483–5490. [Google Scholar] [CrossRef] [PubMed]
- Sabina, J.; Söll, D. The RNA-binding PUA domain of archaeal tRNA-guanine transglycosylase is not required for archaeosine formation. J. Biol. Chem. 2006, 281, 6993–7001. [Google Scholar] [CrossRef] [PubMed]
- Oliva, R.; Tramontano, A.; Cavallo, L. Mg2+ binding and archaeosine modification stabilize the G15 C48 Levitt base pair in tRNAs. RNA 2007, 13, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Phillips, G.; Chikwana, V.M.; Maxwell, A.; El-Yacoubi, B.; Swairjo, M.A.; Iwata-Reuyl, D.; de Crécy-Lagard, V. Discovery and characterization of an amidinotransferase involved in the modification of archaeal tRNA. J. Biol. Chem. 2010, 285, 12706–12713. [Google Scholar] [CrossRef] [PubMed]
- Phillips, G.; Swairjo, M.A.; Gaston, K.W.; Bailly, M.; Limbach, P.A.; Iwata-Reuyl, D.; de Crécy-Lagard, V. Diversity of archaeosine synthesis in crenarchaeota. ACS Chem. Biol. 2012, 7, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Nomura, Y.; Onda, Y.; Ohno, S.; Taniguchi, H.; Ando, K.; Oka, N.; Nishikawa, K.; Yokogawa, T. Purification and comparison of native and recombinant tRNA-guanine transglycosylases from Methanosarcina acetivorans. Protein Expr. Purif. 2013, 88, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Alvarez, J.; Bon Ramos, A.; Samanta, U.; Iwata-Reuyl, D.; Swairjo, M.A. Crystal structure of the archaeosine synthase QueF-like-Insights into amidino transfer and tRNA recognition by the tunnel fold. Proteins 2017, 85, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Bon Ramos, A.; Bao, L.; Turner, B.; de Crécy-Lagard, V.; Iwata-Reuyl, D. QueF-Like, a Non-homologous archaeosine synthase from the crenarchaeota. Biomolecules 2017, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Takada-Guerrier, C.; Grosjean, H.; Dirheimer, G.; Keith, G. The primary structure of tRNA2Val from Bacillus stearothermophilus. FEBS Lett. 1976, 62, 1–3. [Google Scholar] [CrossRef]
- Bishop, A.C.; Xu, J.; Johnson, R.C.; Schimmel, P.; de Crécy-Lagard, V. Identification of the tRNA-dihydrouridine synthase family. J. Biol. Chem. 2002, 277, 25090–25095. [Google Scholar] [CrossRef] [PubMed]
- Bou-Nader, C.; Montémont, H.; Guérineau, V.; Jean-Jean, O.; Brégeon, D.; Hamdane, D. Unveiling structural and functional divergences of bacterial tRNA dihydrouridine synthases: Perspectives on the evolution scenario. Nucleic Acids Res. 2018, 46, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Horie, N.; Hara-Yokoyama, M.; Yokoyama, S.; Watanabe, K.; Kuchino, Y.; Nishimura, S.; Miyazawa, T. Two tRNAIle1 species from an extreme thermophile, Thermus thermophilus HB8: Effect of 2-thiolation of ribothymidine on the thermostability of tRNA. Biochemistry 1985, 24, 5711–5715. [Google Scholar] [CrossRef] [PubMed]
- Persson, B.C.; Jäger, G.; Gustafsson, C. The spoU gene of Escherichia coli, the fourth gene of the spoT operon, is essential for tRNA (Gm18) 2′-O-methyltransferase activity. Nucleic Acids Res. 1997, 25, 4093–4097. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, I.; Watanabe, K.; Oshima, T. A thermostable tRNA (guanosine-2′)-methyltransferase from Thermus. thermophilus HB27 and the effect of ribose methylation on the conformational stability of tRNA. J. Biol. Chem. 1982, 257, 7388–7395. [Google Scholar] [PubMed]
- Matsumoto, T.; Ohta, T.; Kumagai, I.; Oshima, T.; Murao, K.; Hasegawa, T.; Ishikura, H.; Watanabe, K. A thermostable Gm-methylase recognizes the tertiary structure of tRNA. J. Biochem. 1987, 101, 1191–1198. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Nishikawa, K.; Hori, H.; Ohta, T.; Miura, K.; Watanabe, K. Recognition sites of tRNA by a thermostable tRNA(guanosine-2′-)-methyltransferase from Thermus thermophilus HB27. J. Biochem. 1990, 107, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Yamazaki, N.; Matsumoto, T.; Watanabe, Y.; Ueda, T.; Nishikawa, K.; Kumagai, I.; Watanabe, K. Substrate recognition of tRNA (Guanosine-2′-)-methyltransferase from Thermus thermophilus HB27. J. Biol. Chem. 1998, 273, 25721–25727. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Suzuki, T.; Sugawara, K.; Inoue, Y.; Shibata, T.; Kuramitsu, S.; Yokoyama, S.; Oshima, T.; Watanabe, K. Identification and characterization of tRNA (Gm18) methyltransferase from Thermus thermophilus HB8: Domain structure and conserved amino acid sequence motifs. Genes Cells 2002, 7, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Kubota, S.; Watanabe, K.; Kim, J.M.; Ogasawara, T.; Sawasaki, T.; Endo, Y. Aquifex aeolicus tRNA (Gm18) methyltransferase has unique substrate specificity: tRNA recognition mechanism of the enzyme. J. Biol. Chem. 2003, 278, 25081–25090. [Google Scholar] [CrossRef] [PubMed]
- Nureki, O.; Watanabe, K.; Fukai, S.; Ishii, R.; Endo, Y.; Hori, H.; Yokoyama, S. Deep knot structure for construction of active site and cofactor binding site of tRNA modification enzyme. Structure 2004, 12, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Nureki, O.; Fukai, S.; Ishii, R.; Okamoto, H.; Yokoyama, S.; Endo, Y.; Hori, H. Roles of conserved amino acid sequence motifs in the SpoU (TrmH) RNA methyltransferase family. J. Biol. Chem. 2005, 280, 10368–10377. [Google Scholar] [CrossRef] [PubMed]
- Pleshe, E.; Truesdell, J.; Batey, R.T. Structure of a class II TrmH tRNA-modifying enzyme from Aquifex aeolicus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2005, 61, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Nureki, O.; Fukai, S.; Endo, Y.; Hori, H. Functional categorization of the conserved basic amino acid residues in TrmH (tRNA (Gm18) methyltransferase) enzymes. J. Biol. Chem. 2006, 281, 34630–34639. [Google Scholar] [CrossRef] [PubMed]
- Ochi, A.; Makabe, K.; Kuwajima, K.; Hori, H. Flexible recognition of the tRNA G18 methylation target site by TrmH methyltransferase through first binding and induced fit processes. J. Biol. Chem. 2010, 285, 9018–9029. [Google Scholar] [CrossRef] [PubMed]
- Ochi, A.; Makabe, K.; Yamagami, R.; Hirata, A.; Sakaguchi, R.; Hou, Y.M.; Watanabe, K.; Nureki, O.; Kuwajima, K.; Hori, H. The catalytic domain of topological knot tRNA methyltransferase (TrmH) discriminates between substrate tRNA and nonsubstrate tRNA via an induced-fit process. J. Biol. Chem. 2013, 288, 25562–25574. [Google Scholar] [CrossRef] [PubMed]
- Hori, H.; Terui, Y.; Nakamoto, C.; Iwashita, C.; Ochi, A.; Watanabe, K.; Oshima, T. Effects of polyamines from Thermus thermophilus, an extreme-thermophilic eubacterium, on tRNA methylation by tRNA (Gm18) methyltransferase (TrmH). J. Biochem. 2016, 159, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Savage, D.F.; de Crécy-Lagard, V.; Bishop, A.C. Molecular determinants of dihydrouridine synthase activity. FEBS Lett. 2006, 580, 5198–5202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Tanaka, Y.; Yamamoto, S.; Nakamura, A.; Kita, S.; Hirano, N.; Tanaka, I.; Yao, M. Crystallization and preliminary X-ray crystallographic analysis of dihydrouridine synthase from Thermus thermophilus and its complex with tRNA. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Tanaka, Y.; Yamashita, K.; Suzuki, T.; Nakamura, A.; Hirano, N.; Suzuki, T.; Yao, M.; Tanaka, I. Molecular basis of dihydrouridine formation on tRNA. Proc. Natl. Acad. Sci. USA 2011, 108, 19593–19598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusuba, H.; Yoshida, T.; Iwasaki, E.; Awai, T.; Kazayama, A.; Hirata, A.; Tomikawa, C.; Yamagami, R.; Hori, H. In vitro dihydrouridine formation by tRNA dihydrouridine synthase from Thermus thermophilus, an extreme-thermophilic eubacterium. J. Biochem. 2015, 158, 513–521. [Google Scholar] [PubMed]
- Roovers, M.; Kaminska, K.H.; Tkaczuk, K.L.; Gigot, D.; Droogmans, L.; Bujnicki, J.M. The YqfN protein of Bacillus subtilis is the tRNA: M1A22 methyltransferase (TrmK). Nucleic Acids Res. 2008, 36, 3252–3262. [Google Scholar] [CrossRef] [PubMed]
- Blaby, I.K.; Majumder, M.; Chatterjee, K.; Jana, S.; Grosjean, H.; de Crécy-Lagard, V.; Gupta, R. Pseudouridine formation in archaeal RNAs: The case of Haloferax volcanii. RNA 2011, 17, 1367–1380. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, C.; Lünse, C.E.; Mörl, M. tRNA modifications: Impact on structure and thermal adaptation. Biomolecules 2017, 7, 35. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Anraku, R.; Hasegawa, T.; Tomikawa, C.; Hori, H. Transfer RNA methyltransferases from Thermoplasma acidophilum, a thermoacidophilic archaeon. Int. J. Mol. Sci. 2014, 16, 91–113. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.H.; Kjellin-Straby, K. Studies on microbial ribonucleic acid. IV. Two mutants of Saccharomyces cerevisiae lacking N-2-dimethylguanine in soluble ribonucleic acid. J. Mol. Biol. 1967, 26, 509–518. [Google Scholar] [CrossRef]
- Constantinesco, F.; Benachenhou, N.; Motorin, Y.; Grosjean, H. The tRNA(guanine-26,N2-N2) methyltransferase (Trm1) from the hyperthermophilic archaeon Pyrococcus furiosus: Cloning, sequencing of the gene and its expression in Escherichia coli. Nucleic Acids Res. 1998, 26, 3753–3761. [Google Scholar] [CrossRef] [PubMed]
- Constantinesco, F.; Motorin, Y.; Grosjean, H. Characterisation and enzymatic properties of tRNA (guanine 26, N (2), N (2))-dimethyltransferase (Trm1p) from Pyrococcus furiosus. J. Mol. Biol. 1999, 291, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Ihsanawati; Nishimoto, M.; Higashijima, K.; Shirouzu, M.; Grosjean, H.; Bessho, Y.; Yokoyama, S. Crystal structure of tRNA N2,N2-guanosine dimethyltransferase Trm1 from Pyrococcus horikoshii. J. Mol. Biol. 2008, 383, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Vakiloroayaei, A.; Shah, N.S.; Oeffinger, M.; Bayfield, M.A. The RNA chaperone La promotes pre-tRNA maturation via indiscriminate binding of both native and misfolded targets. Nucleic Acids Res. 2017, 45, 11341–11355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awai, T.; Ochi, A.; Ihsanawati; Sengoku, T.; Hirata, A.; Bessho, Y.; Yokoyama, S.; Hori, H. Substrate tRNA recognition mechanism of a multisite-specific tRNA methyltransferase, Aquifex aeolicus Trm1, based on the X-ray crystal structure. J. Biol. Chem. 2011, 286, 35236–35246. [Google Scholar] [CrossRef] [PubMed]
- Somme, J.; Van Laer, B.; Roovers, M.; Steyaert, J.; Versées, W.; Droogmans, L. Characterization of two homologous 2′-O-methyltransferases showing different specificities for their tRNA substrates. RNA 2014, 20, 1257–1271. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, C.G.; Crain, P.F.; Gupta, R.; Hashizume, T.; Hocart, C.H.; Kowalak, J.A.; Pomerantz, S.C.; Stetter, K.O.; McCloskey, J.A. Posttranscriptional modification of tRNA in Thermophilic archaea (Archaebacteria). J. Bacteriol. 1991, 173, 3138–3148. [Google Scholar] [CrossRef] [PubMed]
- Kowalak, J.A.; Dalluge, J.J.; McCloskey, J.A.; Stetter, K.O. The role of posttranscriptional modification in stabilization of transfer RNA from Hyperthermophiles. Biochemistry 1994, 33, 7869–7876. [Google Scholar] [CrossRef] [PubMed]
- McCloskey, J.A.; Graham, D.E.; Zou, S.; Crain, P.F.; Ibba, M.; Konisky, J.; Soll, D.; Olsen, G.J. Post-transcriptional modification in archaeal tRNAs: Identities and phylogenetic relations of nucleotides from mesophilic and hyperthermophilic Methanococcales. Nucleic Acids Res. 2001, 29, 4299–4706. [Google Scholar] [CrossRef]
- Sauerwald, A.; Sitaramaiah, D.; McCloskey, J.A.; Söll, D.; Crain, P.F. N6-Acetyladenosine: A new modified nucleoside from Methanopyrus kandleri tRNA. FEBS Lett. 2005, 579, 2807–2810. [Google Scholar] [CrossRef] [PubMed]
- Purta, E.; van Vliet, F.; Tkaczuk, K.L.; Dunin-Horkawicz, S.; Mori, H.; Droogmans, L.; Bujnicki, J.M. The yfhQ gene of Escherichia coli encodes a tRNA: Cm32/Um32 methyltransferase. BMC Mol. Biol. 2006, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Jaroensuk, J.; Atichartpongkul, S.; Chionh, Y.H.; Wong, Y.H.; Liew, C.W.; McBee, M.E.; Thongdee, N.; Prestwich, E.G.; DeMott, M.S.; Mongkolsuk, S.; et al. Methylation at position 32 of tRNA catalyzed by TrmJ alters oxidative stress response in Pseudomonas aeruginosa. Nucleic Acids Res. 2016, 44, 10834–10848. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.; Gerber, A.P.; Keller, W. tadA, an essential tRNA-specific adenosine deaminase from Escherichia coli. EMBO J. 2002, 21, 3841–3851. [Google Scholar] [CrossRef] [PubMed]
- Elias, Y.; Huang, R.H. Biochemical and structural studies of A-to-I editing by tRNA: A34 deaminases at the wobble position of transfer RNA. Biochemistry 2005, 44, 12057–12065. [Google Scholar] [CrossRef] [PubMed]
- Kuratani, M.; Ishii, R.; Bessho, Y.; Fukunaga, R.; Sengoku, T.; Shirouzu, M.; Sekine, S.; Yokoyama, S. Crystal structure of tRNA adenosine deaminase (TadA) from Aquifex aeolicus. J. Biol. Chem. 2005, 280, 16002–16008. [Google Scholar] [CrossRef] [PubMed]
- Yokobori, S.; Kitamura, A.; Grosjean, H.; Bessho, Y. Life without tRNAArg-adenosine deaminase TadA: Evolutionary consequences of decoding the four CGN codons as arginine in Mycoplasmas and other Mollicutes. Nucleic Acids Res. 2013, 41, 6531–6543. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Piñeyro, D.; Filonava, L.; Stracker, T.H.; Batlle, E.; Ribas de Pouplana, L. A-to-I editing on tRNAs: Biochemical, biological and evolutionary implications. FEBS Lett. 2014, 588, 4279–4286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muramatsu, T.; Yokoyama, S.; Horie, N.; Matsuda, A.; Ueda, T.; Yamaizumi, Z.; Kuchino, Y.; Nishimura, S.; Miyazawa, T. A novel lysine-substituted nucleoside in the first position of the anticodon of minor isoleucine tRNA from Escherichia coli. J. Biol. Chem. 1988, 263, 9261–9267. [Google Scholar] [PubMed]
