Expression of DinJ-YafQ System of Lactobacillus casei Group Strains in Response to Food Processing Stresses

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Sequence Analysis
2.3. Culture Conditions for Gene Expression Studies
2.4. RNA Extraction and cDNA Synthesis
2.5. Reference Gene Selection
2.6. Relative Quantification of DinJ-YafQ Expression
2.7. Statistical Analysis
3. Results
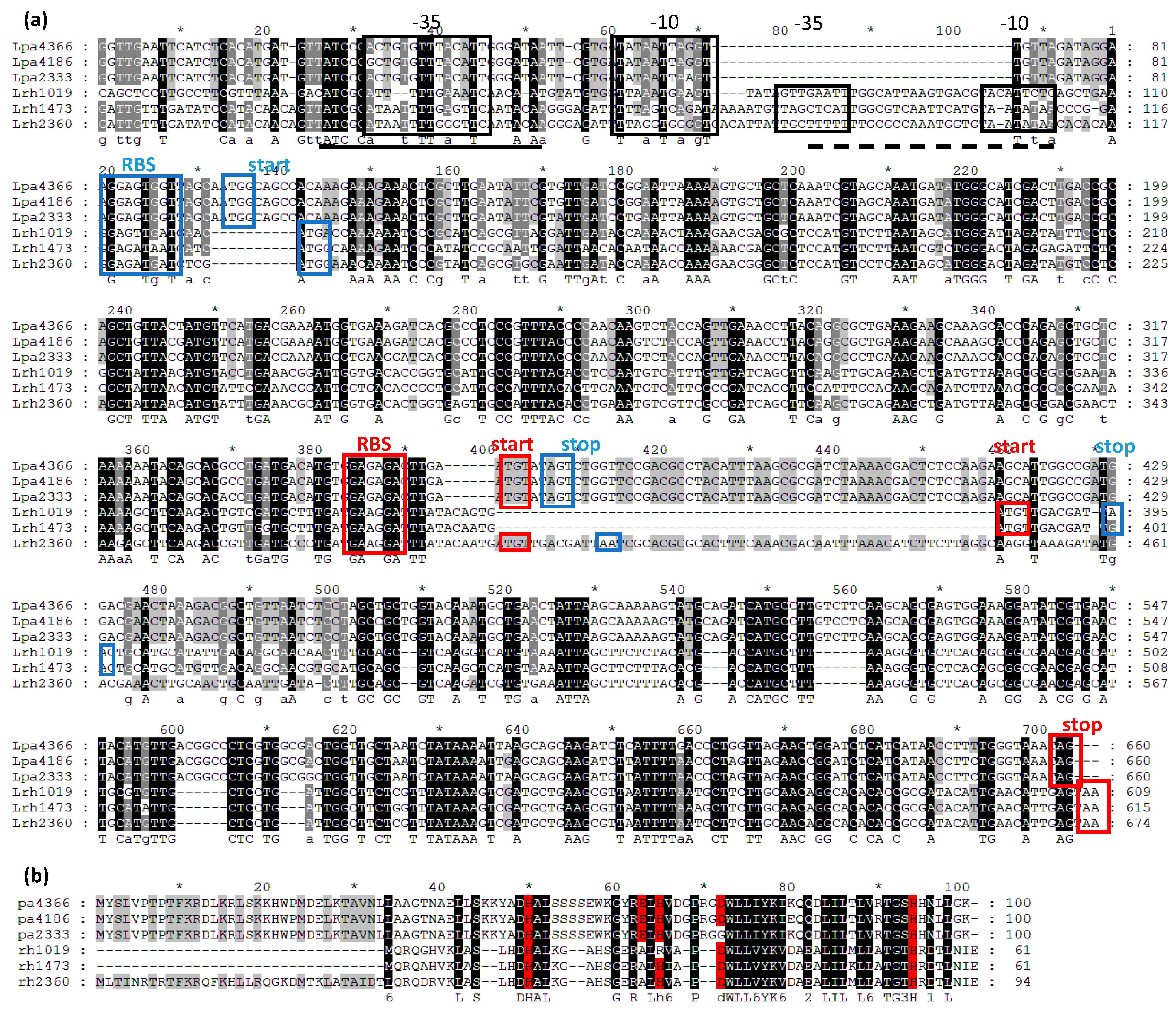
3.1. Sequence Analysis
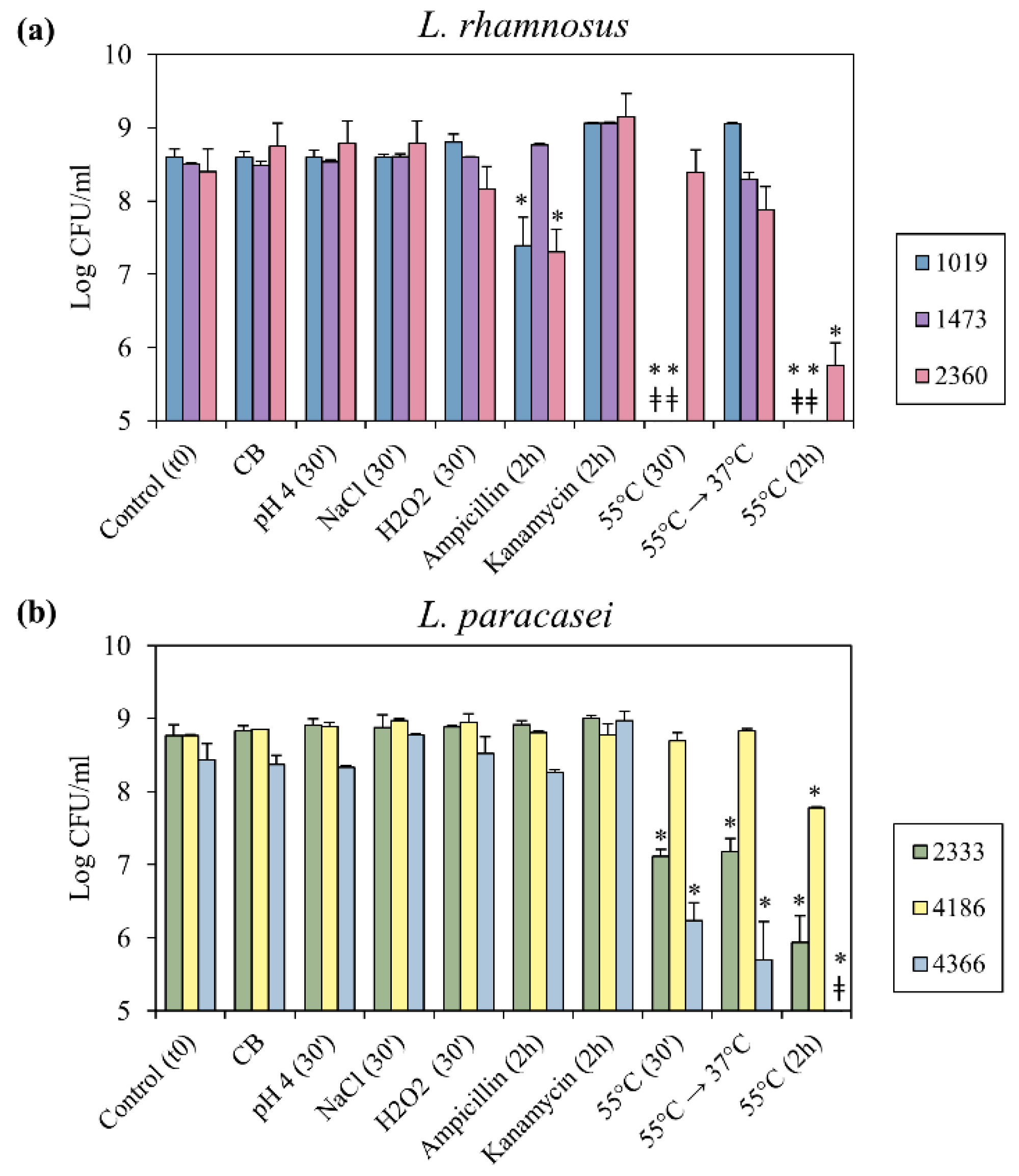
3.2. Bacterial Culturability in Response to Food Processing Stresses and to the Exposure to Antibiotics
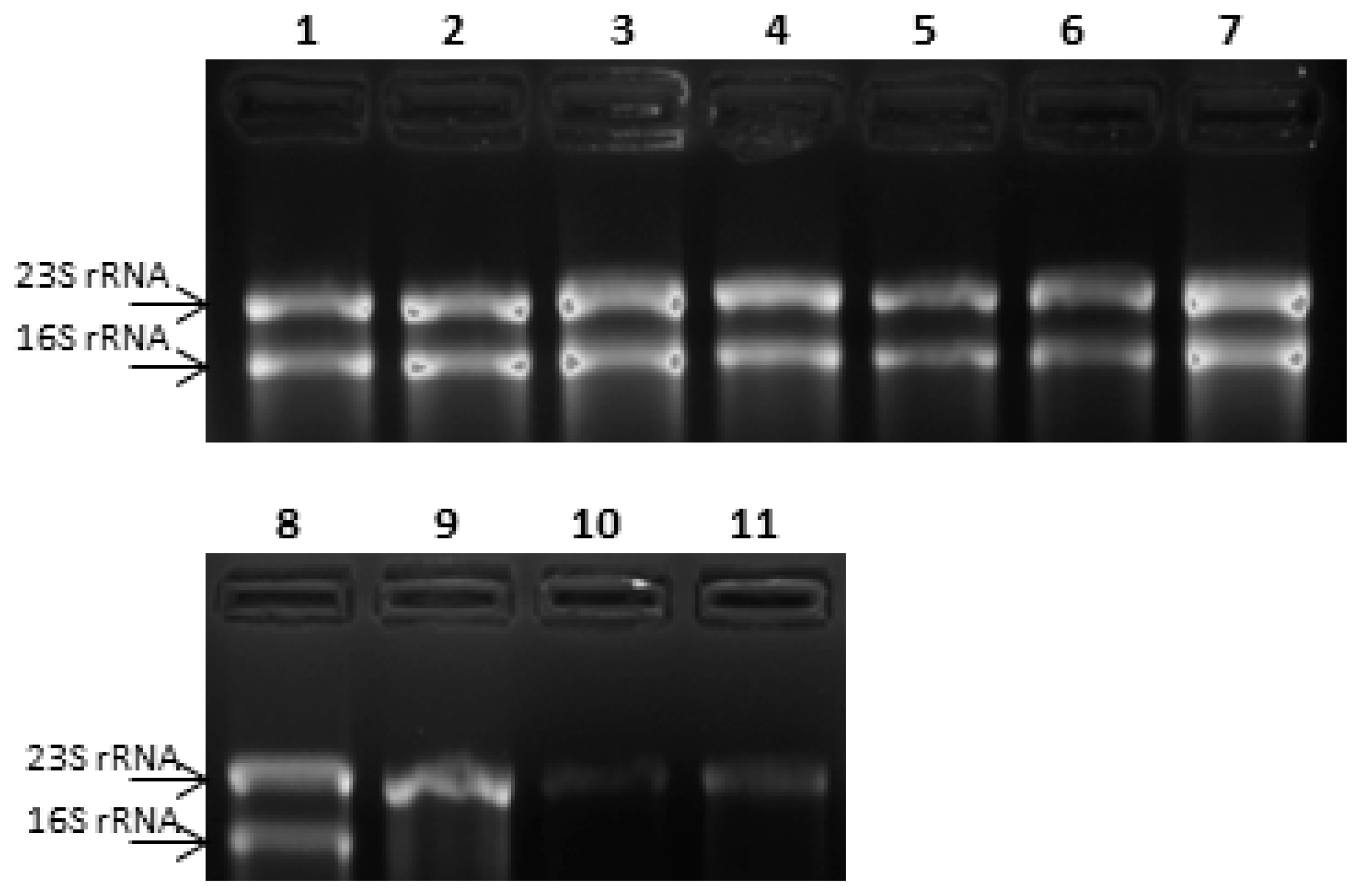
3.3. Selection of a Reference Gene for Relative Expression Studies
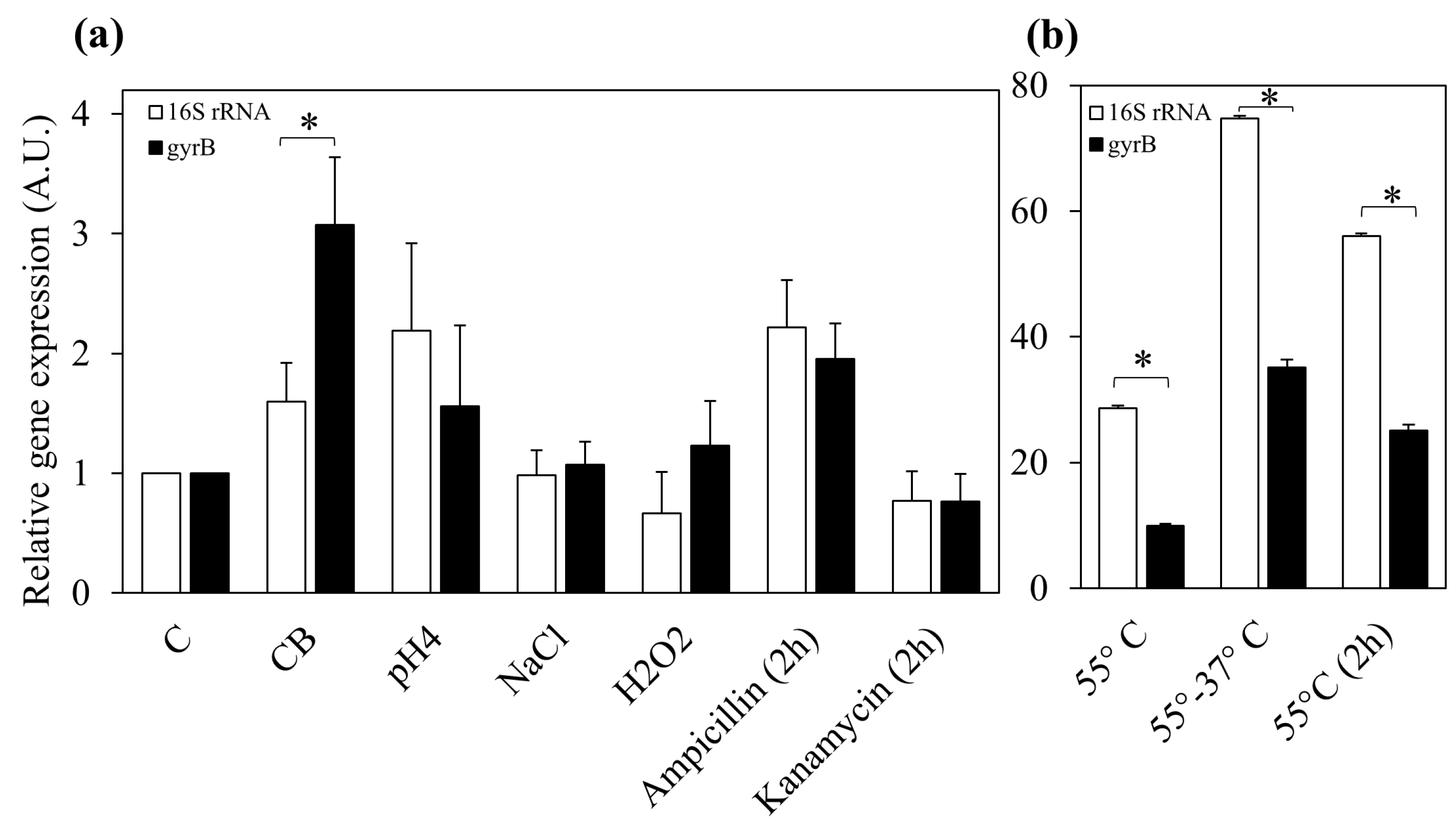
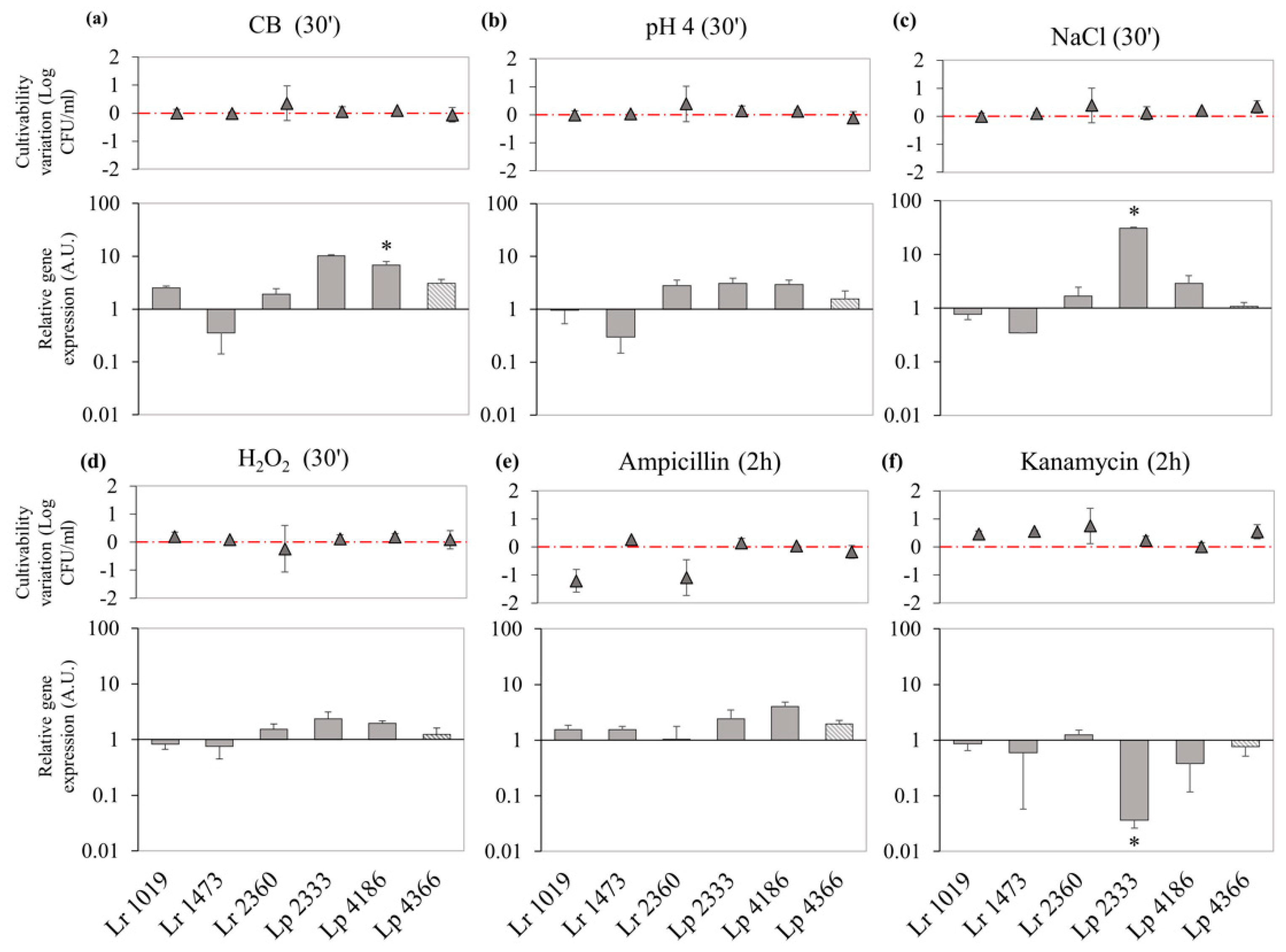
3.4. Effect of Food-Related Stress Conditions and Exposure to Antibiotic on DinJ-YafQ Expression
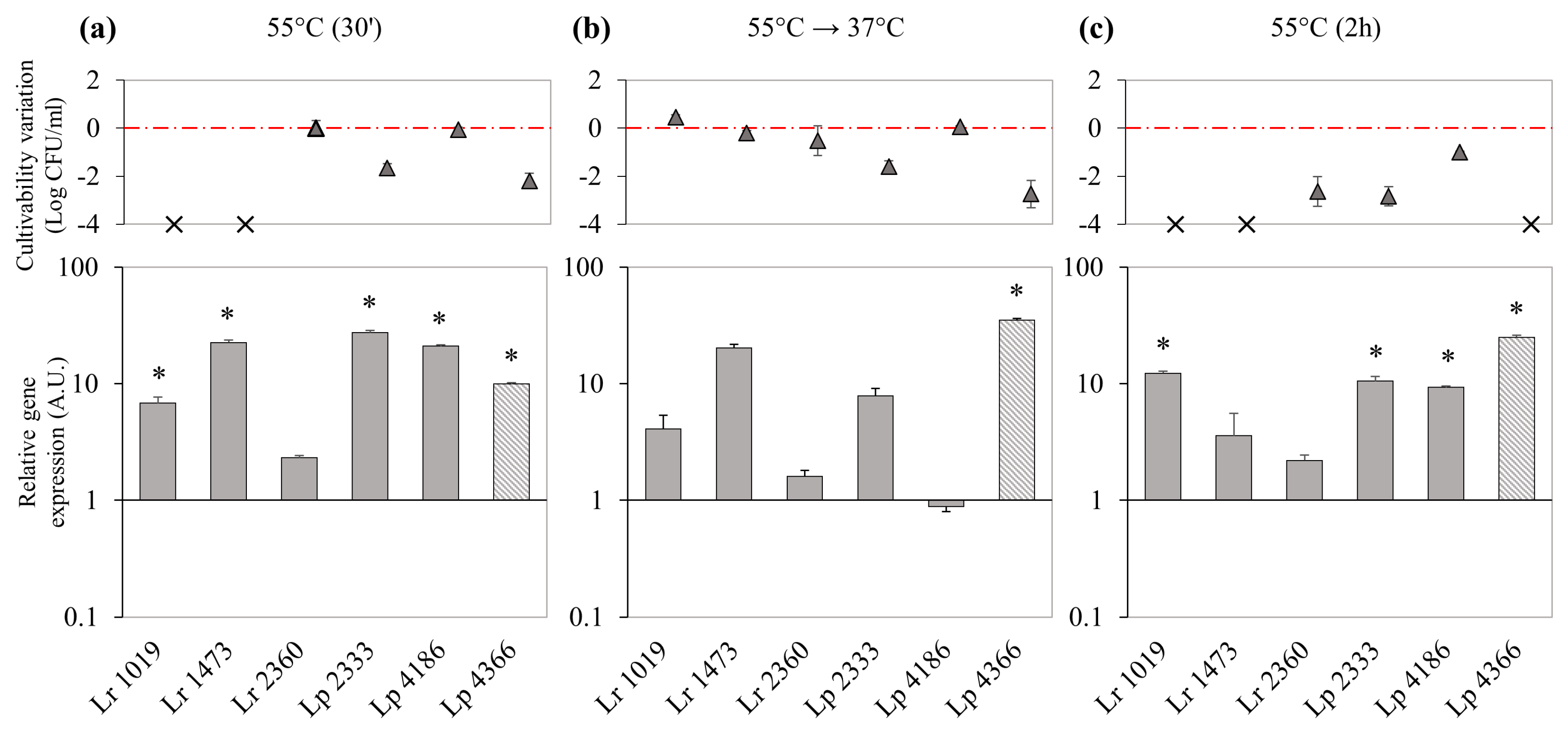