- Muramatsu, T.; Nishikawa, K.; Nemoto, F.; Kuchino, Y.; Nishimura, S.; Miyazawa, T.; Muramatsu, T.; Nishikawa, K.; Nemoto, F.; Kuchino, Y.; et al. Codon and amino-acid specificities of a transfer RNA are both converted by a single post-transcriptional modification. Nature 1988, 336, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Soma, A.; Ikeuchi, Y.; Kanemasa, S.; Kobayashi, K.; Ogasawara, N.; Ote, T.; Kato, J.; Watanabe, K.; Sekine, Y.; Suzuki, T. An RNA-modifying enzyme that governs both the codon and amino acid specificities of isoleucine tRNA. Mol. Cell 2003, 12, 689–698. [Google Scholar] [CrossRef]
- Nakanishi, K.; Fukai, S.; Ikeuchi, Y.; Soma, A.; Sekine, Y.; Suzuki, T.; Nureki, O. Structural basis for lysidine formation by ATP pyrophosphatase accompanied by a lysine-specific loop and a tRNA-recognition domain. Proc. Natl. Acad. Sci. USA 2005, 102, 7487–7492. [Google Scholar] [CrossRef] [PubMed]
- Kuratani, M.; Yoshikawa, Y.; Bessho, Y.; Higashijima, K.; Ishii, T.; Shibata, R.; Takahashi, S.; Yutani, K.; Yokoyama, S. Structural basis of the initial binding of tRNA(Ile) lysidine synthetase TilS with ATP and L-lysine. Structure 2007, 15, 1642–1653. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K.; Bonnefond, L.; Kimura, S.; Suzuki, T.; Ishitani, R.; Nureki, O. Structural basis for translational fidelity ensured by transfer RNA lysidine synthetase. Nature 2009, 461, 1144–1148. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Numata, T. Convergent evolution of AUA decoding in bacteria and archaea. RNA Biol. 2014, 11, 1586–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numata, T. Mechanisms of the tRNA wobble cytidine modification essential for AUA codon decoding in prokaryotes. Biosci. Biotechnol. Biochem. 2015, 79, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, Y.; Kimura, S.; Numata, T.; Nakamura, D.; Yokogawa, T.; Ogata, T.; Wada, T.; Suzuki, T.; Suzuki, T. Agmatine-conjugated cytidine in a tRNA anticodon is essential for AUA decoding in archaea. Nat. Chem. Biol. 2010, 6, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Mandal, D.; Köhrer, C.; Su, D.; Russell, S.P.; Krivos, K.; Castleberry, C.M.; Blum, P.; Limbach, P.A.; Söll, D.; RajBhandary, U.L. Agmatidine, a modified cytidine in the anticodon of archaeal tRNA(Ile), base pairs with adenosine but not with guanosine. Proc. Natl. Acad. Sci. USA 2010, 107, 2872–2877. [Google Scholar] [CrossRef] [PubMed]
- Terasaka, N.; Kimura, S.; Osawa, T.; Numata, T.; Suzuki, T. Biogenesis of 2-agmatinylcytidine catalyzed by the dual protein and RNA kinase TiaS. Nat. Struct. Mol. Biol. 2011, 18, 1268–1274. [Google Scholar] [CrossRef] [PubMed]
- Osawa, T.; Kimura, S.; Terasaka, N.; Inanaga, H.; Suzuki, T.; Numata, T. Structural basis of tRNA agmatinylation essential for AUA codon decoding. Nat. Struct. Mol. Biol. 2011, 18, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Voorhees, R.M.; Mandal, D.; Neubauer, C.; Köhrer, C.; RajBhandary, U.L.; Ramakrishnan, V. The structural basis for specific decoding of AUA by isoleucine tRNA on the ribosome. Nat. Struct. Mol. Biol. 2013, 20, 641–643. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Hwang, J.; Inouye, M. Characterization of GTPase activity of TrmE, a member of a novel GTPase superfamily, from Thermotoga maritima. J. Bacteriol. 2000, 182, 7078–7082. [Google Scholar] [CrossRef] [PubMed]
- Urbonavicius, J.; Qian, Q.; Durand, J.M.; Hagervall, T.G.; Björk, G.R. Improvement of reading frame maintenance is a common function for several tRNA modifications. EMBO J. 2001, 20, 4863–4873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbonavicius, J.; Stahl, G.; Durand, J.M.; Ben Salem, S.N.; Qian, Q.; Farabaugh, P.J.; Björk, G.R. Transfer RNA modifications that alter +1 frameshifting in general fail to affect -1 frameshifting. RNA 2003, 9, 760–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takai, K.; Yokoyama, S. Roles of 5-substituents of tRNA wobble uridines in the recognition of purine-ending codons. Nucleic Acids Res. 2003, 31, 6383–6391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bujnicki, J.M.; Oudjama, Y.; Roovers, M.; Owczarek, S.; Caillet, J.; Droogmans, L. Identification of a bifunctional enzyme MnmC involved in the biosynthesis of a hypermodified uridine in the wobble position of tRNA. RNA 2004, 18, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Armengod, M.E.; Moukadiri, I.; Prado, S.; Ruiz-Partida, R.; Benítez-Páez, A.; Villarroya, M.; Lomas, R.; Garzón, M.J.; Martínez-Zamora, A.; Meseguer, S.; Navarro-González, C. Enzymology of tRNA modification in the bacterial MnmEG pathway. Biochimie 2012, 94, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Moukadiri, I.; Garzón, M.J.; Björk, G.R.; Armengod, M.E. The output of the tRNA modification pathways controlled by the Escherichia coli MnmEG and MnmC enzymes depends on the growth conditions and the tRNA species. Nucleic Acids Res. 2014, 42, 2602–2623. [Google Scholar] [CrossRef] [PubMed]
- Armengod, M.E.; Meseguer, S.; Villarroya, M.; Prado, S.; Moukadiri, I.; Ruiz-Partida, R.; Garzón, M.J.; Navarro-González, C.; Martínez-Zamora, A. Modification of the wobble uridine in bacterial and mitochondrial tRNAs reading NNA/NNG triplets of 2-codon boxes. RNA Biol. 2014, 11, 1495–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalhor, H.R.; Clarke, S. Novel methyltransferase for modified uridine residues at the wobble position of tRNA. Mol. Cell Biol. 2003, 23, 9283–9292. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Johansson, M.J.; Byström, A.S. An early step in wobble uridine tRNA modification requires the Elongator complex. RNA 2005, 11, 424–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begley, U.; Dyavaiah, M.; Patil, A.; Rooney, J.P.; DiRenzo, D.; Young, C.M.; Conklin, D.S.; Zitomer, R.S.; Begley, T.J. Trm9-catalyzed tRNA modifications link translation to the DNA damage response. Mol. Cell 2007, 28, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Mazauric, M.H.; Dirick, L.; Purushothaman, S.K.; Björk, G.R.; Lapeyre, B. Trm112p is a 15-kDa zinc finger protein essential for the activity of two tRNA and one protein methyltransferases in yeast. J. Biol. Chem. 2010, 285, 18505–18515. [Google Scholar] [CrossRef] [PubMed]
- Chavarria, N.E.; Hwang, S.; Cao, S.; Fu, X.; Holman, M.; Elbanna, D.; Rodriguez, S.; Arrington, D.; Englert, M.; Uthandi, S.; et al. Archaeal Tuc1/Ncs6 homolog required for wobble uridine tRNA thiolation is associated with ubiquitin-proteasome, translation, and RNA processing system homologs. PLoS ONE 2014, 9, e99104. [Google Scholar] [CrossRef] [PubMed]
- Arragain, S.; Bimai, O.; Legrand, P.; Caillat, S.; Ravanat, J.L.; Touati, N.; Binet, L.; Atta, M.; Fontecave, M.; Golinelli-Pimpaneau, B. Nonredox thiolation in tRNA occurring via sulfur activation by a [4Fe-4S] cluster. Proc. Natl. Acad. Sci. USA 2017, 114, 7355–7360. [Google Scholar] [CrossRef] [PubMed]
- Black, K.A.; Dos Satos, P.C. Abbreviated Pathway for Biosynthesis of 2-Thiouridine in Bacillus subtilis. J. Bacteriol. 2015, 197, 1952–1962. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, A.; Nishimoto, M.; Sengoku, T.; Shibata, R.; Jäger, G.; Björk, G.R.; Grosjean, H.; Yokoyama, S.; Bessho, Y. Characterization and structure of the Aquifex aeolicus protein DUF752: A bacterial tRNA-methyltransferase (MnmC2) functioning without the usually fused oxidase domain (MnmC1). J. Biol. Chem. 2012, 287, 43950–43960. [Google Scholar] [CrossRef] [PubMed]
- Mandal, D.; Köhrer, C.; Su, D.; Babu, I.R.; Chan, C.T.; Liu, Y.; Söll, D.; Blum, P.; Kuwahara, M.; Dedon, P.C.; Rajbhandary, U.L. Identification and codon reading properties of 5-cyanomethyl uridine, a new modified nucleoside found in the anticodon wobble position of mutant haloarchaeal isoleucine tRNAs. RNA 2014, 20, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Scrima, A.; Vetter, I.R.; Armengod, M.E.; Wittinghofer, A. The structure of the TrmE GTP-binding protein and its implications for tRNA modification. EMBO J. 2005, 24, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Scrima, A.; Wittinghofer, A. Dimerisation-dependent GTPase reaction of MnmE: How potassium acts as GTPase-activating element. EMBO J. 2006, 25, 2940–2951. [Google Scholar] [CrossRef] [PubMed]
- Osawa, T.; Inanaga, H.; Numata, T. Crystallization and preliminary X-ray diffraction analysis of the tRNA-modification enzyme GidA from Aquifex aeolicus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2009, 65, 508–511. [Google Scholar] [CrossRef] [PubMed]
- Osawa, T.; Ito, K.; Inanaga, H.; Nureki, O.; Tomita, K.; Numata, T. Conserved cysteine residues of GidA are essential for biogenesis of 5-carboxymethylaminomethyluridine at tRNA anticodon. Structure 2009, 17, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Selcadurai, K.; Wang, P.; Seimetz, J.; Huang, R.H. Archaeal Elp3 catalyzes tRNA wobble uridine modification at C5 via a radical mechanism. Nat. Chem. Biol. 2014, 10, 810–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benítez-Páez, A.; Villarroya, M.; Douthwaite, S.; Gabaldón, T.; Armengod, M.E. YibK is the 2′-O-methyltransferase TrmL that modifies the wobble nucleotide in Escherichia coli tRNA(Leu) isoacceptors. RNA 2010, 16, 2131–2143. [Google Scholar] [CrossRef] [PubMed]
- Pang, P.; Deng, X.; Wang, Z.; Xie, W. Structural and biochemical insights into the 2′-O-methylation of pyrimidines 34 in tRNA. FEBS J. 2017, 284, 2251–2263. [Google Scholar] [CrossRef] [PubMed]
- Okada, N.; Noguchi, S.; Nishimura, S.; Ohgi, T.; Goto, T.; Crain, P.F.; McCloskey, J.A. Structure determination of a nucleoside Q precursor isolated from E. coli tRNA: 7-(aminomethyl)-7-deazaguanosine. Nucleic Acids Res. 1978, 5, 2289–2296. [Google Scholar] [CrossRef] [PubMed]
- Okada, N.; Noguchi, S.; Kasai, H.; Shindo-Okada, N.; Ohgi, T.; Goto, T.; Nishimura, S. Novel mechanism of post-transcriptional modification of tRNA. Insertion of bases of Q precursors into tRNA by a specific tRNA transglycosylase reaction. J. Biol. Chem. 1979, 254, 3067–3073. [Google Scholar] [PubMed]
- Nakanishi, S.; Ueda, T.; Hori, H.; Yamazaki, N.; Okada, N.; Watanabe, K. A UGU sequence in the anticodon loop is a minimum requirement for recognition by Escherichia coli tRNA-guanine transglycosylase. J. Biol. Chem. 1994, 269, 32221–32225. [Google Scholar] [PubMed]
- Slany, R.K.; Bösl, M.; Kersten, H. Transfer and isomerization of the ribose moiety of AdoMet during the biosynthesis of queuosine tRNAs, a new unique reaction catalyzed by the QueA protein from Escherichia coli. Biochimie 1994, 76, 389–393. [Google Scholar] [CrossRef]
- Van Lanen, S.G.; Kinzie, S.D.; Matthieu, S.; Link, T.; Culp, J.; Iwata-Reuyl, D. tRNA modification by S-adenosylmethionine: tRNA ribosyltransferase-isomerase. Assay development and characterization of the recombinant enzyme. J. Biol. Chem. 2003, 278, 10491–10499. [Google Scholar] [CrossRef] [PubMed]
- Miles, Z.D.; Myers, W.K.; Kincannon, W.M.; Britt, R.D.; Bandarian, V. Biochemical and Spectroscopic Studies of Epoxyqueuosine Reductase: A Novel Iron-Sulfur Cluster- and Cobalamin-Containing Protein Involved in the Biosynthesis of Queuosine. Biochemistry 2015, 54, 4927–4935. [Google Scholar] [CrossRef] [PubMed]
- Mathews, I.; Schwarzenbacher, R.; McMullan, D.; Abdubek, P.; Ambing, E.; Axelrod, H.; Biorac, T.; Canaves, J.M.; Chiu, H.J.; Deacon, A.M.; et al. Crystal structure of S-adenosylmethionine: tRNA ribosyltransferase-isomerase (QueA) from Thermotoga maritima at 2.0 A resolution reveals a new fold. Proteins 2005, 59, 869–974. [Google Scholar] [CrossRef] [PubMed]
- Vinayak, M.; Pathak, C. Queuosine modification of tRNA: Its divergent role in cellular machinery. Biosci. Rep. 2009, 30, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Hutinet, G.; Swarjo, M.A.; de Crécy-Lagard, V. Deazaguanine derivatives, examples of crosstalk between RNA and DNA modification pathways. RNA Biol. 2017, 14, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Clouet d’Orval, B.; Bortolin, M.L.; Gaspin, C.; Bachellerie, J.P. Box C/D RNA guides for the ribose methylation of archaeal tRNAs. The tRNATrp intron guides the formation of two ribose-methylated nucleosides in the mature tRNATrp. Nucleic Acids Res. 2001, 29, 4518–4529. [Google Scholar] [CrossRef] [PubMed]
- Bortolin, M.L.; Bachellerie, J.P.; Clouet-d’Orval, B. In vitro RNP assembly and methylation guide activity of an unusual box C/D RNA, cis-acting archaeal pre-tRNATrp. Nucleic Acids Res. 2003, 31, 6524–6535. [Google Scholar] [CrossRef] [PubMed]
- Byström, A.S.; Björk, G.R. Chromosomal location and cloning of the gene (trmD) responsible for the synthesis of tRNA (m1G) methyltransferase in Escherichia coli K-12. Mol. Gen. Genet. 1982, 188, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Björk, G.R.; Wikstrom, P.M.; Byström, A.S. Prevention of translational frameshifting by the modified nucleoside 1-methylguanosine. Science 1989, 244, 986–989. [Google Scholar] [CrossRef] [PubMed]
- Farabaugh, P.J.; Björk, G.R. How translational accuracy influences reading frame maintenance. EMBO J. 1999, 18, 1427–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björk, G.R.; Jacobsson, K.; Nilsson, K.; Johansson, M.J.; Byström, A.S.; Persson, O.P. A primordial tRNA modification required for the evolution of life? EMBO J. 2001, 20, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Wang, W.; Shin, D.H.; Yokota, H.; Kim, R.; Kim, S.H. Crystal structure of tRNA (m1G37) methyltransferase from Aquifex aeolicus at 2.6 A resolution: A novel methyltransferase fold. Proteins 2003, 53, 326–328. [Google Scholar] [CrossRef] [PubMed]
- Christian, T.; Evilia, C.; Williams, S.; Hou, Y.M. Distinct origins of tRNA(m1G37) methyltransferase. J. Mol. Biol. 2004, 339, 707–719. [Google Scholar] [CrossRef] [PubMed]
- Takeda, H.; Toyooka, T.; Ikeuchi, Y.; Yokobori, S.; Okadome, K.; Takano, F.; Oshima, T.; Suzuki, T.; Endo, Y.; Hori, H. The substrate specificity of tRNA (m1G37) methyltransferase (TrmD) from Aquifex aeolicus. Genes Cells 2006, 11, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Christian, T.; Evilia, C.; Hou, Y.M. Catalysis by the second class of tRNA(m1G37) methyl transferase requires a conserved proline. Biochemistry 2006, 45, 7463–7473. [Google Scholar] [CrossRef] [PubMed]
- Christian, T.; Hou, Y.M. Distinct determinants of tRNA recognition by the TrmD and Trm5 methyl transferases. J. Mol. Biol. 2007, 373, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Toyooka, T.; Awai, T.; Kanai, T.; Imanaka, T.; Hori, H. Stabilization of tRNA (m1G37) methyltransferase [TrmD] from Aquifex aeolicus by an intersubunit disulfide bond formation. Genes Cells 2008, 13, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Goto-Ito, S.; Ito, T.; Ishii, R.; Muto, Y.; Bessho, Y.; Yokoyama, S. Crystal structure of archaeal tRNA(m(1)G37)methyltransferase aTrm5. Proteins 2008, 72, 1274–1289. [Google Scholar] [CrossRef] [PubMed]
- Goto-Ito, S.; Ito, T.; Kuratani, M.; Bessho, Y.; Yokoyama, S. Tertiary structure checkpoint at anticodon loop modification in tRNA functional maturation. Nat. Struct. Mol. Biol. 2009, 16, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Lahoud, G.; Goto-Ito, S.; Yoshida, K.; Ito, T.; Yokoyama, S.; Hou, Y.M. Differentiating analogous tRNA methyltransferases by fragments of the methyl donor. RNA 2011, 17, 1236–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, R.; Giessing, A.; Dai, Q.; Lahoud, G.; Liutkeviciute, Z.; Klimasauskas, S.; Piccirilli, J.; Kirpekar, F.; Hou, Y.M. Recognition of guanosine by dissimilar tRNA methyltransferases. RNA 2012, 18, 1687–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christian, T.; Gamper, H.; Hou, Y.M. Conservation of structure and mechanism by Trm5 enzymes. RNA 2013, 19, 1192–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Masuda, I.; Yoshida, K.; Goto-Ito, S.; Sekine, S.; Suh, S.W.; Hou, Y.M.; Yokoyama, S. Structural basis for methyl-donor-dependent and sequence-specific binding to tRNA substrates by knotted methyltransferase TrmD. Proc. Natl. Acad. Sci. USA 2015, 112, E4197–4205. [Google Scholar] [CrossRef] [PubMed]
- Goto-Ito, S.; Ishii, R.; Ito, T.; Shibata, R.; Fusatomi, E.; Sekine, S.I.; Bessho, Y.; Yokoyama, S. Structure of an archaeal TYW1, the enzyme catalyzing the second step of wye-base biosynthesis. Acta Crystallogr. D Biol. Crystallogr. 2007, 63, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Noma, A.; Suzuki, T.; Senda, M.; Senda, T.; Ishitani, R.; Nureki, O. Crystal structure of the radical SAM enzyme catalyzing tricyclic modified base formation in tRNA. J. Mol. Biol. 2007, 372, 1204–1214. [Google Scholar] [CrossRef] [PubMed]
- Umitsu, M.; Nishimasu, H.; Noma, A.; Suzuki, T.; Ishitani, R.; Nureki, O. Structural basis of AdoMet-dependent aminocarboxypropyl transfer reaction catalyzed by tRNA-wybutosine synthesizing enzyme, TYW2. Proc. Natl. Acad. Sci. USA 2009, 106, 15616–15621. [Google Scholar] [CrossRef] [PubMed]
- Perche-Letuvée, P.; Kathirvelu, V.; Berggren, G.; Clemancey, M.; Latour, J.M.; Maurel, V.; Douki, T.; Armengaud, J.; Mulliez, E.; Fontecave, M.; et al. 4-Demethylwyosine synthase from Pyrococcus abyssi is a radical-S-adenosyl-L-methionine enzyme with an additional [4Fe-4S](+2) cluster that interacts with the pyruvate co-substrate. J. Biol. Chem. 2012, 287, 41174–41185. [Google Scholar] [CrossRef] [PubMed]
- Urbonavičius, J.; Rutkienė, R.; Lopato, A.; Tauraitė, D.; Stankevičiūtė, J.; Aučynaitėm, A.; Kaliniene, L.; van Tilbeurgh, H.; Meškys, R. Evolution of tRNAPhe: imG2 methyltransferases involved in the biosynthesis of wyosine derivatives in Archaea. RNA 2016, 22, 1871–1883. [Google Scholar]
- Wang, C.; Jia, Q.; Chen, R.; Wei, Y.; Li, J.; Ma, J.; Xie, W. Crystal structures of the bifunctional tRNA methyltransferase Trm5a. Sci. Rep. 2016, 6, 33553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currie, M.A.; Brown, G.; Wong, A.; Ohira, T.; Sugiyama, K.; Suzuki, T.; Yakunin, A.F.; Jia, Z. Structural and functional characterization of the TYW3/Taw3 class of SAM-dependent methyltransferases. RNA 2017, 23, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Jia, Q.; Zeng, J.; Chen, R.; Xie, W. Structural insight into the methyltransfer mechanism of the bifunctional Trm5. Sci. Adv. 2017, 3, e1700195. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Jia, Q.; Wu, S.; Zeng, H.; Sun, Y.; Wang, C.; Ge, R.; Xie, W. The crystal structure of the Pyrococcus abyssi mono-functional methyltransferase PaTrm5b. Biochem. Biophys. Res. Commun. 2017, 493, 240–245. [Google Scholar] [CrossRef] [PubMed]
- de Crécy-Lagard, V.; Brochier-Armanet, C.; Urbonavicius, J.; Fernandez, B.; Phillips, G.; Lyons, B.; Noma, A.; Alvarez, S.; Droogmans, L.; Armengaud, J.; et al. Biosynthesis of wyosine derivatives in tRNA: An ancient and highly diverse pathway in Archaea. Mol. Biol. Evol. 2010, 27, 2062–2077. [Google Scholar] [CrossRef] [PubMed]
- Urbonavičius, J.; Meškys, R.; Grosjean, H. Biosynthesis of wyosine derivatives in tRNA(Phe) of Archaea: Role of a remarkable bifunctional tRNA(Phe):m1G/imG2 methyltransferase. RNA 2014, 20, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Perche-Letuvée, P.; Molle, T.; Forouhar, F.; Mulliez, E.; Atta, M. Wybutosine biosynthesis: Structural and mechanistic overview. RNA Biol. 2014, 11, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.Y.; Neculai, D.; Downey, M.; Orlicky, S.; Haffani, Y.Z.; Ceccarelli, D.F.; Ho, J.S.; Szilard, R.K.; Zhang, W.; Ho, C.S.; et al. Atomic structure of the KEOPS complex: An ancient protein kinase-containing molecular machine. Mol. Cell 2008, 32, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Hecker, A.; Lopreiato, R.; Graille, M.; Collinet, B.; Forterre, P.; Libri, D.; van Tilbeurgh, H. Structure of the archaeal Kae1/Bud32 fusion protein MJ1130: A model for the eukaryotic EKC/KEOPS subcomplex. EMBO J. 2008, 27, 2340–2351. [Google Scholar] [CrossRef] [PubMed]
- Hecker, A.; Graille, M.; Madec, E.; Gadelle, D.; Le Cam, E.; van Tilbergh, H.; Forterre, P. The universal Kae1 protein and the associated Bud32 kinase (PRPK), a mysterious protein couple probably essential for genome maintenance in Archaea and Eukarya. Biochem. Soc. Trans. 2009, 37, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Perrochia, L.; Crozat, E.; Hecker, A.; Zhang, W.; Bareille, J.; Collinet, B.; van Tilbeurgh, H.; Forterre, P.; Basta, T. In vitro biosynthesis of a universal t6A tRNA modification in Archaea and Eukarya. Nucleic Acids Res. 2013, 41, 1953–1964. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.C.; Pillon, M.C.; Thevakumaran, N.; Sun, Y.; Chakrabartty, A.; Guarné, A.; Kurinov, I.; Durocher, D.; Sicheri, F. Structural and functional characterization of KEOPS dimerization by Pcc1 and its role in t6A biosynthesis. Nucleic Acids Res. 2016, 44, 6971–6980. [Google Scholar] [CrossRef] [PubMed]
- Pichard-Kostuch, A.; Zhang, W.; Liger, D.; Daugeron, M.C.; Letoquart, J.; Li de la Sierra-Gallay, I.; Forterre, P.; Collinet, B.; van Tilbeurgh, H.; Basta, T. Structure-function analysis of Sua5 protein reveals novel functional motifs required for the biosynthesis of the universal t6A tRNA modification. RNA 2018, 24, 926–938. [Google Scholar] [CrossRef] [PubMed]
- Luthra, A.; Swinehart, W.; Bayooz, S.; Phan, P.; Stec, B.; Iwata-Reuyl, D.; Swairjo, M.A. Structure and mechanism of a bacterial t6A biosynthesis system. Nucleic Acids Res. 2018, 46, 1395–1411. [Google Scholar] [CrossRef] [PubMed]
- Thiaville, P.C.; Iwata-Reuyl, D.; de Crécy-Lagard, V. Diversity of the biosynthesis pathway for threonylcarbamoyladenosine (t6A), a universal modification of tRNA. RNA Biol. 2014, 11, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Caillet, J.; Droogmans, L. Molecular cloning of the Escherichia coli miaA gene involved in the formation of delta 2-isopentenyl adenosine in tRNA. J. Bacteriol. 1988, 170, 4147–4152. [Google Scholar] [CrossRef] [PubMed]
- Esberg, B.; Leung, H.C.; Tsui, H.C.; Björk, G.R.; Winkler, M.E. Identification of the miaB gene, involved in methylthiolation of isopentenylated A37 derivatives in the tRNA of Salmonella typhimurium and Escherichia coli. J. Bacteriol. 1999, 181, 7256–7265. [Google Scholar] [PubMed]
- Pierrel, F.; Hernandez, H.L.; Johnson, M.K.; Fontecave, M.; Atta, M. MiaB protein from Thermotoga maritima. Characterization of an extremely thermophilic tRNA-methylthiotransferase. J. Biol. Chem. 2003, 278, 29515–29524. [Google Scholar] [CrossRef] [PubMed]
- Pierrel, F.; Douki, T.; Fontecave, M.; Atta, M. MiaB protein is a bifunctional radical-S-adenosylmethionine enzyme involved in thiolation and methylation of tRNA. J. Biol. Chem. 2004, 279, 47555–47563. [Google Scholar] [CrossRef] [PubMed]
- Hernández, H.L.; Pierrel, F.; Elleingand, E.; García-Serres, R.; Huynh, B.H.; Johnson, M.K.; Fontecave, M.; Atta, M. MiaB, a bifunctional radical-S-adenosylmethionine enzyme involved in the thiolation and methylation of tRNA, contains two essential [4Fe-4S] clusters. Biochemistry 2007, 46, 5140–5147. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, U.; Bohleber, S.; Fradejas-Villar, N. The modified base isopentenyladenosine and its derivatives in tRNA. RNA Biol. 2017, 14, 1197–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golovina, A.Y.; Sergiev, P.V.; Golovin, A.V.; Serebryakova, M.V.; Demina, I.; Govorun, V.M.; Dontsova, O.A. The yfiC gene of E. coli encodes an adenine-N6 methyltransferase that specifically modifies A37 of tRNA1Val(cmo5UAC). RNA 2009, 15, 1134–1141. [Google Scholar] [CrossRef] [PubMed]
- Allaudeen, H.S.; Yang, S.K.; Söll, D. Leucine tRNA(1) from HisT mutant of Salmonella typhimurium lacks two pseudouridines. FEBS Lett. 1972, 28, 205–208. [Google Scholar] [CrossRef]
- Kammen, H.O.; Marvel, C.C.; Hardy, L.; Penhoet, E.E. Purification, structure, and properties of Escherichia coli tRNA pseudouridine synthase I. J. Biol. Chem. 1988, 263, 2255–2263. [Google Scholar] [PubMed]
- Lecointe, F.; Simos, G.; Sauer, A.; Hurt, E.C.; Motorin, Y.; Grosjean, H. Characterization of yeast protein Deg1 as pseudouridine synthase (Pus3) catalyzing the formation of psi 38 and psi 39 in tRNA anticodon loop. J. Biol. Chem. 1998, 273, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Bessho, Y.; Shibata, R.; Nishimoto, M.; Shirouzu, M.; Kuramitsu, S.; Yokoyama, S. Crystal structure of tRNA pseudouridine synthase TruA from Thermus thermophilus HB8. RNA Biol. 2006, 3, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Spenkuch, F.; Motorin, Y.; Helm, M. Pseudouridine: Still mysterious, but never a fake (uridine)! RNA Biol. 2014, 11, 1540–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morozov, I.A.; Gambaryan, A.S.; Lvova, T.N.; Nedospasov, A.A.; Venkstern, T.V. Purification and characterization of tRNA (adenine-1-)-methyltransferase from Thermus flavus strain 71. Eur. J. Biochem. 1982, 129, 429–436. [Google Scholar] [CrossRef] [PubMed]
- De Bie, L.G.; Roovers, M.; Oudjama, Y.; Wattiez, R.; Tricot, C.; Stalon, V.; Droogmans, L.; Bujnicki, J.M. The yggH gene of Escherichia coli encodes a tRNA (m7G46) methyltransferase. J. Bacteriol. 2003, 185, 3238–3243. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, H.; Watanabe, K.; Ikeuchi, Y.; Suzuki, T.; Endo, Y.; Hori, H. Substrate tRNA recognition mechanism of tRNA (m7G46) methyltransferase from Aquifex aeolicus. J. Biol. Chem. 2004, 279, 49151–49159. [Google Scholar] [CrossRef] [PubMed]
- Tomikawa, C.; Ochi, A.; Hori, H. The C-terminal region of thermophilic tRNA (m7G46) methyltransferase (TrmB) stabilizes the dimer structure and enhances fidelity of methylation. Proteins 2008, 71, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Tomikawa, C.; Takai, K.; Hori, H. Kinetic characterization of substrate-binding sites of thermostable tRNA methyltransferase (TrmB). J. Biochem. 2018, 163, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Auxilien, S.; El Khadali, F.; Rasmussen, A.; Douthwaite, S.; Grosjean, H. Archease from Pyrococcus abyssi improves substrate specificity and solubility of a tRNA m5C methyltransferase. J. Biol. Chem. 2007, 282, 18711–18721. [Google Scholar] [CrossRef] [PubMed]
- Kuratani, M.; Hirano, M.; Goto-Ito, S.; Itoh, Y.; Hikida, Y.; Nishimoto, M.; Sekine, S.; Bessho, Y.; Ito, T.; Grosjean, H.; et al. Crystal structure of Methanocaldococcus jannaschii Trm4 complexed with sinefungin. J. Mol. Biol. 2010, 401, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Roovers, M.; Hale, C.; Tricot, C.; Terns, M.P.; Terns, R.M.; Grosjean, H.; Droogmans, L. Formation of the conserved pseudouridine at position 55 in archaeal tRNA. Nucleic Acids Res. 2006, 34, 4293–4301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurha, P.; Gupta, R. Archaeal Pus10 proteins can produce both pseudouridine 54 and 55 in tRNA. RNA 2008, 14, 2521–2527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joardar, A.; Jana, S.; Fitzek, E.; Gurha, P.; Majumder, M.; Chatterjee, K.; Geisler, M.; Gupta, R. Role of forefinger and thumb loops in production of Ψ54 and Ψ55 in tRNAs by archaeal Pus10. RNA 2013, 19, 1279–1294. [Google Scholar] [CrossRef] [PubMed]