3.5. Effect of Thermal Stress on Bacterial Culturability and DinJ-YafQ Expression
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, targets, and triggers: An overview of toxin-antitoxin biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef] [PubMed]
- Amato, S.M.; Orman, M.A.; Brynildsen, M.P. Metabolic control of persister formation in Escherichia coli. Mol. Cell 2013, 50, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Leplae, R.; Geeraerts, D.; Hallez, R.; Guglielmini, J.; Dreze, P.; Van Melderen, L. Diversity of bacterial type II toxin-antitoxin systems: A comprehensive search and functional analysis of novel families. Nucleic Acids Res. 2011, 39, 5513–5525. [Google Scholar] [CrossRef] [PubMed]
- Pandey, D.P.; Gerdes, K. Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic Acids Res. 2005, 33, 966–976. [Google Scholar] [CrossRef] [PubMed]
- Duar, R.M.; Lin, X.X.B.; Zheng, J.S.; Martino, M.E.; Grenier, T.; Perez-Munoz, M.E.; Leulier, F.; Ganzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, J.R.; Neeno-Eckwall, E.C.; Stahl, B.; Tandee, K.; Cai, H.; Morovic, W.; Horvath, P.; Heidenreich, J.; Perna, N.T.; Barrangou, R.; et al. Analysis of the Lactobacillus casei supragenome and its influence in species evolution and lifestyle adaptation. BMC Genom. 2012, 13, 533. [Google Scholar] [CrossRef] [PubMed]
- Buron-Moles, G.; Chailyan, A.; Dolejs, I.; Forster, J.; Miks, M.H. Uncovering carbohydrate metabolism through a genotype-phenotype association study of 56 lactic acid bacteria genomes. Appl. Microbiol. Biot. 2019, 103, 3135–3152. [Google Scholar] [CrossRef] [Green Version]
- Ricci, A.; Cirlini, M.; Levante, A.; Dall’Asta, C.; Galaverna, G.; Lazzi, C. Volatile profile of elderberry juice: Effect of lactic acid fermentation using L-plantarum, L-rhamnosus and L-casei strains. Food Res. Int. 2018, 105, 412–422. [Google Scholar] [CrossRef]