- Kamalampeta, R.; Keffer-Wilkes, L.C.; Kothe, U. tRNA binding, positioning, and modification by the pseudouridine synthase Pus10. J. Mol. Biol. 2013, 425, 3863–3874. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Yuan, Y.A. Crystal structure of Mj1640/DUF358 protein reveals a putative SPOUT-class RNA methyltransferase. J. Mol. Cell. Biol. 2010, 2, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Wurm, J.P.; Griese, M.; Bahr, U.; Held, M.; Heckel, A.; Karas, M.; Soppa, J.; Wöhnert, J. Identification of the enzyme responsible for N1-methylation of pseudouridine 54 in archaeal tRNAs. RNA 2012, 18, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, K.; Blaby, I.K.; Thiaville, P.C.; Majumder, M.; Grosjean, H.; Yuan, Y.A.; Gupta, R.; de Crécy-Lagard, V. The archaeal COG1901/DUF358 SPOUT-methyltransferase members, together with pseudouridine synthase Pus10, catalyze the formation of 1-methylpseudouridine at position 54 of tRNA. RNA 2012, 18, 421–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davanloo, P.; Sprinzl, M.; Watanabe, K.; Albani, M.; Kersten, H. Role of ribothymidine in the thermal stability of transfer RNA as monitored by proton magnetic resonance. Nucleic Acids Res. 1979, 6, 1571–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Himeno, H.; Ohta, T. Selective utilization of 2-thioribothymidine- and ribothymidine-containing tRNAs by the protein synthetic systems of Thermus thermophilus HB 8 depending on the environmental temperature. J. Biochem. 1984, 96, 1625–1632. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Watanabe, K.; Miyazawa, T. Dynamic structures and functions of transfer ribonucleic acids from extreme thermophiles. Adv. Biophys. 1987, 23, 115–147. [Google Scholar] [CrossRef]
- Urbonavicius, J.; Skouloubris, S.; Myllykallio, H.; Grosjean, H. Identification of a novel gene encoding a flavin-dependent tRNA:m5U methyltransferase in bacteria—evolutionary implications. Nucleic Acids Res. 2005, 33, 3955–3964. [Google Scholar] [CrossRef] [PubMed]
- Cicmil, N. Crystallization and preliminary X-ray crystallographic characterization of TrmFO, a folate-dependent tRNA methyltransferase from Thermotoga maritima. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2008, 64, 193–195. [Google Scholar] [CrossRef] [PubMed]
- Nishimasu, H.; Ishitani, R.; Yamashita, K.; Iwashita, C.; Hirata, A.; Hori, H.; Nureki, O. Atomic structure of a folate/FAD-dependent tRNA T54 methyltransferase. Proc. Natl. Acad. Sci. USA 2009, 106, 8180–8185. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, R.; Yamashita, K.; Nishimasu, H.; Tomikawa, C.; Ochi, A.; Iwashita, C.; Hirata, A.; Ishitani, R.; Nureki, O.; Hori, H. The tRNA recognition mechanism of folate/FAD-dependent tRNA methyltransferase (TrmFO). J. Biol. Chem. 2012, 287, 42480–42494. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, R.; Tomikawa, C.; Shigi, N.; Kazayama, A.; Asai, S.; Takuma, H.; Hirata, A.; Fourmy, D.; Asahara, H.; Watanabe, K.; et al. The folate/FAD-dependent tRNA methyltransferase (TrmFO) from Thermus thermophilus regulates the other modifications in tRNA at low temperatures. Genes Cells 2016, 21, 740–754. [Google Scholar] [CrossRef] [PubMed]
- Hamdane, D.; Grosjean, H.; Fontecave, M. Flavin-Dependent Methylation of RNAs: Complex Chemistry for a Simple Modification. J. Mol. Biol. 2016, 428, 4867–4881. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, R.; Miyake, R.; Fukumoto, A.; Nakashima, M.; Hori, H. Consumption of N5, N10-methylenetetrahydrofolate in Thermus thermophilus under nutrient-poor condition. J. Biochem. 2018, 164, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Shigi, N.; Suzuki, T.; Tamakoshi, M.; Oshima, T.; Watanabe, K. Conserved bases in the TPsi C loop of tRNA are determinants for thermophile-specific 2-thiouridylation at position 54. J. Biol. Chem. 2002, 277, 39128–39135. [Google Scholar] [CrossRef] [PubMed]
- Shigi, N.; Suzuki, T.; Terada, T.; Shirouzu, M.; Yokoyama, S.; Watanabe, K. Temperature-dependent biosynthesis of 2-thioribothymidine of Thermus thermophilus tRNA. J. Biol. Chem. 2006, 281, 2104–2113. [Google Scholar] [CrossRef] [PubMed]
- Shigi, N.; Sakaguchi, Y.; Suzuki, T.; Watanabe, K. Identification of two tRNA thiolation genes required for cell growth at extremely high temperatures. J. Biol. Chem. 2006, 281, 14296–14306. [Google Scholar] [CrossRef] [PubMed]
- Shigi, N.; Sakaguchi, Y.; Asai, S.; Suzuki, T.; Watanabe, K. Common thiolation mechanism in the biosynthesis of tRNA thiouridine and sulphur-containing cofactors. EMBO J. 2008, 27, 3267–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigi, N. Posttranslational modification of cellular proteins by a ubiquitin-like protein in bacteria. J. Biol. Chem. 2012, 287, 17568–17577. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Kuratani, M.; Goto-Ito, S.; Ito, T.; Katsura, K.; Terada, T.; Shirouzu, M.; Sekine, S.; Shigi, N.; Yokoyama, S. Crystallographic and mutational studies on the tRNA thiouridine synthetase TtuA. Proteins 2013, 81, 1232–1244. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Narai, S.; Omura, N.; Shigi, N.; Chimnaronk, S.; Tanaka, Y.; Yao, M. Crystallographic study of the 2-thioribothymidine-synthetic complex TtuA-TtuB from Thermus thermophilus. Acta Crystallogr. F Struct. Biol. Commun. 2016, 72, 777–781. [Google Scholar] [CrossRef] [PubMed]
- Shigi, N.; Asai, S.-I.; Watanabe, K. Identification of a rhodanese-like protein involved in thiouridine biosynthesis in Thermus thermophilus tRNA. FEBS Lett. 2016, 590, 4628–4637. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Asai, S.-I.; Narai, S.; Nambu, S.; Omura, N.; Sakaguchi, Y.; Suzuki, T.; Ikeda-Saito, M.; Watanabe, K.; Yao, M.; et al. Biochemical and structural characterization of oxygen-sensitive 2-thiouridine synthesis catalyzed by an iron-sulfur protein TtuA. Proc. Natl. Acad. Sci. USA 2017, 114, 4954–4959. [Google Scholar] [CrossRef] [PubMed]
- Urbonavicius, J.; Auxilien, S.; Walbott, H.; Trachana, K.; Golinelli-Pimpaneau, B.; Brochier-Armanet, C.; Grosjean, H. Acquisition of a bacterial RumA-type tRNA(uracil-54, C5)-methyltransferase by Archaea through an ancient horizontal gene transfer. Mol. Microbiol. 2008, 67, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Walbott, H.; Leulliot, N.; Grosjean, H.; Golinelli-Pimpaneau, B. The crystal structure of Pyrococcus abyssi tRNA (uracil-54, C5)-methyltransferase provides insights into its tRNA specificity. Nucleic Acids Res. 2008, 36, 4929–4940. [Google Scholar] [CrossRef] [PubMed]
- Shigi, N. Biosynthesis and functions of sulfur modifications in tRNA. Front. Genet. 2014, 5, 67. [Google Scholar] [CrossRef] [PubMed]
- Gurha, P.; Joardar, A.; Chaurasia, P.; Gupta, R. Differential roles of archaeal box H/ACA proteins in guide RNA-dependent and independent pseudouridine formation. RNA Biol. 2007, 4, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Fourmann, J.B.; Loegler, C.; Charpentier, B.; Branlant, C. Identification of determinants in the protein partners aCBF5 and aNOP10 necessary for the tRNA: Psi55-synthase and RNA-guided RNA: Psi-synthase activities. Nucleic Acids Res. 2007, 35, 5610–5624. [Google Scholar] [CrossRef] [PubMed]
- Kamalampeta, R.; Kothe, U. Archaeal proteins Nop10 and Gar1 increase the catalytic activity of Cbf5 in pseudouridylating tRNA. Sci. Rep. 2012, 2, 663. [Google Scholar] [CrossRef] [PubMed]
- Nurse, K.; Wrzesinski, J.; Bakin, A.; Lane, B.G.; Ofengand, J. Purification, cloning, and properties of the tRNA psi 55 synthase from Escherichia coli. RNA 1995, 1, 102–112. [Google Scholar] [PubMed]
- Pan, H.; Agarwalla, S.; Moustakas, D.T.; Finer-Moore, J.; Stroud, R.M. Structure of tRNA pseudouridine synthase TruB and its RNA complex: RNA recognition through a combination of rigid docking and induced fit. Proc. Natl. Acad. Sci. USA 2003, 100, 12648–12653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wouters, J.; Tricot, C.; Durbecq, V.; Roovers, M.; Stalon, V.; Droogmans, L. Preliminary X-ray crystallographic analysis of tRNA pseudouridine 55 synthase from the thermophilic eubacterium Thermotoga maritima. Acta Crystallogr. D Biol. Crystallogr. 2003, 59, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Phannachet, K.; Huang, R.H. Conformational change of pseudouridine 55 synthase upon its association with RNA substrate. Nucleic Acids Res. 2004, 32, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Phannachet, K.; Elias, Y.; Huang, R.H. Dissecting the roles of a strictly conserved tyrosine in substrate recognition and catalysis by pseudouridine 55 synthase. Biochemistry 2005, 44, 15488–15494. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Kunibayashi, T.; Tomikawa, C.; Ochi, A.; Kanai, T.; Hirata, A.; Iwashita, C.; Hori, H. Pseudouridine at position 55 in tRNA controls the contents of other modified nucleotides for low-temperature adaptation in the extreme-thermophilic eubacterium Thermus thermophilus. Nucleic Acids Res. 2011, 39, 2304–2318. [Google Scholar] [CrossRef] [PubMed]
- Renalier, M.H.; Joseph, N.; Gaspin, C.; Thebault, P.; Mougin, A. The Cm56 tRNA modification in archaea is catalyzed either by a specific 2′-O-methylase, or a C/D sRNP. RNA 2005, 11, 1051–1063. [Google Scholar] [CrossRef] [PubMed]
- Tkaczuk, K.L.; Dunin-Horkawicz, S.; Purta, E.; Bujnicki, J.M. Structural and evolutionary bioinformatics of the SPOUT superfamily of methyltransferases. BMC Bioinfor. 2007, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Kuratani, M.; Bessho, Y.; Nishimoto, M.; Grosjean, H.; Yokoyama, S. Crystal structure and mutational study of a unique SpoU family archaeal methylase that forms 2′-O-methylcytidine at position 56 of tRNA. J. Mol. Biol. 2008, 375, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.T. Mycoplasma evolution: A review of the use of ribosomal and transfer RNA nucleotide sequences in the determination of phylogenetic relationships. Yale J. Biol. Med. 1983, 56, 367–372. [Google Scholar] [PubMed]
- Yamaizumi, Z.; Ihara, M.; Kuchino, Y.; Gupta, R.; Woese, C.R.; Nishimura, S. Archaebacterial tRNA contains 1-methylinosine at residue 57 in T psi C-loop. Nucleic Acids Symp. Ser. 1982, 11, 209–213. [Google Scholar]
- Grosjean, H.; Constantinesco, F.; Foiret, D.; Benachenhou, N. A novel enzymatic pathway leading to 1-methylinosine modification in Haloferax volcanii tRNA. Nucleic Acids Res. 1995, 23, 4312–4319. [Google Scholar] [CrossRef] [PubMed]
- Roovers, M.; Wouters, J.; Bujnicki, J.M.; Tricot, C.; Stalon, V.; Grosjean, H.; Droogmans, L. A primordial RNA modification enzyme: The case of tRNA (m1A) methyltransferase. Nucleic Acids Res. 2004, 32, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Guelorget, A.; Roovers, M.; Guérineau, V.; Barbey, C.; Li, X.; Golinelli-Pimpaneau, B. Insights into the hyperthermostability and unusual region-specificity of archaeal Pyrococcus abyssi tRNA m1A57/58 methyltransferase. Nucleic Acids Res. 2010, 38, 6206–6218. [Google Scholar] [CrossRef] [PubMed]
- Guelorget, A.; Barraud, P.; Tisné, C.; Golinelli-Pimpaneau, B. Structural comparison of tRNA m(1)A58 methyltransferases revealed different molecular strategies to maintain their oligomeric architecture under extreme conditions. BMC Struct. Biol. 2011, 11, 48. [Google Scholar] [CrossRef] [PubMed]
- Hamdane, D.; Guelorget, A.; Guérineau, V.; Golinelli-Pimpaneau, B. Dynamics of RNA modification by a multi-site-specific tRNA methyltransferase. Nucleic Acids Res. 2014, 42, 11697–11706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biou, V.; Yaremchuk, A.; Tukalo, M.; Cusack, S. The 2.9 A crystal structure of T. thermophilus seryl-tRNA synthetase complexed with tRNA(Ser). Science 1994, 263, 1404–1410. [Google Scholar] [CrossRef] [PubMed]
- Droogmans, L.; Roovers, M.; Bujnicki, J.M.; Tricot, C.; Hartsch, T.; Stalon, V.; Grosjean, H. Cloning and characterization of tRNA (m1A58) methyltransferase (TrmI) from Thermus thermophilus HB27, a protein required for cell growth at extreme temperatures. Nucleic Acids Res. 2003, 31, 2148–2156. [Google Scholar] [CrossRef] [PubMed]
- Barraud, P.; Golinelli-Pimpaneau, B.; Atmanene, C.; Sanglier, S.; Van Dorsselaer, A.; Droogmans, L.; Dardel, F.; Tisné, C. Crystal structure of Thermus thermophilus tRNA m1A58 methyltransferase and biophysical characterization of its interaction with tRNA. J. Mol. Biol. 2008, 377, 535–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuratani, M.; Yanagisawa, T.; Ishii, R.; Matsuno, M.; Si, S.Y.; Katsura, K.; Ushikoshi-Nakayama, R.; Shibata, R.; Shirouzu, M.; Bessho, Y.; et al. Crystal structure of tRNA m1A58 methyltransferase TrmI from Aquifex aeolicus in complex with S-adenosyl-L-methionine. J. Struct. Funct. Genomics 2014, 15, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Takuma, H.; Ushio, N.; Minoji, M.; Kazayama, A.; Shigi, N.; Hirata, A.; Ochi, A.; Hori, H. Substrate tRNA recognition mechanism of eubacterial tRNA (m1A58) methyltransferase (TrmI). J. Biol. Chem. 2015, 290, 5912–5925. [Google Scholar] [CrossRef] [PubMed]
- Dégut, C.; Ponchon, L.; Folly-Klan, M.; Barraud, P.; Tisné, C. The m1A58 modification in eubacterial tRNA: An overview of tRNA recognition and mechanism of catalysis by TrmI. Biophys. Chem. 2016, 210, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Väre, V.Y.; Eruysal, E.R.; Narendran, A.; Sarachan, K.L.; Agris, P.F. Chemical and Conformational Diversity of Modified Nucleosides Affects tRNA Structure and Function. Biomolecules 2017, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Motorin, Y.; Helm, M. tRNA stabilization by modified nucleotides. Biochemistry 2010, 49, 4934–4944. [Google Scholar] [CrossRef] [PubMed]
- El Yacoubi, B.; Bailly, M.; de Crécy-Lagard, V. Biosynthesis and function of posttranscriptional modifications of transfer RNAs. Annu. Rev. Genet. 2012, 46, 69–95. [Google Scholar] [CrossRef] [PubMed]
- Manickam, N.; Joshi, K.; Bhatt, M.J.; Farabaugh, P.J. Effects of tRNA modification on translational accuracy depend on intrinsic codon-anticodon strength. Nucleic Acids Res. 2016, 44, 1871–1881. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, H.; Westhof, E. An integrated, structure- and energy-based view of the genetic code. Nucleic Acids Res. 2016, 44, 8020–8040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agris, P.F.; Eruysal, E.R.; Narendran, A.; Väre, V.Y.P.; Vangaveti, S.; Ranganathan, S.V. Celebrating wobble decoding: Half a century and still much is new. RNA Biol. 2017, 16, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Perret, V.; Garcia, A.; Grosjean, H.; Ebel, J.P.; Florentz, C.; Giegé, R. Relaxation of a transfer RNA specificity by removal of modified nucleotides. Nature 1990, 344, 787–789. [Google Scholar] [CrossRef] [PubMed]
- Aström, S.U.; Byström, A.S. Rit1, a tRNA backbone-modifying enzyme that mediates initiator and elongator tRNA discrimination. Cell 1994, 79, 535–546. [Google Scholar] [CrossRef]
- Kaneko, T.; Suzuki, T.; Kapushoc, S.T.; Rubio, M.A.; Ghazvini, J.; Watanabe, K.; Simpson, L.; Suzuki, T. Wobble modification differences and subcellular localization of tRNAs in Leishmania tarentolae: Implication for tRNA sorting mechanism. EMBO J. 2003, 22, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Phan, L.; Cuesta, R.; Carison, B.A.; Pak, M.; Asano, K.; Björk, G.R.; Tamame, M.; Hinnebusch, A.G. The essential Gcd10p-Gcd14p nuclear complex is required for 1-methyladenosine modification and maturation of initiator methionyl-tRNA. Genes Dev. 1998, 12, 3650–3652. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Phan, L.; Hinnebusch, A.G. The Gcd10p/Gcd14p complex is the essential two-subunit tRNA(1-methyladenosine) methyltransferase of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2000, 97, 5173–5178. [Google Scholar] [CrossRef] [PubMed]
- Ohira, T.; Suzuki, T. Retrograde nuclear import of tRNA precursors is required for modified base biogenesis in yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 10502–10507. [Google Scholar] [CrossRef] [PubMed]
- Ohira, T.; Suzuki, T. Precursors of tRNAs are stabilized by methylguanosine cap structures. Nat. Chem. Biol. 2016, 12, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, K.; Nostramo, R.T.; Wan, Y.; Hopper, A.K. tRNA dynamics between the nucleus, cytoplasm and mitochondrial surface: Location, location, location. Biochim. Biophys. Acta 2018, 1861, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Kadaba, S.; Krueger, A.; Trice, T.; Krecic, A.M.; Hinnebusch, A.G.; Anderson, J. Nuclear surveillance and degradation of hypomodified initiator tRNAMet in S. cerevisiae. Genes Dev. 2004, 18, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Alexandrov, A.; Chernyakov, I.; Gu, W.; Hiley, S.L.; Hughes, T.R.; Grayhack, E.J.; Phizicky, E.M. Rapid tRNA decay can result from lack of nonessential modifications. Mol. Cell 2006, 21, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Phizicky, E.M.; Hopper, A.K. tRNA biology charges to the front. Genes Dev. 2010, 24, 1832–1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, J.M.; Okada, N.; Tobe, T.; Watarai, M.; Fukuda, I.; Suzuki, T.; Nakata, N.; Komatsu, K.; Yoshikawa, M.; Sasakawa, C. vacC, a virulence-associated chromosomal locus of Shigella flexneri, is homologous to tgt, a gene encoding tRNA-guanine transglycosylase (Tgt) of Escherichia coli K-12. J. Bacteriol. 1994, 176, 4627–4634. [Google Scholar] [CrossRef] [PubMed]
- Saga, A.E.; Vasil, A.I.; Vasil, M.L. Molecular characterization of mutants affected in the osmoprotectant-dependent induction of phspholipase C in Pseudomonas aeruginosa PAO1. Mol. Microbiol. 1997, 23, 43–56. [Google Scholar] [CrossRef]
- Takano, Y.; Takayanagi, N.; Hori, H.; Ikeuchi, Y.; Suzuki, T.; Kimura, A.; Okuno, T. A gene involved in modifying transfer RNA is required for fungal pathogenicity and stress tolerance of Colletotrichum lagenarium. Mol. Microbiol. 2006, 60, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Sleiman, D.; Goldschmidt, V.; Barraud, P.; Marquet, R.; Paillart, J.C.; Tisné, C. Initiation of HIV-1 reverse transcription and functional role of nucleocapsid-mediated tRNA/viral genome interactions. Virus Res. 2012, 169, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Saadatmand, J.; Kleiman, L. Aspects of HIV-1 assembly that promote primer tRNA (Lys3) annealing to viral RNA. Virus Res. 2012, 169, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Gehrig, S.; Eberle, M.-E.; Botschen, F.; Rimbach, K.; Eberle, F.; Eigenbrod, T.; Kaiser, S.; Holmes, W.M.; Erdmann, V.A.; Sprinzl, M.; et al. Identification of modifications in microbial, native tRNA that suppress immunostimulatory activity. J. Exp. Med. 2012, 209, 225–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jöckel, S.; Nees, G.; Sommer, R.; Zhao, Y.; Cherkasov, D.; Hori, H.; Ehm, G.; Schnare, M.; Nain, M.; Kaufmann, A.; Bauer, S. The 2′-O-methylation status of a single guanosine controls transfer RNA-mediated Toll-like receptor 7 activation or inhibition. J. Exp. Med. 2012, 209, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, F.C.F.; Freund, I.; Weigand, M.A.; Helm, M.; Dalpke, A.H.; Eigenbrod, T. Identification of an optimized 2′-O-methylated trinucleotide RNA motif inhibiting Toll-like receptors 7 and 8. RNA 2017, 23, 1344–1351. [Google Scholar] [CrossRef] [PubMed]
- Keller, P.; Freund, I.; Marchand, V.; Bec, G.; Huang, R.; Motorin, Y.; Eigenbrod, T.; Dalpke, A.; Helm, M. Double methylation of tRNA-U54 to 2′-O-methylthymidine (Tm) synergistically decreases immune response by Toll-like receptor 7. Nucleic Acids Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.R.; Nicoghosian, K.; Cedergren, R.J.; Wong, J.T. Sequences of halobacterial tRNAs and the paucity of U in the first position of their anticodons. Nucleic Acids Res. 1983, 11, 5433–5442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosjean, H.; Gaspin, C.; Marck, C.; Decatur, W.A.; de Crecy-Lagard, V. RNomics and Modomics in the halophilic archaea Haloferax volcanii: Identification of RNA modification genes. BMC Genom. 2008, 9, 470. [Google Scholar] [CrossRef] [PubMed]
- Phillips, G.; de Crécy-Lagard, V. Biosynthesis and function of tRNA modifications in Archaea. Curr. Opin. Microbiol. 2011, 14, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, H.; Gupta, R.; Maxwell, S. Modified nucleotides in arhcaeal RNAs. In Archaea: New Models for Prokaryotic Biology; Blum, P., Ed.; Caister Academic Press: Portland, OR, USA, 2008; pp. 171–196. [Google Scholar]
- Rintala-Dempsey, A.C.; Kothe, U. Eukaryotic stand-alone pseudouridine synthases–RNA modifying enzymes and emerging regulators of gene expression? RNA Biol. 2017, 14, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Hori, H. Methylated nucleosides in tRNA and tRNA methyltransferases. Front. Genet. 2014, 5, 144. [Google Scholar] [CrossRef] [PubMed]
- Grosjean, H.; Oshima, T. How nucleic acids cope with high temperature. In Physiology and Biochemistry of Extremophiles; Gerday, C., Glansdorff, N., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 39–56. [Google Scholar]
- Kumazawa, Y.; Yokogawa, T.; Tsurui, H.; Miura, K.; Watanabe, K. Effect of the higher-order structure of tRNAs on the stability of hybrids with oligodeoxyribonucleotides: Separation of tRNA by an efficient solution hybridization. Nucleic Acids Res. 1992, 20, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Tsurui, H.; Kumazawa, Y.; Sanokawa, R.; Watanabe, Y.; Kuroda, T.; Wada, A.; Watanabe, K.; Shirai, T. Batchwise purification of specific tRNAs by a solid-phase DNA probe. Anal. Biochem. 1994, 221, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Ueda, T.; Watanabe, K. A new method for identifying the amino acid attached to a particular RNA in the cell. FEBS Lett. 1996, 381, 195–198. [Google Scholar] [CrossRef] [Green Version]
- Yokogawa, T.; Kitamura, Y.; Nakamura, D.; Ohno, S.; Nishikawa, K. Optimization of the hybridization-based method for purification of thermostable tRNAs in the presence of tetraalkylammonium salts. Nucleic Acids Res. 2010, 38, e89. [Google Scholar] [CrossRef] [PubMed]
- Kazayama, A.; Yamagami, R.; Yokogawa, T.; Hori, H. Improved solid-phase DNA probe method for tRNA purification: Large-scale preparation and alteration of DNA fixation. J. Biochem. 2015, 157, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Stanley, J.; Vassilenko, S. A different approach to RNA sequencing. Nature 1978, 274, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Kuchino, Y.; Kato, M.; Sugisaki, H.; Nishimura, S. Nucleotide sequence of starfish initiator tRNA. Nucleic Acids Res. 1979, 6, 3459–3469. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Ikeuchi, Y.; Noma, A.; Suzuki, T.; Sakaguchi, Y. Mass spectrometric identification and characterization of RNA-modifying enzymes. Methods Enzymol. 2007, 425, 211–229. [Google Scholar] [PubMed]
- Suzuki, T.; Suzuki, T. A complete landscape of post-transcriptional modifications in mammalian mitochondrial tRNAs. Nucleic Acids Res. 2014, 42, 7346–7357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengel-Jorgensen, J.; Kirpekar, F. Detection of pseudouridine and other modifications in tRNA by cyanoethylation and MALDI mass spectrometry. Nucleic Acids Res 2002, 30, e135. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Huang, B.; Anderson, J.T.; Byström, A.S. Unexpected accumulation of ncm(5)U and ncm(5)S(2) (U) in a trm9 mutant suggests an additional step in the synthesis of mcm(5)U and mcm(5)S(2)U. PLoS ONE 2011, 6, e20783. [Google Scholar]
- Agris, P.F.; Koh, H.; Söll, D. The effect of growth temperatures on the in vivo ribose methylation of Bacillus stearothermophilus transfer RNA. Arch Biochem Biophys. 1973, 154, 277–282. [Google Scholar] [CrossRef]
- Andachi, Y.; Yamao, F.; Muto, A.; Osawa, S. Codon recognition patterns as deduced from sequences of the complete set of transfer RNA species in Mycoplasma capricolum. Resemblance to mitochondria. J. Mol. Biol. 1989, 209, 37–54. [Google Scholar] [CrossRef]
- Matsugi, J.; Jia, H.T.; Murao, K.; Ishikura, H. Nucleotide sequences of serine tRNAs from Bacillus subtilis. Biochem. Biophys. Acta 1992, 1130, 333–335. [Google Scholar] [CrossRef]
- Hori, H. Transfer RNA methyltransferases with a SpoU-TrmD (SPOUT) fold and their modified nucleosides in tRNA. Biomolecules 2017, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Reddy, D.M.; Crain, P.F.; Edmonds, C.G.; Gupta, R.; Hashizume, T.; Stetter, K.O.; Widdel, F.; McCloskey, J.A. Structure determination of two new amino acid-containing derivatives of adenosine from tRNA of thermophilic bacteria and archaea. Nucleic Acids Res. 1992, 20, 5607–5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusack, S.; Yaremchuk, A.; Krikliviy, I.; Tukalo, M. tRNAPro anticodon recognition by Thermus thermophilus prolyl-tRNA synthetase. Structure 1998, 6, 101–108. [Google Scholar] [CrossRef]
- Li, F.; Dong, J.; Hu, X.; Gong, W.; Li, J.; Shen, J.; Tian, H.; Wang, J. A covalent approach for site-specific RNA labeling in Mammalian cells. Angew. Chem. Int. Ed. Engl. 2015, 54, 4597–4602. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Gurha, P.; Tran, E.J.; Maxwell, E.S.; Gupta, R. Sequential 2′-O-methylation of archaeal pre-tRNATrp nucleotides is guided by the intron-encoded but trans-acting box C/D ribonucleoprotein of pre-tRNA. J. Biol. Chem. 2004, 279, 47661–47671. [Google Scholar] [CrossRef] [PubMed]
- Haas, E.S.; Daniels, C.J.; Reeve, J.N. Genes encoding 5S rRNA and tRNAs in the extremely thermophilic archaebacterium Methanothermus fervidus. Gene 1989, 77, 253–263. [Google Scholar] [CrossRef]
- Randau, L.; Münch, R.; Hohn, M.J.; Jahn, D.; Söll, D. Nanoarchaeum equitans creates functional tRNAs from separate genes for their 5′- and 3′-halves. Nature 2005, 433, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Randau, L.; Pearson, M.; Söll, D. The complete set of tRNA species in Nanoarchaeum equitans. FEBS Lett. 2005, 579, 2945–2947. [Google Scholar] [CrossRef] [PubMed]
- Richter, H.; Mohr, S.; Randau, L. C/D box sRNA, CRISPR RNA and tRNA processing in an archaeon with a minimal fragmented genome. Biochem. Soc. Trans. 2013, 41, 411–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardin, J.W.; Reyes, F.E.; Batey, R.T. Analysis of a critical interaction within the archaeal box C/D small ribonucleoprotein complex. J. Biol. Chem. 2009, 284, 15317–15324. [Google Scholar] [CrossRef] [PubMed]
- McCloskey, J.A.; Crain, P.F.; Edmonds, C.G.; Gupta, R.; Hashizume, T.; Phillipson, D.W.; Stetter, K.O. Structure determination of a new fluorescent tricyclic nucleoside from archaebacterial tRNA. Nucleic Acids Res. 1987, 15, 683–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushida, C.; Muramatsu, T.; Mizushima, H.; Ueda, T.; Watanabe, K.; Stetter, K.O.; Crain, P.F.; McCloskey, J.A.; Kuchino, Y. Structural feature of the initiator tRNA gene from Pyrodictium occultum and the thermal stability of its gene product, tRNA(imet). Biochimie 1996, 79, 847–855. [Google Scholar] [CrossRef]