- Bottari, B.; Levante, A.; Neviani, E.; Gatti, M. How the Fewest Become the Greatest. L. casei’s Impact on Long Ripened Cheeses. Front. Microbiol. 2018, 9, 2866. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; Di Cagno, R.; Calasso, M.; Neviani, E.; Fox, P.F.; De Angelis, M. Drivers that establish and assembly the lactic acid bacteria biota in cheeses. Trends Food Sci. Tech. 2018, 78, 244–254. [Google Scholar] [CrossRef]
- Jones, B.V. The human gut mobile metagenome: A metazoan perspective. Gut Microbes 2010, 1, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.V.; Sun, F.N.; Marchesi, J.R. Comparative metagenomic analysis of plasmid encoded functions in the human gut microbiome. BMC Genom. 2010, 11, 46. [Google Scholar] [CrossRef]
- Folli, C.; Levante, A.; Percudani, R.; Amidani, D.; Bottazzi, S.; Ferrari, A.; Rivetti, C.; Neviani, E.; Lazzi, C. Toward the identification of a type I toxin-antitoxin system in the plasmid DNA of dairy Lactobacillus rhamnosus. Sci. Rep. 2017, 7, 12051. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, A.; Maggi, S.; Montanini, B.; Levante, A.; Lazzi, C.; Yamaguchi, Y.; Rivetti, C.; Folli, C. Identification and first characterization of DinJ-YafQ toxin-antitoxin systems in Lactobacillus species of biotechnological interest. Sci. Rep. 2019, 9, 7645. [Google Scholar] [CrossRef] [PubMed]
- Klimina, K.M.; Kjasova, D.K.; Poluektova, E.U.; Krugel, H.; Leuschner, Y.; Saluz, H.P.; Danilenko, V.N. Identification and characterization of toxin-antitoxin systems in strains of Lactobacillus rhamnosus isolated from humans. Anaerobe 2013, 22, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Krugel, H.; Klimina, K.M.; Mrotzek, G.; Tretyakov, A.; Schofl, G.; Saluz, H.P.; Brantl, S.; Poluektova, E.U.; Danilenko, V.N. Expression of the toxin-antitoxin genes yefM(Lrh), yoeB(Lrh) in human Lactobacillus rhamnosus isolates. J. Basic Microb. 2015, 55, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Park, J.H.; Inouye, M. Toxin-Antitoxin Systems in Bacteria and Archaea. Annu. Rev. Genet. 2011, 45, 61–79. [Google Scholar] [CrossRef] [Green Version]
- Gatti, M.; Bottari, B.; Lazzi, C.; Neviani, E.; Mucchetti, G. Invited review: Microbial evolution in raw-milk, long-ripened cheeses produced using undefined natural whey starters. J. Dairy Sci. 2014, 97, 573–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. Clustal-W–Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Nicholas, K.B.; Nicholas, H.B. GeneDoc: A tool for editing and annotating multiple sequence alignments. Ver. 2.7.000. 1996. Distributed by the author. Available online: https://www.semanticscholar.org.