- McCloskey, J.A.; Liu, X.H.; Crain, P.F.; Bruenger, E.; Guymon, R.; Hashizume, T.; Stetter, K.O. Posttranscriptional modification of transfer RNA in the submarine hyperthermophile Pyrolobus fumarii. Nucleic Acids Symp. Ser. 2000, 44, 267–268. [Google Scholar] [CrossRef]
- Zhou, S.; Sitaramaiah, D.; Noon, K.R.; Guymon, R.; Hashizume, T.; McCloskey, J.A. Structures of two new “minimalist” modified nucleosides from archaeal tRNA. Bioorg. Chem. 2004, 32, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Lin, J.; Ye, K. Box C/D guide RNAs recognize a maximum of 10 nt of substrates. Proc. Natl. Acad. Sci. USA 2016, 27, 10878–10883. [Google Scholar] [CrossRef] [PubMed]
- Agari, Y.; Sato, S.; Wakamatsu, T.; Bessho, Y.; Ebihara, A.; Yokoyama, S.; Kuramitsu, S.; Shinkai, A. X-ray crystal structure of a hypothetical Sua5 protein from Sulfolobus tokodaii strain 7. Proteins 2008, 70, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Kuratani, M.; Kasai, T.; Akasaka, R.; Higashijima, K.; Terada, T.; Kigawa, T.; Shinkai, A.; Bessho, Y.; Yokoyama, S. Crystal structure of Sulfolobus tokodaii Sua5 complexed with L-threonine and AMPPNP. Proteins 2011, 79, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Parthier, C.; Goerlich, S.; Jaenecke, F.; Breithaupt, C.; Brauer, U.; Fandrich, U.; Clausnitzer, D.; Wehmeier, U.F.; Bottcher, C.; Scheel, D.; et al. The O-carbamoyltransferase TobZ catalyzes an ancient enzymatic reaction. Angew. Chem. Int. Ed. Engl. 2012, 51, 4046–4052. [Google Scholar] [CrossRef] [PubMed]
- Klenk, H.P.; Schwass, V.; Zillig, W. Nucleotide sequence of the genes encoding proline tRNA(UGG) and threonine tRNA(GGU) and consensus promoter model of Thermococcus celer. Biochim. Biophys. Acta 1993, 1172, 236–238. [Google Scholar] [CrossRef]
- Watanabe, K.; Oshima, T.; Saneyoshi, M.; Nishimura, S. Replacement of ribothymidine by 5-methyl-2-thiouridine in sequence GT psi C in tRNA of an extreme thermophile. FEBS Lett. 1974, 43, 59–63. [Google Scholar] [CrossRef]
- Dalluge, J.J.; Hashizume, T.; Sopchik, A.E.; McCloskey, J.A.; Davis, D.R. Conformational flexibility in RNA: The role of dihydrouridine. Nucleic Acids Res. 1996, 24, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Sokołowski, M.; Klassen, R.; Bruch, A.; Schaffrath, R.; Glatt, S. Cooperativity between different tRNA modifications and their modification pathways. Biochem. Biophys. Acta 2018, 1861, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.J.; Zhou, M.; Fang, Z.P.; Wang, M.; Zhou, X.L.; Wang, E.D. The tRNA recognition mechanism of the minimalist SPOUT methyltransferase, TrmL. Nucleic Acids Res. 2013, 41, 7828–7842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, A.J.; Swairjo, M.A.; Schimmel, P. Structure-specific tRNA-binding protein from the extreme thermophile Aquifex aeolicus. EMBO J. 1999, 18, 3475–3483. [Google Scholar] [CrossRef] [PubMed]
- Swairjo, M.A.; Morales, A.J.; Wang, C.C.; Ortiz, A.R.; Schimmel, P. Crystal structure of trbp111: A structure-specific tRNA-binding protein. EMBO J. 2000, 19, 6278–6298. [Google Scholar] [CrossRef] [PubMed]
- Kushiro, T.; Schimmel, P. Trbp111 selectively binds a noncovalently assembled tRNA-like structure. Proc. Natl. Acad. Sci. USA 2002, 99, 16631–16635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, K.K.; Cheng, C.L.; Bingman, C.A.; Phillips, G.N., Jr.; Raines, R.T. A tRNA splicing operon: Archease endows RtcB with dual GTP/ATP cofactor specificity and accelerates RNA ligation. Nucleic Acids Res. 2014, 42, 3931–3942. [Google Scholar] [CrossRef] [PubMed]
- Popow, J.; Jurkin, J.; Schleiffer, A.; Martinez, J. Analysis of orthologous groups reveals archease and DDX1 as tRNA splicing factors. Nature 2014, 511, 104–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maraia, R.J.; Arimbasseri, A.G. Factors That Shape Eukaryotic tRNAomes: Processing, Modification and Anticodon-Codon Use. Biomolecules 2017, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Blewett, N.H.; Maraia, R.J. La involvement in tRNA and other RNA processing events including differences among yeast and other eukaryotes. Biochim. Biophys. Acta 2018, 1861, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Gutgsell, N.; Englund, N.; Niu, L.; Kaya, Y.; Lane, B.G.; Ofengand, J. Deletion of the Escherichia coli pseudouridine synthase gene truB blocks formation of pseudouridine 55 in tRNA in vivo, does not affect exponential growth, but confers a strong selective disadvantage in competition with wild-type cells. RNA 2000, 6, 1870–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keffer-Wilkes, L.C.; Veerareddygari, G.R.; Kothe, U. RNA modification enzyme TruB is a tRNA chaperone. Proc. Natl. Acad. Sci. USA 2016, 113, 14306–14311. [Google Scholar] [CrossRef] [PubMed]
- Quigley, G.J.; Teeter, M.M.; Rich, A. Structural analysis of spermine and magnesium ion binding to yeast phenylalanine transfer RNA. Proc. Natl. Acad. Sci. USA 1978, 75, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Frydman, B.; de los Santos, C.; Frydman, R.B. A 13C NMR study of [5,8-13C2]spermidine binding to tRNA and to Escherichia coli macromolecules. J. Biol. Chem. 1990, 265, 20874–20878. [Google Scholar] [PubMed]
- Terui, Y.; Ohnuma, M.; Hiraga, K.; Kawashima, E.; Oshima, T. Stabilization of nucleic acids by unusual polyamines produced by an extreme thermophile, Thermus thermophilus. Biochem. J. 2005, 388, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Ouameur, A.A.; Bourassa, P.; Tajmir-Riahi, H.A. Probing tRNA interaction with biogenic polyamines. RNA 2010, 16, 1968–1979. [Google Scholar] [CrossRef] [PubMed]
- Oshima, T. Unique polyamines produced by an extreme thermophiles, Thermus thermophilus. Amino Acids 2007, 33, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Oshima, T.; Moriya, T.; Terui, Y. Identification, chemical synthesis, and biological functions of unusual polyamines produced by extreme thermophiles. Methods Mol. Biol. 2011, 720, 81–111. [Google Scholar] [PubMed]
- Hamana, K.; Niitsu, M.; Samejima, K.; Matsuzaki, S. Polyamine distributions in thermophilic eubacteria belonging to Thermus and Acidothermus. J. Biochem. 1991, 109, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Hamana, K.; Tanaka, T.; Hosoya, R.; Niitsu, M.; Itoh, T. Cellular polyamines of the acidophilic, thermophilic and thermoacidophilic archaebacteria, Acidilobus, Ferroplasma, Pyrobaculum, Pyrococcus, Staphylothermus, Thermococcus, Thermodiscus and Vulcanisaeta. J. Gen. Appl. Microbiol. 2003, 49, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Hayrapetyan, A.; Grosjean, H.; Helm, M. Effect of a quaternary pentamine on RNA stabilization and enzymatic methylation. Biol. Chem. 2009, 390, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, M.; Yamagami, R.; Tomikawa, C.; Ochi, Y.; Moriya, T.; Asahara, H.; Fourmy, D.; Yoshizawa, S.; Oshima, T.; Hori, H. Long and branched polyamines are required for maintenance of the ribosome, tRNAHis and tRNATyr in Thermus thermophilus cells at high temperatures. Genes Cells 2017, 22, 628–645. [Google Scholar] [CrossRef] [PubMed]
- Yue, D.; Kintanar, A.; Horowitz, J. Nucleoside modifications stabilize Mg2+ binding in Escherichia coli tRNA(Val): An imino proton NMR investigation. Biochemistry 1994, 33, 8905–8911. [Google Scholar] [CrossRef] [PubMed]
- Agris, P.F. The importance of being modified: Roles of modified nucleosides and Mg2+ in RNA structure and function. Prog. Nucleic Acid Res. Mol. Biol. 1996, 53, 79–129. [Google Scholar] [PubMed]
- Nobles, K.N.; Yarian, C.S.; Liu, G.; Guenther, R.H.; Agris, P.F. Highly conserved modified nucleosides influence Mg2+-dependent tRNA folding. Nucleic Acids Res. 2002, 30, 4751–4760. [Google Scholar] [CrossRef] [PubMed]
- Hensel, R.; Konig, H. Thermoadaptation of metanogenic bacteria by interacellular ion concentration. FEMS Microbiol. Lett. 1988, 49, 75–79. [Google Scholar] [CrossRef]
- Nawrot, B.; Sochacka, E.; Duchler, M. tRNA structural and functional changes induced by oxidative stress. Cell Mol. Life Sci. 2011, 68, 4023–4032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, M.A.; D’Silva, S.; Kon, Y.; Phizicky, E.M. tRNAHis 5-methylcytidine levels increase in response to several growth arrest conditions in Saccharomyces cerevisiae. RNA 2013, 19, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Endres, L.; Dedon, P.C.; Begley, T.J. Codon-biased translation can be regulated by wobble-base tRNA modification systems during cellular stress responses. RNA Biol. 2015, 12, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.T.; Deng, W.; Li, F.; DeMott, M.S.; Babu, I.R.; Begley, T.J.; Dedon, P.C. Highly Predictive Reprogramming of tRNA Modifications Is Linked to Selective Expression of Codon-Biased Genes. Chem. Res. Toxicol. 2015, 28, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Polikarpov, I.; Prade, R.; Caldana, C.; Paes Leme, A.F.; Mercadante, A.Z.; Riaño-Pachón, D.M.; Squina, F.M. Thermal adaptation strategies of the extremophile bacterium Thermus filiformis based on multi-omics analysis. Extremophiles 2017, 21, 775–788. [Google Scholar]
- Kimura, S.; Suzuki, T. Iron-sulfur proteins responsible for RNA modifications. Biochim. Biophys. Acta 2015, 1853, 1272–1283. [Google Scholar] [CrossRef] [PubMed]
- Ramamurthy, V.; Swann, S.L.; Spedaliere, C.J.; Mueller, E.G. Role of cysteine residues in pseudouridine synthases of different families. Biochemistry 1999, 38, 13106–13111. [Google Scholar] [CrossRef] [PubMed]
- Urbonavicius, J.; Jäger, G.; Björk, G.R. Amino acid residues of the Escherichia coli tRNA(m5U54)methyltransferase (TrmA) critical for stability, covalent binding of tRNA and enzymatic activity. Nucleic Acids Res. 2007, 35, 3297–3305. [Google Scholar] [CrossRef] [PubMed]
- Alian, A.; Lee, T.T.; Griner, S.L.; Stroud, R.M.; Finer-Moore, J. Structure of a TrmA-RNA complex: A consensus RNA fold contributes to substrate selectivity and catalysis in m5U methyltransferases. Proc. Natl. Acad. Sci. USA 2008, 105, 6876–6881. [Google Scholar] [CrossRef] [PubMed]
- Byrne, R.T.; Jenkins, H.T.; Peters, D.T.; Whelan, F.; Stowell, J.; Aziz, N.; Kasatsky, P.; Rodnina, M.V.; Koonin, E.V.; Konevega, A.L.; et al. Major reorientation of tRNA substrates defines specificity of dihydrouridine synthases. Proc. Natl. Acad. Sci. USA 2015, 112, 6033–6037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryals, J.; Hsu, R.Y.; Lipsett, M.N.; Bremer, H. Isolation of single-site Escherichia coli mutants deficient in thiamine and 4-thiouridine syntheses: Identification of a nuvC mutant. J. Bacteriol. 1982, 151, 899–904. [Google Scholar] [PubMed]
- Liu, Y.; Zhu, X.; Nakamura, A.; Orlando, R.; Söll, D.; Whitman, W.B. Biosynthesis of 4-thiouridine in tRNA in the methanogenic archaeon Methanococcus maripaludis. J. Biol. Chem. 2012, 287, 36683–36692. [Google Scholar] [CrossRef] [PubMed]





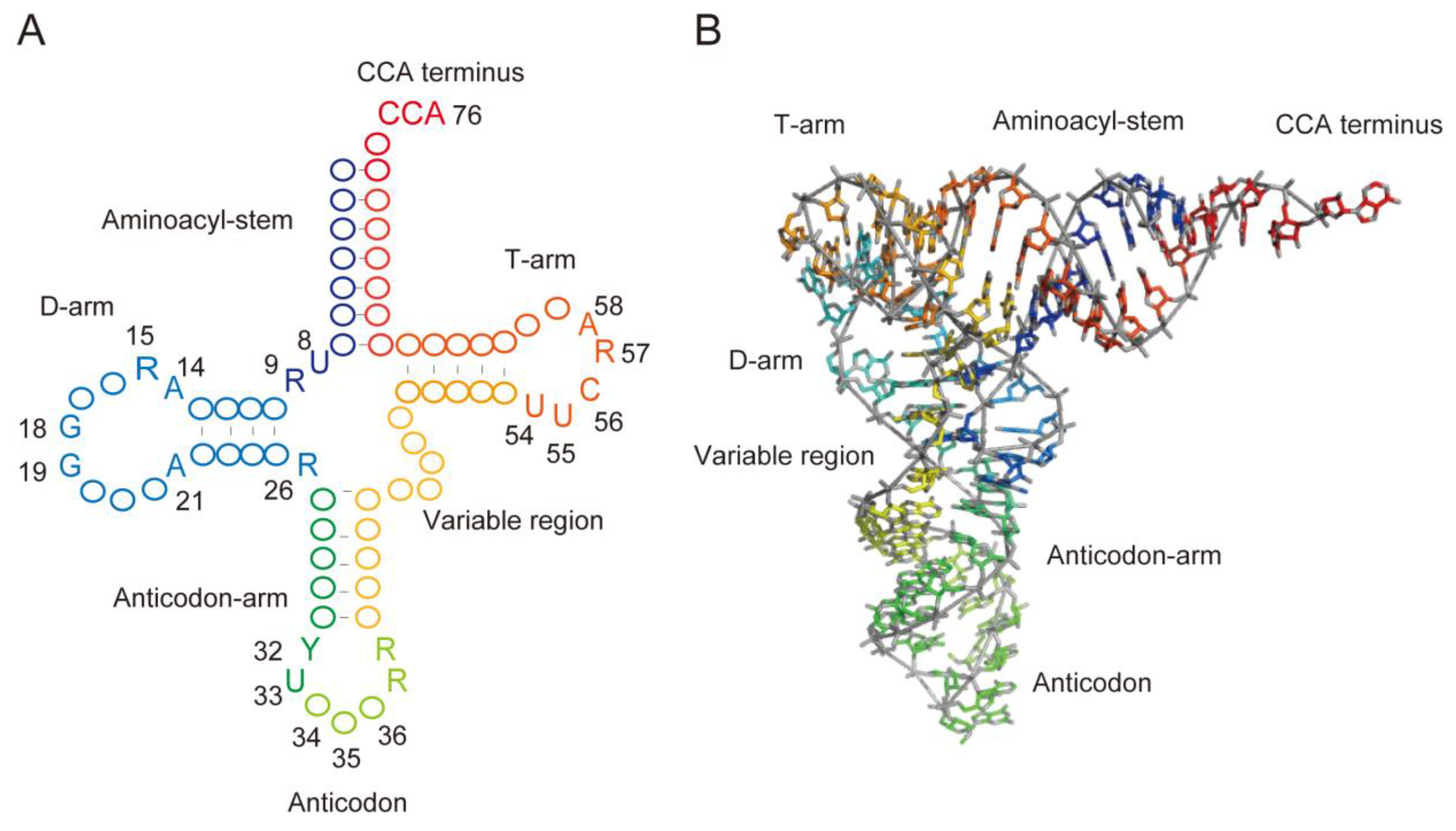{kind=link}
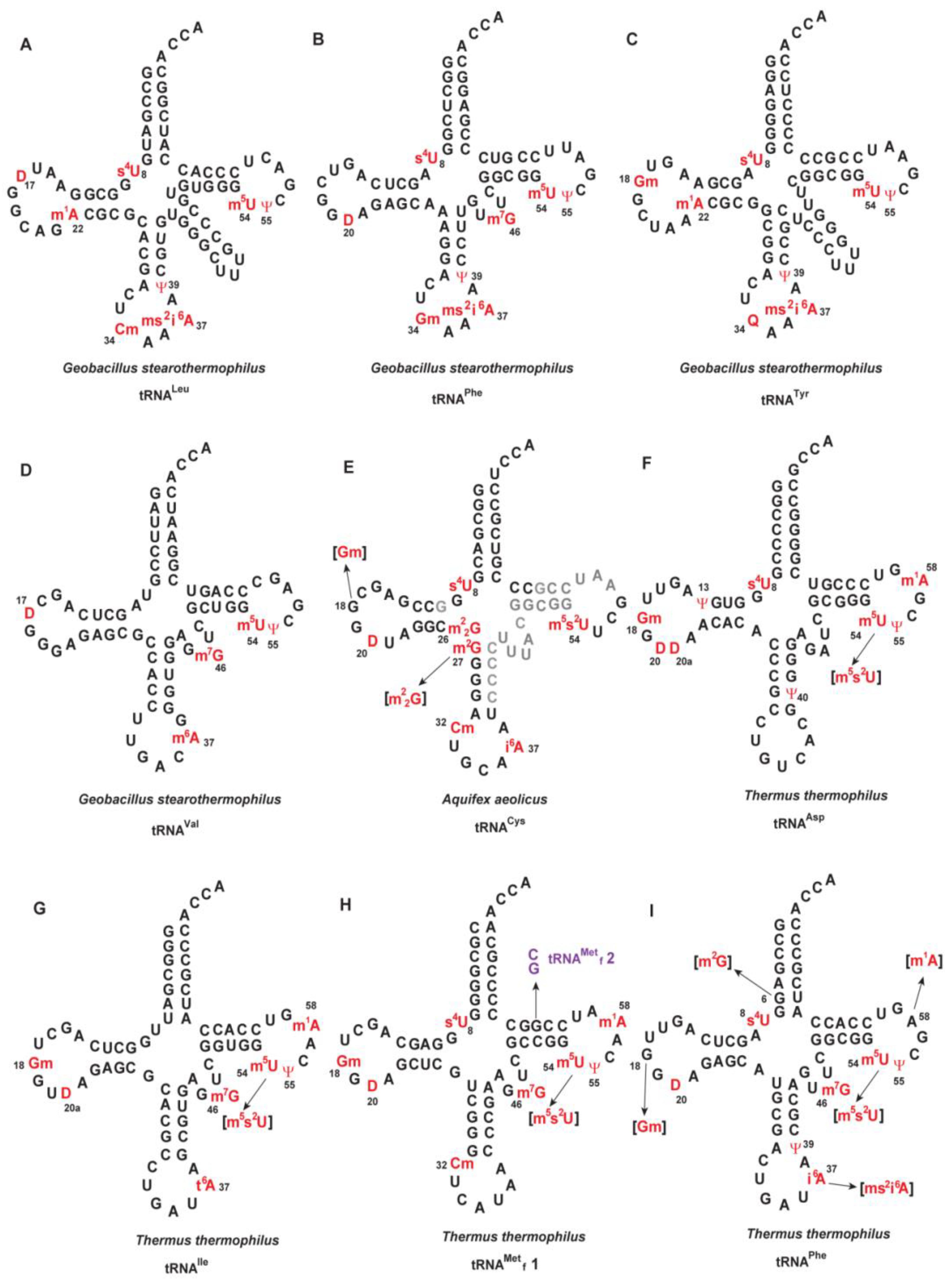{kind=link}
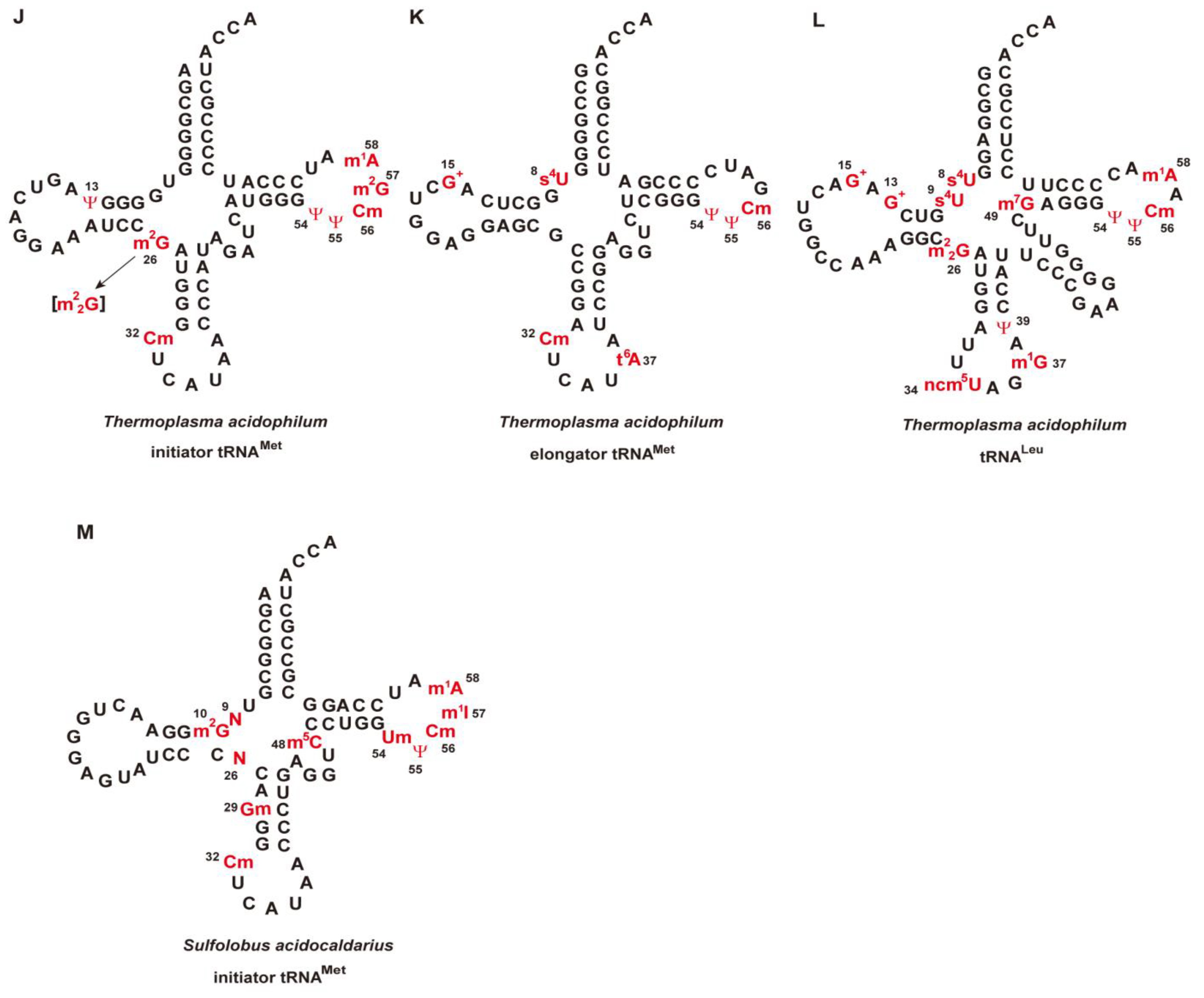{kind=link}
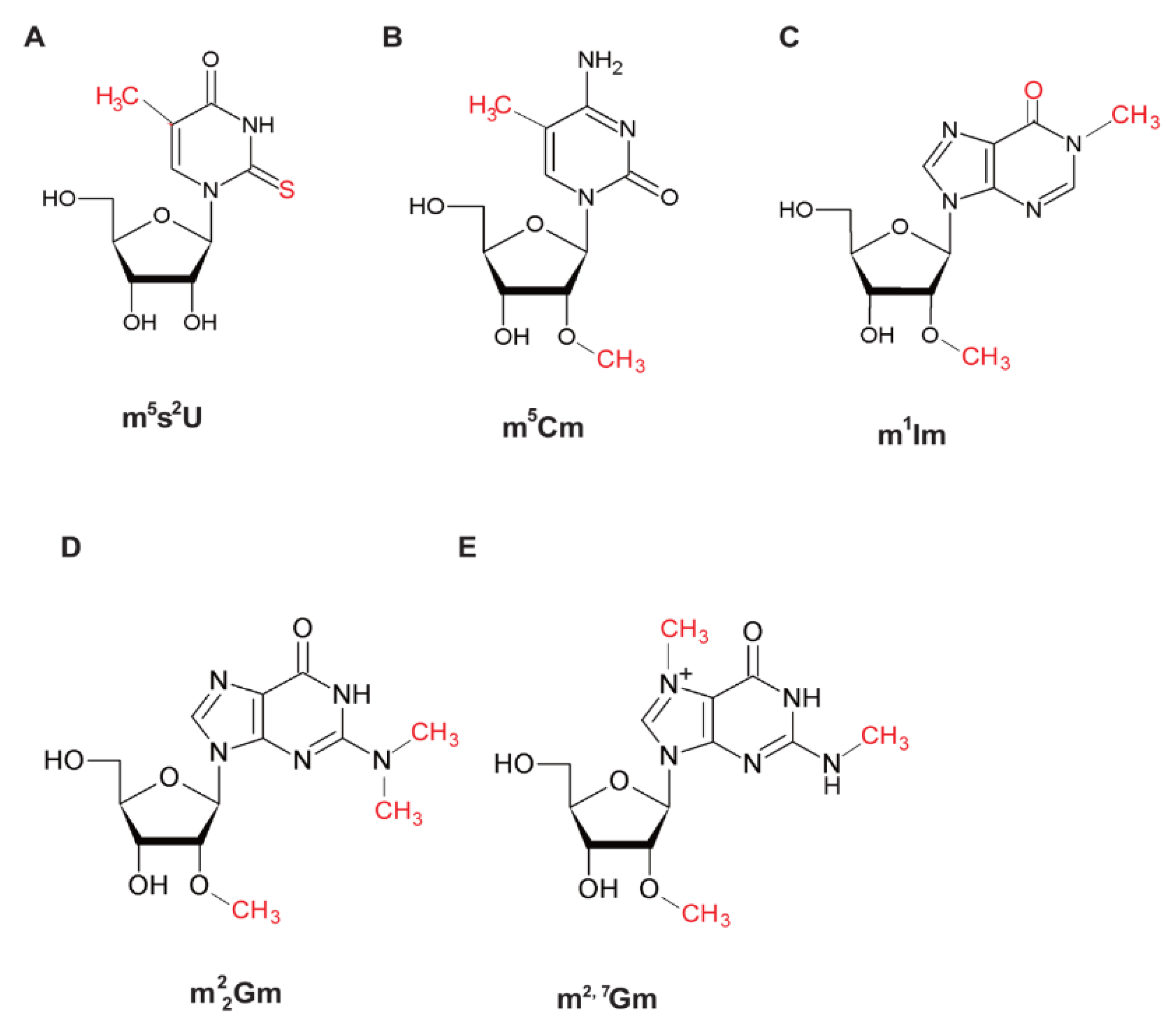{kind=link}
| Modified Nucleoside and Position | Distrib. | Modification Enzyme | Predicted Functions and Additional Information | References |
|---|---|---|---|---|
| Am6 | A | Unknown | Stabilization of aminoacyl-stem Enzymatic activity for Am6 formation has been detected in the cell extract of Pyrococcus furiosus | [9] |
| m2G6 | B/A | TrmN/Trm14 | Stabilization of aminoacyl-stem | [10,11,12,13,14,15] |
| U8 | A | CDAT8 | Increasing G-C content in tRNA genes In Methanopyrus kandleri, U8 in several tRNA is produced from C8 by the deamination [16] In Methanopyrus kandleri, numerous nucleosides in RNA may be 2-O-methylated (see main text) [17] | [16,17] |
| s4U8 | B/A | ThiI + IscS/ThiI | UV resistance in E. coli and Salmonella typhimurium (see main text) Stabilization of D-arm structure in E. coli (see main text) | [10,11,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35] |
| s4U8 and s4U9 | A | ThiI + α? | UV resistance Stabilization of D-arm structure (see main text) Sulfur-containing modifications in tRNA are reviewed in Reference [35]. | [36] |
| m1A9 | A | Archaeal Trm10 | Stabilization of the D-arm structure Prevention of formation of a Watson–Crick base pair Correct folding of the D-arm region | [37,38] |
| m1G9 and m1A9 | A | archaeal Trm10 | Stabilization of D-arm structure Prevention of formation of a Watson–Crick base pair Correct folding of D-arm region Thermococcus kodakarensis Trm10 forms m1G9 and m1A9, whereas Sulfolobus acidocaldarius Trm10 forms only m1A9 | [37,39] |
| (m2G10 and) m22G10 | A | archaeal Trm11 (Trm-G10; Trm-m22G10 enzyme) | Prevention of formation of a Watson-Crick base pair Correct folding of tRNA in Pyroccocus furiosus Correct folding of the D-arm region | [40,41,42,43] |
| Ψ13 | B/A | TruD/TruD or archaeal Pus7 | Stabilization of D-stem structure Archaeal Pus7 generally catalyzes formation of Ψ35 in tRNATyr, but Sulfolobus solfaraticus Pus7 has weak Ψ13 formation activity [46] | [23,44,45,46] |
| G+13 | A | ArcTGT + ArcS? | Stabilization of the D-arm structure Thermoplasma acidophilum tRNALeu exceptionally possesses a G+13 modification and T. acidophilum ArcTGT acts on positions 13 and 15 in this tRNA [47] | [36,47] |
| G+15 | A | ArcTGT + ArcS or QueF-like protein | Stabilization of interaction between the D-arm and the variable region Several archaea possess a split-type ArcTGT [60,61] Several species in Crenarchaeota possess a QueF-like protein instead of ArcS [60,62,63] G+ is not found in nucleosides from a Stetteria hydrogenophila tRNA mixture [56] | [25,36,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63] |
| D17 | B | Dus family protein? | Maintenance of D-loop flexibility D17 and D20 modifications have been reported in Geobacillus stearothermophilus tRNA. However, D17 and D20 are formed by DusB and DusA, respectively, in Escherichia coli [65,66] and the G. stearothermophilus genome possesses only one dus-like gene. This is also observed in Bacillus subtilis, which is a mesophilic eubacterium. | [18,19,64,65,66] |
| Gm18 | B | TrmH | Stabilization of the D-arm and the T-arm interaction. TrmH from thermophiles possess relative broad substrate tRNA specificities as compared with TrmH from E. coli. The substrate tRNA specificities of TrmH enzymes differ among thermophiles. TrmH from Thermus thermophilus can methylate all tRNA species. | [10,11,20,21,23,24,30,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81] |
| D20 | B | Dus family protein | Stabilization of local structure of D-loop in E. coli? In A. aeolicus, the nucleosides at positions 20 and 20a in tRNACys are D20 and U20a, respectively. Therefore, Dus from A. aeolicus may act only on U20 in tRNA. | [24,33,65,66,82] |
| D20 and D20a | B | DusA | Stabilization of local structure of the D-loop The melting temperature of a tRNA mixture from the E. coli dusA gene disruptant strain is lower than that from the wild-type strain [33]. Therefore, D20 and D20a modifications may contribute to stabilize local structure of the D-loop. Thermus thermophilus Dus was recently confirmed as a member of the DusA family [65,66,84,85]. | [21,22,23,33,65,66,67,83,84,85] |
| m1A22 | B | TrmK | Prevention of formation of a Watson–Crick base pair | [18,20,86] |
| Ψ22 | A | Unknown | The Ψ13-Ψ22 base pair may stabilize D-arm structure [88] | [87,88] |
| m2G26 and m22G26 | A | Trm1 | Stabilization of three-dimensional core structure Correct folding of tRNA Recently, it has been reported that m22G26 modification is required for correct folding of precursor tRNASer from Schizosaccharomyces pombe [94]. Therefore, a similar phenomenon may take place in thermophiles. | [9,25,44,89,90,91,92,93,94] |
| m2G26, m22G26, m2G27 and m22G27 | B | Trm1 | Stabilization of three-dimensional core structure in A. aeolicus. In the case of m2G27 and m22G27, stabilization of aminoacyl-stem | [24,95] |
| m22Gm26 | A | Trm1 + unknown MT | Stabilization of three-dimensional core structure The presence of m22Gm has been confirmed in nucleosides of a tRNA mixture from several thermophilic archaea [56,97,98,99,100]. Although the nucleoside at position 26 in S. acidocaldarius tRNAMeti was originally reported as an unidentified G modification [44], it was recently described as m22Gm [96]. The MT for 2’-O-methylation is unknown. | [44,96] |
| Cm32 | A | archaeal TrmJ | Stabilization of anticodon-loop | [96] |
| Cm32 and Nm32 | B | TrmJ | Stabilization of anticodon-loop TrmJ from E. coli does not recognize the base at position 32 [96,102]. Um32 and Am32 have not been reported in tRNAs from thermophilic eubacteria. | [96,101,102] |
| I34 | B | TadA | Alteration of codon–anticodon interaction A-to-I editing in tRNA is reviewed in Reference [107] | [103,104,105,106,107] |
| k2C34 | B | TilS | Alteration of codon–anticodon interaction (E. coli and B. subtilis) Change of recognition by aminoacyl-tRNA synthetase (E. coli and B. subtilis) Decoding of AUA codons by k2C34 and agm2C34 modifications is reviewed in References [114,115]. | [108,109,110,111,112,113] |
| agm2C34 | A | TiaS | Alteration of codon–anticodon interaction (Arhaeoglobus fulgidus and Haloarcula marismourtui) Change of recognition by aminoacyl-tRNA synthetase (Arhaeoglobus fulgidus and Haloarcula marismourtui) Decoding of AUA codons by k2C34 and agm2C34 modifications is reviewed in References [114,115]. | [114,115,116,117,118,119,120] |
| xm5U34 derivatives | B/A | MnmE + MnmG + MnmC (for mnm5U34 in eubacteria)/Elp3? + α (for cm5U34 in archaea) IscS + TusA + TusBCD + TusE + mnmA (for 2-thiolation in E. coli) or YrvO + mnmA (for 2-thiolation in B. subtilis) SAMP2 + UbaA + NcsA (for 2-thiolation in M. maripuludis) | Reinforcement of codon–anticodon interaction (E. coli and other mesophiles) Restriction of wobble base pairing (E. coli and other mesophiles) Prevention of frameshift errors (E. coli and other mesophiles) Biosynthesis pathways of xm5U34 derivatives are not completely clarified. Although the information on xm5U34 derivatives in tRNA from thermophiles is limited, the biosynthesis pathways may be common with those from mesophiles. For the functions and biosynthesis pathways for xm5U34 derivatives, see References [121,122,123,124,125,126,127,128,129,130,131,132,136,137,138,139,142]. For the thiolation of xm5s2U34 derivatives, see References [35,133,134,135]. Aquifex aeolicus exceptionally possesses a DUF752 protein, which is an MT for the xm5U34 modifications without an oxidase domain [136]. A mnm5U nucleoside has been found in modified nucleosides from unfractionated tRNA in several methane archaea [56]. Thermoplasma acidophilum tRNALeu possesses ncm5U34 [36]. Some thermophiles in Euryarchaea may have a cnm5U34 modification in tRNA [137]. The cm5U34 formation activity of Elp3 from Methanocaldococcus infernus has been reported [142]. Several related proteins for synthesis of xm5U34 derivatives from thermophiles have been used for structural studies [136,138,139,140,141]. | [34,35,36,56,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142] |
| Cm34 and cmnm5Um34 | B | TrmL | Reinforcement of codon–anticodon interaction (E. coli) | [18,143,144] |
| Gm34 | B | Unknown | Reinforcement of codon–anticodon interaction (G. stearothermophilus) | [19] |
| Q34 derivatives | B | Tgt + QueA + QueG | Reinforcement of codon–anticodon interaction (E. coli) Prevention of frame-shift error (E. coli) Biosynthesis pathways and functions of Q derivatives are reviewed in References [152,153]. A crystal structure of QueA from T. maritima has been reported [151]. | [20,122,145,146,147,148,149,150,151,152,153] |
| Cm34 and Um39 (or Cm39) | A | L7Ae + Nop5 + archaeal fibrillarin + Box C/D guide RNA (intron) | Reinforcement of codon–anticodon interaction Reinforcement of anticodon-arm In several archaea, an intron in precursor tRNATrp functions as a Box C/D guide RNA. | [9,154,155] |
| Ψ35 | A | aPus7 and H/ACA guide RNA system | Reinforcement of codon–anticodon interaction | [46] |
| m1G37 | B/A | TrmD/Trm5 | Prevention of frame-shift error (E. coli and other mesophiles) Recognition by aminoacyl-tRNA synthetase (Saccharomyces cerevisiae) | [36,156,157,158,159,160,161,162,163,164,165,166,167,168,169,170,171] |
| wyosine37 derivatives | A | Trm5 + Taw1 + Taw2 + Taw3 | Reinforcement of codon–anticodon interaction Prevention of the frame-shift error In several archaea, m1G37 in tRNAPhe is further modified to wyosine derivatives. For the biogenesis pathway of wyosine derivatives, see References [181,182,183]. | [172,173,174,175,176,177,178,179,180,181,182,183] |
| t6A37 derivatives | B/A | TsaB, TsaC (TsaC2), TsaD and TsaE/KEOPS complex: Kae1, Bud32, Cgi121 and Pcc1 + Sua5 | Reinforcement of codon–anticodon interaction Prevention of frame-shift error Recognition by aminoacyl-tRNA synthetases The biogenesis pathway for t6A derivatives is reviewed in Reference [191] | [68,184,185,186,187,188,189,190,191] |
| i6A37 derivatives | B | MiaA + MiaB | Prevention of frame-shift error Reinforcement of codon–anticodon interaction Recognition by aminoacyl-tRNA synthetases i6A derivatives are reviewed in Reference [197] | [10,11,18,19,20,24,192,193,194,195,196,197] |
| m6A37 | B | YfiC (TrmG?) | [64,198] | |
| Ψ38, Ψ39 and Ψ40 | B/A | TruA/Pus3 | Prevention of frame-shift error (E. coli) Reinforcement of anticodon-arm | [10,11,18,19,20,23,87,199,200,201,202,203] |
| m7G46 | B | TrmB | Stabilization of three-dimensional core In the case of T. thermophilus, m7G46 modification functions a key factor in a network between modified nucleosides in tRNA and tRNA modification enzymes (see main text) [11] | [10,11,19,67,204,205,206,207,208] |
| m5C48 and m5C49 | A | archaeal Trm4 | Stabilization of three-dimensional core | [9,209,210] |
| m7G49 | A | Unknown | [36] | |
| m5C51 | A | Unknown | Stabilization of T-arm structure | [209] |
| m5C52 | A | Unknown | Stabilization of T-arm structure | [209] |
| Ψ54 and Ψ55 | A | Pus10 | Stabilization of D-arm and T-arm interaction | [211,212,213,214] |
| m1Ψ54 | A | Pus10 + TrmY | Stabilization of D-arm and T-arm interaction | [215,216,217] |
| m5U54 + m5s2U54 | B/A | TrmFO + TtuA + TtuB + TtuC + TtuD + IscS/TrmA + TtuA? + TtuB? + α | Stabilization of D-arm and T-arm interaction (see main text) 2-Thiolation of m5s2U54 in tRNA is reviewed in Reference [239] | [10,11,21,22,23,24,67,97,98,134,218,219,220,221,222,223,224,225,226,227,228,229,230,231,232,233,234,235,236,237,238,239] |
| Um54 | A | Unknown | Stabilization of D-arm and T-arm interaction | [44] |
| Ψ55 | B/A | TruB/Pus10 or archaeal Cbf5 + α | Stabilization of D-arm and T-arm interaction In the case of T. thermophilus, Ψ55 is required for low-temperature adaptation (see main text) [248]. | [10,11,18,19,20,23,25,36,44,64,67,211,212,213,214,240,241,242,243,244,245,246,247,248] |
| Cm56 | A | Trm56 | Stabilization of D-arm and T-arm interaction | [9,25,36,44,48,89,249,250,251] |
| m2G57 | A | Unknown | [44,252] | |
| m1I57 | A | archaeal TrmI + unknown deaminase | Stabilization of T-arm structure | [44,253,254] |
| m1A57 and m1A58 | A | archaeal TrmI | Stabilization of T-arm structure | [44,255,256,257,258] |
| m1A58 | B | TrmI | Stabilization of T-arm structure | [11,23,67,204,259,260,261,262,263,264] |
| Species | Predicted Enzyme | Distinct tRNA Modifications and General Information | References |
|---|---|---|---|
| Moderate Thermophiles | |||
| Eubacteria | |||
| Geobacillus stearothermophilus (Bacillus stearothermophilus) 30–75 °C | Sequences of tRNALeu [18], tRNAPhe [19], tRNATyr [20], and tRNAVal2 [64] have been reported (Figure 2). The majority of modifications in tRNA are similar to those in B. subtilis. With increasing culture temperature, the extent of 2’-O-methylation in the tRNA mixture increases [309]. | ||
| Gm18 (TrmH?) | Although trmH is not encoded in the B. subtilis genome, a trmH-like gene is encoded in the G. stearothermophilus genome. Gm18 has been found in tRNATyr but not in tRNALeu. This modification pattern suggests that the substrate tRNA specificity of G. stearothermophilus TrmH may be different from that of other known TrmH enzymes. | [20] | |
| D17, D20 and D20a (Dus family protein?) | In G. stearothermophilus tRNA, D17, D20, and D20a modifications have been reported. In E. coli, three Dus family proteins known as DusA, DusB, and DusC, produce D20 and D20a, D17, and D16, respectively [65,66]. In the G. stearothermophilus genome, however, only one gene is annotated as a dus-like gene. Therefore, D modifications in G. stearothermophilus cannot be explained by the tRNA substrate specificity of the known Dus proteins. | [17,19,64] | |
| m1A22 (TrmK?) | The m1A22 modification has been found in tRNATyr and tRNASer from B. subtilis and Mycoplasma capricolum [310,311]. G. stearothermophilus tRNALeu and tRNATyr possess m1A22 [18,20]. The presence of a trmK-like gene in the genome of G. stearothermophilus has been reported [86]. | [20,86] | |
| Gm34 (unknown MT) | G. stearothermophilus tRNAPhe possesses Gm34 (Figure 2B) [19]. In contrast, the nucleoside at position 34 in E. coli tRNAPhe is unmodified G. Given that E. coli TrmL acts only on tRNALeu isoacceptors [143], the 2′-O-methylation of G34 in tRNAPhe from G. stearothermophilus is cannot be simply explained by the activity of known TrmL. | [19] | |
| m6A37 (YfiC; TrmG?) | [198] | ||
| Archaea | |||
| Methanobacterium thermoaggregans Optimum growth temperature 60 °C | Sequences of tRNAAsn and tRNAGly have been reported [8]. | ||
| Methanobacterium thermoautotrophicum 45–75 °C | The modified nucleosides in unfractionated tRNA are essentially common to those in tRNA from mesophilic methane archaea [97]. | ||
| Methanococcus thermolithotrophicus 17–62 °C | The modified nucleosides in unfractionated tRNA are essentially common to those in tRNA from mesophilic methane archaea [99]. | ||
| Thermoplasma acidophilum Optimum growth temperature 55–60 °C | Sequences of tRNAMeti [44,252], tRNAMetm [25], and tRNALeu [36] have been reported. Several recombinant tRNA modification enzymes have been used for biochemical studies. | ||
| s4U8 and s4U9 (ThiI? + α) | The s4U9 modification has been found in tRNALeu [36]. The sulfur donor for s4U formation is unknown [35]. | [36] | |
| G+13 and G+15 (ArcTGT + ArcS?) | The G+13 modification has been found only in tRNALeu from T. acidophilum [36]. ArcTGT from T. acidophilum acts on both G13 and G15 in tRNALeu [47]. | [36,47] | |
| m22G26 (Trm1) | [89] | ||
| ncm5U34 (Elp3?) | [36] | ||
| m1G37 (Trm5) | [89] | ||
| m7G49 (unknown MT) | [36] | ||