- Neviani, E.; Lindner, J.D.; Bernini, V.; Gatti, M. Recovery and differentiation of long ripened cheese microflora through a new cheese-based cultural medium. Food Microbiol. 2009, 26, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.J.; Li, Y.; Gao, P.F.; Sun, Z.H.; Sun, T.S.; Zhang, H.P. Validation of reference genes for real-time quantitative PCR studies in gene expression levels of Lactobacillus casei Zhang. J. Ind. Microbiol. Biot. 2011, 38, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034-1. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Prysak, M.H.; Mozdzierz, C.J.; Cook, A.M.; Zhu, L.; Zhang, Y.L.; Inouye, M.; Woychik, N.A. Bacterial toxin YafQ is an endoribonuclease that associates with the ribosome and blocks translation elongation through sequence-specific and frame-dependent mRNA cleavage. Mol. Microbiol. 2009, 71, 1071–1087. [Google Scholar] [CrossRef]
- Maehigashi, T.; Ruangprasert, A.; Miles, S.J.; Dunham, C.M. Molecular basis of ribosome recognition and mRNA hydrolysis by the E. coli YafQ toxin. Nucleic Acids Res. 2015, 43, 8002–8012. [Google Scholar] [CrossRef]
- Schuster, C.F.; Bertram, R. Toxin-antitoxin systems are ubiquitous and versatile modulators of prokaryotic cell fate. FEMS Microbiol. Lett. 2013, 340, 73–85. [Google Scholar] [CrossRef]
- Lazzi, C.; Turroni, S.; Mancini, A.; Sgarbi, E.; Neviani, E.; Brigidi, P.; Gatti, M. Transcriptomic clues to understand the growth of Lactobacillus rhamnosus in cheese. BMC Microbiol. 2014, 14, 28. [Google Scholar] [CrossRef]
- Toh, H.; Oshima, K.; Nakano, A.; Takahata, M.; Murakami, M.; Takaki, T.; Nishiyama, H.; Igimi, S.; Hattori, M.; Morita, H. Genomic Adaptation of the Lactobacillus casei Group. PLoS ONE 2013, 8, e75073. [Google Scholar] [CrossRef]
- Hu, Y.; Kwan, B.W.; Osbourne, D.O.; Benedik, M.J.; Wood, T.K. Toxin YafQ increases persister cell formation by reducing indole signalling. Environ. Microbiol. 2015, 17, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Brzozowska, I.; Zielenkiewicz, U. Regulation of toxin-antitoxin systems by proteolysis. Plasmid 2013, 70, 33–41. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Species | Source | Ripening Time (Months) |
|---|---|---|---|
| ATCC 334 | L. paracasei | Cheese | unknown |
| 2333 | L. paracasei | Cheese | 6 |
| 4186 | L. paracasei | Cheese | 4 |
| 4366 | L. paracasei | Raw cow’s milk | 0 |
| 1019 | L. rhamnosus | Cheese | 4 |
| 1473 | L. rhamnosus | Cheese | 20 |
| 2360 | L. rhamnosus | Cheese | 13 |
| Name | Sequence (5′–3′) | Strains | Reference |
|---|---|---|---|
| PLr2360 FW | CGGACAATTTTATATCGACCG | L. rhamnosus 2360 | this work |
| dinj-yafQ_rh6 minus | TTACTCAATGTTCAATGTATCGCG | [14] | |
| PLp4366 FW | ATACTATGTCGGTAAGGTCAG | L. paracasei 4366 | this work |
| dinj-yafQ_ca4_pa3 minus | AAGGTTATGATGAGATCCGGTTC | [14] |
| Condition | Label | Media | Temperature (°C) | Time (min) |