| Cm56 (Trm56) | The presence of unusual trm56-like gene in the T. acidophilum genome has been reported in a bioinformatics study [250]. The Trm56 enzymatic activity has been confirmed via the recombinant protein [89]. T. acidophilum Trm56 exceptionally possesses a long C-terminal region in the SPOUT tRNA MT [312]. | [89,250,312] | |
| Extreme-thermophiles and Hyper-thermophiles | |||
| Eubacteria | |||
| Aquifex aeolicus Optimum growth temperature 85–94 °C | The partial sequence of tRNACys has been reported [24] (Figure 2E). Several tRNA MT activities have been detected in the A. aeolicus cell extract using an E. coli tRNA mixture [24]. The tRNA modification enzymes listed below were characterized via recombinant proteins. | ||
| Gm18 (TrmH) | [74,77] | ||
| D20 (Dus) | D20 exists in tRNACys. However, the nucleoside at position 20a is unmodified U [24]. Therefore, A. aeolicus Dus may act only on U20. | [24,82] | |
| m2G26, m22G26, m2G27 and m22G27 (Trm1) | Aquifex aeolicus exceptionally possesses Trm1 in eubacteria [24]. | [24,95] | |
| I34 (TadA) | [104,105] | ||
| mnm5U34 (MnmC2) | MnmC catalyzes the final methylation step of mnm5U synthesis. Aquifex aeolicus MnmC2 comprises only an MT domain. | [136] | |
| (MnmD; previously called GidA) | [140,141] | ||
| k2C34 (TilS) | [111,112,113] | ||
| m1G37 (TrmD) | The dimer structure of A. aeolicus TrmD is stabilized by inter-subunit disulfide bonds [165]. | [160,162,165] | |
| m7G46 (TrmB) | TrmB proteins from thermophiles (A. aeolicus and T. thermophilus) possess a long C-terminal region. | [206,207,208] | |
| m5U54 and m5s2U54 (TrmFO) | The presence of trmFO gene in A. aeolicus genome was initially described in Reference [221]. | [24,221] | |
| m1A58 (TrmI) | [257,262] | ||
| Thermotoga maritima 80–90 °C | Sequences of tRNA from T. maritima have not been reported. Recombinant proteins have been used for biochemical and structural studies. | ||
| hn6A (?) | hn6A was first identified in modified nucleosides from unfractionated tRNA from T. maritima [313]. Because hn6A was subsequently found in modified nucleosides from psychrophilic archaea [56], it is not a thermophile-specific modification. Thermotoga maritima and Thermodesulfobacterium commune exceptionally possess hn6A in eubacteria. The modification position in tRNA, modified tRNA species, and biosynthesis pathway of hn6A are unknown. | [56,313] | |
| s4U8 (ThiI + IscS) | [31,32] | ||
| oQ34 (QueA) | [151] | ||
| mnm5U34 (TrmE) | [138,139] | ||
| t6A37 (TsaB, TsaC/TsaC2, TsaD and TsaE) | [190] | ||
| ms2i6A37 (MiaB) | [194,195,196] | ||
| m1G37 (TrmD) | [171] | ||
| m5U54 and m5s2U54 (TrmFO and TtuA) | The m5s2U nucleoside has been found in unfractionated tRNA from T. maritima [97]. | [97,134,221,222] | |
| Ψ55 (TruD) | [244,245,246,247] | ||
| m1A58 (TrmI) | [257] | ||
| Thermodesulfobacterium commune Optimum growth temperature 70 °C | hn6A and ms2hn6A (?) | hn6A and ms2hn6A have been found in modified nucleosides from unfractionated tRNA from T. commune. The ms2hn6A modification may be derived from hn6A. So far, T. commune is the only eubacterium that possesses ms2hn6A in tRNA. The modification position in tRNA, modified tRNA species, and biogenesis pathway of hn6A and ms2hn6A are unknown. | [313] |
| Thermus flavus Optimum growth temperature 70 °C | Partial purification of tRNA m1A58 MT has been reported: the activity of tRNA m7G46 MT has also been described [204]. | ||
| Thermus thermophiles 50–83 °C | Sequences of tRNAMetf1 [21], tRNAMetf2 [21], tRNAIle1 [67], tRNAAsp [23], and tRNAPhe [10,11] have been reported (Figure 2). Partial sequences of tRNASerGGA [259], tRNAProGGG [314], and tRNAProGGA [314] have been determined. The modification extent of Gm18, m5s2U54 and m1A58 changes with the culture temperature. At high temperatures (>75 °C), m7G46 [11], m5s2U54 [230], and m1A58 [260] modifications are essential for survival. At low temperatures (<55 °C), Ψ55 is essential for survival [248] and m5U54 supports this effect [225] (see the main text). Recombinant proteins have been used for biochemical and structural studies. | ||
| m2G6 (TrmN) | [10,11,13,14,15] | ||
| Gm18 (TrmH) | [10,11,21,23,30,69,70,71,72,73,75,76,78,79,80,81] | ||
| D20 and D20a (DusA) | [10,11,23,67,83,84,85] | ||
| Cm34 and cmnm5Um34 (TrmL) | [144] | ||
| Ψ39 and Ψ40 (TruA) | [10,11,18,23,202] | ||
| m7G46 (TrmB) | [10,11,21,23] | ||
| m5U54 and m5s2U54 (TrmFO + TtuA + TtuB + TtuC + TtuD + IscS) | [10,11,17,21,23,67,218,219,220,221,222,223,224,225,226,227,228,229,230,231,232,233,234,235,236,239] | ||
| Ψ55 (TruB) | [10,11,21,23,67,248] | ||
| m1A58 (TrmI) | [11,30,257,259,260,261,263,264] | ||
| Archaea | |||
| Aerophyrum pernix 80–100 °C | Ψ13 and Ψ15 (archaeal Pus7 and H/ACA guide RNA system) | A guide RNA for Ψ formation has been predicted based on genome sequencing. | [46] |
| Archaeoglobus fulgidus 60–95 °C | Modified nucleosides in unfractionated tRNA from A. fulgidus have been reported [97]. | ||
| agm2C34 (TiaS) | [116,118,119,315] | ||
| Methanocaldcoccus igneus (Methanococcus igneus; Methanotorris igneus) 45–91 °C | Modified nucleosides in unfractionated tRNA from M. igneus have been reported [56,99]. | ||
| Methanocaldococcus infernus 55–92 °C | cm5U34 (Elp3) | [142] | |
| Methanocaldcoccus jannashii (Methanococcus janaschii) 48–94 °C | Although sequences of tRNA are unknown, the recombinant proteins listed below have been used for biochemical and structural studies. | ||
| m2G6 (Trm14) | [12] | ||
| G+15 (ArcTGT + ArcS) | [51,59] | ||
| Cm34 and Um39 (L7Ae, Nop5, aFib, Box C/D guide RNA system) | [316] | ||
| m1G37 (Trm5) | [159,161,163,164,166,167,168,169,170] | ||
| imG237 (Trm5b + Taw1) | [173,179] | ||
| yW-8637 (Taw2) | [174] | ||
| m5C48 and m5C49 (archaeal Trm4) | [210] | ||
| Ψ54 and Ψ55 (Pus10) | [211,212,213,214] | ||
| m1Ψ54 (Pus10 + TrmY) | [215,216,217] | ||
| Ψ55 (archaeal Cbf5) | [240] | ||
| Methanopyrus kandleri 84–110 °C (Strain 116: up to 122 °C) | Many unique modified nucleosides have been found in unfractionated tRNA [100]. tRNAs likely contain many 2’-O-methylated nucleosides derived from the C/D box guide RNA system [17]. | ||
| ac6A (?) | The ac6A nucleoside has been purified from the modified nucleosides in unfractionated tRNA and its structure determined. The modification site, modified tRNA species, and biosynthesis pathway are unknown. | [100] | |
| U8 (CDAT8) | [16] | ||
| Methanothermus fervidus 80–97 °C | Only tRNA genes were reported in an early study [317]. | ||
| Nanoarchaeum equitans 70–98 °C | A unique tRNA processing system has been found [318,319]. The processing of small RNAs in N. equitans is reviewed in Reference [320]. | ||
| m1G37 and imG237 (Trm5a) | [176] | ||
| m5U54 (TrmA-like protein) | [237] | ||
| Pyrobaculum aerophilum Optimum growth temperature 100 °C | Cm56 (L7Ae, Nop5, aFib, Box C/D guide RNA system) | Cm56 in tRNA is generally produced by Trm56. However, this modification in P. aerophilum is synthesized by the C/D box guide RNA system. | [249] |
| Pyrobaculum calidifontis 90–95 °C | G+15 (ArcTGT + QueF-like protein) | Eubacterial QueF catalyzes the conversion from preQ0 to preQ1. In P. caldifontis, however, QueF-like protein catalyzes the conversion from preQ0 at position 15 in tRNA to G+15. | [60,62,63] |
| Pyrobaculum islandicum Optimum growth temperature 100 °C | Modified nucleosides in unfractionated tRNA from P. islandicum have been reported [97]. | ||
| Pyrococcus abyssi Optimum growth temperature 96 °C | No tRNA sequence has been determined. However, the tRNA modification enzymes listed below have been characterized. | ||
| m2G10 and m22G10 (archaeal Trm11, Trm-G10 enzyme, Trm-m22G10 enzyme) | [40,41] | ||
| Ψ13 and Ψ35 (archaeal Pus7 and H/ACA guide RNA system) | [46] | ||
| Cm34 and Um39 (L7Ae, Nop5, aFib, and C/D box guide RNA system) | Cm34 and Um39 in tRNATrp are formed by the C/D box guide RNA system in which the intron functions as a guide RNA. | [154,155] | |
| m1G37 (Trm5b) | [180] | ||
| m1G37 and imG237 (Trm5a) | [176,177,179] | ||
| imG-1437 (Taw1) | [173,175] | ||
| t6A37 (Kae1) | [185] | ||
| (KEOPS complex) | [184] | ||
| (Sua5 + KEOPS complex) | [187,189] | ||
| m5C48 and m5C49 (archaeal Trm4 + archaese) | [209] | ||
| m5U54 (TrmA-like protein, PAB0719) | [237,238] | ||
| Ψ55 (Cbf5 + Nop10) | [241] | ||
| Cm56 (Trm56) | [249] | ||
| m1A57 and m1A58 (archaeal TrmI) | [255,256,257,258] | ||
| Pyrococcus furiosus Optimum growth temperature 100 °C | Modified nucleosides in unfractionated tRNA from P. furiosus have been reported [98]. Activity of several tRNA modification enzymes has been detected in the cell extract of P. furious [9]. | ||
| m2G6 (Trm14) | [13,15] | ||
| m2G10 and m22G10 (archaeal Trm11, Trm-G10 enzyme, Trm-m22G10 enzyme) | [42] | ||
| G+15 (ArcTGT) | [57] | ||
| m2G26 and m22G26 (Trm1) | [91,92] | ||
| t6A37 (KEOPS complex) | [188] | ||
| Ψ54 and Ψ55 (Pus10) | [212,214] | ||
| Ψ55 (Cbf5 + Nop10 + Gar1) | [242] | ||
| Pyrococcus horikoshii 80–102 °C | The crystal structure of Nop5 in the C/D box guide RNA system from P. horikoshii has been solved [321]. | ||
| G+15 (ArcTGT) | [50,52,53,54,55,89] | ||
| m2G26 and m22G26 (Trm1) | [89,93] | ||
| yW-8637 (Taw2) | [174] | ||
| m5s2U54 (TtuA) | [233] | ||
| Cm56 (Trm56) | [251] | ||
| Pyrodictium occultum Optimum growth temperature 105 °C | Modified nucleosides in unfractionated tRNA have been analyzed and many 2’-O-methylated nucleosides found [97,98]. mimG was originally found among the modified nucleosides in tRNAs from P. occultum, Sulfolobus solfaraticus, and Thermoproteus neutrophilus [322]. Although the melting temperature of P. occultum tRNAMeti transcript is only 80 °C and that of native tRNAMeti is more than 100 °C (see main text) [323]. | ||
| Pyrolobus fumarii This archaeon can survive at 113 °C. | Modified nucleosides in unfractionated tRNA have been analyzed [324]. | ||
| Stetteria hydrogenophila Optimum growth temperature 95 °C | Modified nucleosides in unfractionated tRNA have been analyzed and methyl-hn6A, ms2hn6A, and m2, 7Gm identified [56]. | ||
| Sulfolobus acidocaldarius Optimum growth temperature 75–80 °C | Sequence of tRNAMeti has been reported [44]. The m1I57 modification was originally found in tRNAs from S. acidocaldarius and Haloferax volcanii [253]. G+ was first isolated from the nucleosides in S. acidocaldarius tRNAs and its structure determined [49]. The structures of wyosine derivatives (imG-14 and imG2) have been determined by using the nucleosides from S. acidocaldarius tRNAs [325]. | ||
| m1A9 (archaeal Trm10) | [37,38] | ||
| Ψ13 and Ψ35 (archaeal Pus7 and H/ACA guide RNA system) | [46] | ||
| Cm32 (archaeal TrmJ) | [96] | ||
| Sulfolobus solfaraticus 55–90 °C | mimG was originally found among the modified nucleosides in tRNAs from P. occultum, S. solfaraticus, and Thermoproteus neutrophilus [322]. The structure of box C/D RNP from S. solfaraticus has been reported [326]. | ||
| agm2C (TiaS) | The identification of agm2C34 in Haloarcula marismortui tRNAIle2 and the presence of agm2C in S. solfaraticus tRNA have been reported. | [117] | |
| Ψ13 and Ψ35 (archaeal Pus7 and H/ACA guide RNA system) | Generally, Ψ35 in tRNATyr is synthesized by archaeal Pus7. However, Pus7 from S. solfaraticus possesses weak Ψ13 formation activity but not Ψ35 formation activity. In S. solfaraticus and A. pernix, a guide RNA for Ψ35 formation exists. | [46] | |
| imG237 (Trm5a; SSO2439 protein) | Trm5a (SSO2439 protein) does not possess m1G37 formation activity and is used only for imG2 formation. | [178] | |
| mimG37 (Taw3) | [180] | ||
| Sulfolobus tokodaii This archaeon can survive at 87 °C. | Ψ13 and Ψ35 (archaeal Pus7 and H/ACA guide RNA system) | [46] | |
| t6A37 (Sua5) | [327,328,329] | ||
| Thermococuus celer This archaeon can survive at 85 °C. | Although tRNA genes were analyzed in an early study [330], there is no information on tRNA modifications. | ||
| Thermococcus kodakarensis (Thermococcus kodakaraensis; Pyrococcus kodakarensis) 65–100 °C | m1A9 and m1G9 (archaeal Trm10) | [37,39] | |
| m2G10 and m22G10 (archaeal Trm11, Trm-G10 enzyme, Trm-m22G10 enzyme) | [43] | ||
| G+15 (ArcTGT) | [47] | ||
| m5U54 (TrmA-like protein) | [237] | ||
| Thermoproteus neutrophilus Optimum growth temperature 85 °C | Modified nucleosides in unfractionated tRNA have been analyzed [97]. mimG was originally found among the modified nucleosides in tRNAs from P. occultum, S. solfaraticus, and T. neutrophilus [322]. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hori, H.; Kawamura, T.; Awai, T.; Ochi, A.; Yamagami, R.; Tomikawa, C.; Hirata, A. Transfer RNA Modification Enzymes from Thermophiles and Their Modified Nucleosides in tRNA. Microorganisms 2018, 6, 110. https://doi.org/10.3390/microorganisms6040110
Hori H, Kawamura T, Awai T, Ochi A, Yamagami R, Tomikawa C, Hirata A. Transfer RNA Modification Enzymes from Thermophiles and Their Modified Nucleosides in tRNA. Microorganisms. 2018; 6(4):110. https://doi.org/10.3390/microorganisms6040110
Chicago/Turabian StyleHori, Hiroyuki, Takuya Kawamura, Takako Awai, Anna Ochi, Ryota Yamagami, Chie Tomikawa, and Akira Hirata. 2018. "Transfer RNA Modification Enzymes from Thermophiles and Their Modified Nucleosides in tRNA" Microorganisms 6, no. 4: 110. https://doi.org/10.3390/microorganisms6040110
APA StyleHori, H., Kawamura, T., Awai, T., Ochi, A., Yamagami, R., Tomikawa, C., & Hirata, A. (2018). Transfer RNA Modification Enzymes from Thermophiles and Their Modified Nucleosides in tRNA. Microorganisms, 6(4), 110. https://doi.org/10.3390/microorganisms6040110