|---|---|---|---|---|
| Control | C | MRS pH 6.4 | 37 | 0 |
| Nutritional stress | CB | Cheese Broth (1) | 37 | 30 |
| Acidic stress | pH 4 | MRS pH 4 | 37 | 30 |
| Osmotic stress | NaCl | MRS pH 6.4, NaCl 1.5% (w/v) | 37 | 30 |
| Oxidative stress | H2O2 | MRS pH 6.4, H2O2 1 μM | 37 | 30 |
| Thermal stress | 55 °C | MRS pH 6.4 | 55 | 30 |
| 55 °C 2 h | MRS pH 6.4 | 55 | 120 | |
| Thermal stress relief | 55–37 °C | MRS pH 6.4 | 55 | 30 |
| 37 | 90 | |||
| Antibiotic exposure | Amp | MRS pH 6.4, ampicillin 0.1 mg/mL | 37 | 120 |
| Kan | MRS pH 6.4, kanamycin 0.05 mg/mL | 37 | 120 |
| Name | Sequence (5′–3′) 1 | Target Gene | Species | E% | Slope | R2 |
|---|---|---|---|---|---|---|
| GyrBFW | GCMCAGCCRCCGTTGTATCG | gyrB | L. rhamnosus L. paracasei | 97.75 | −3.38 | 0.999 |
| GyrBRV | GYTGGCGTCCATTTCMCCAAG | |||||
| GapdH1FW | GTTGGTACCATGACCACCGT | gapdh-1 | L. rhamnosus L. paracasei | 103.65 | −3.25 | 0.998 |
| GapdH1RV | GTGCTGTGAGGAATCGTGTT | |||||
| RecAFW | GATGATGCACTTGGTGTTGG | recA | L. rhamnosus L. paracasei | 96.99 | −3.40 | 0.999 |
| RecARV | TCRGCATCAATATARGCGG | |||||
| TBAFW 2 | CGGCAACGAGCGCAACCC | 16S rRNA | L. rhamnosus L. paracasei | 99.79 | −3.33 | 0.999 |
| TBARV 2 | CCATTGTAGCACGTGTGTAGCC | |||||
| YafQ_lrFW | TGCAGCGTCAAGGTCATGTA | dinJ-yafQ | L. rhamnosus | 95.64 | −3.43 | 0.998 |
| YafQ_lrRV | CAATGTATCGCGGTGTGTGC | |||||
| YafQ_lpFW | GCCGATGGACGAACTAAAGA | dinJ-yafQ | L. paracasei | 98.76 | −3.35 | 0.995 |
| YafQ_lpRV | TATCCTTTCCACTCGCTGCT |
| Strain | Target | Min (Ct) | Max (Ct) | Range | M |
|---|---|---|---|---|---|
| ATCC 334 | 16S rRNA | 7.62 | 9.54 | 1.92 | – |
| gyrB | 17.18 | 19.79 | 2.61 | 0.47 | |
| gapdh-1 | 13.10 | 17.54 | 4.44 | 1.36 | |
| recA | 20.15 | 23.79 | 3.64 | 7.43 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levante, A.; Folli, C.; Montanini, B.; Ferrari, A.; Neviani, E.; Lazzi, C. Expression of DinJ-YafQ System of Lactobacillus casei Group Strains in Response to Food Processing Stresses. Microorganisms 2019, 7, 438. https://doi.org/10.3390/microorganisms7100438
Levante A, Folli C, Montanini B, Ferrari A, Neviani E, Lazzi C. Expression of DinJ-YafQ System of Lactobacillus casei Group Strains in Response to Food Processing Stresses. Microorganisms. 2019; 7(10):438. https://doi.org/10.3390/microorganisms7100438
Chicago/Turabian StyleLevante, Alessia, Claudia Folli, Barbara Montanini, Alberto Ferrari, Erasmo Neviani, and Camilla Lazzi. 2019. "Expression of DinJ-YafQ System of Lactobacillus casei Group Strains in Response to Food Processing Stresses" Microorganisms 7, no. 10: 438. https://doi.org/10.3390/microorganisms7100438
APA StyleLevante, A., Folli, C., Montanini, B., Ferrari, A., Neviani, E., & Lazzi, C. (2019). Expression of DinJ-YafQ System of Lactobacillus casei Group Strains in Response to Food Processing Stresses. Microorganisms, 7(10), 438. https://doi.org/10.3390/microorganisms7100438