Phylogeny of Anoxygenic Photosynthesis Based on Sequences of Photosynthetic Reaction Center Proteins and a Key Enzyme in Bacteriochlorophyll Biosynthesis, the Chlorophyllide Reductase



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Cultivation, Sequencing, and Assembly of DNA Sequences
2.2. Sequences
2.3. Phylogenetic Analyses
3. Results and Discussion
3.1. Strain and Sequence Selection
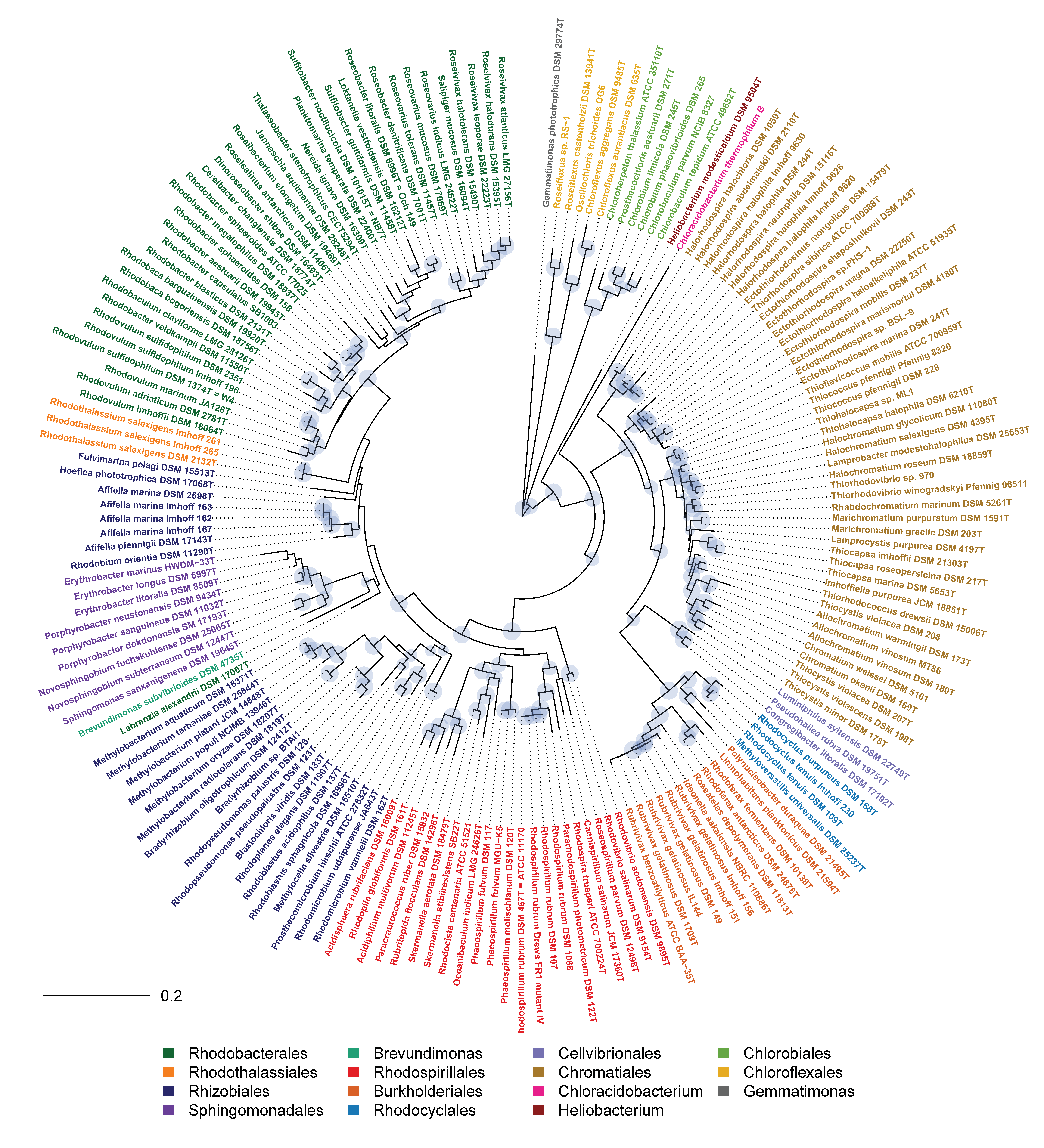
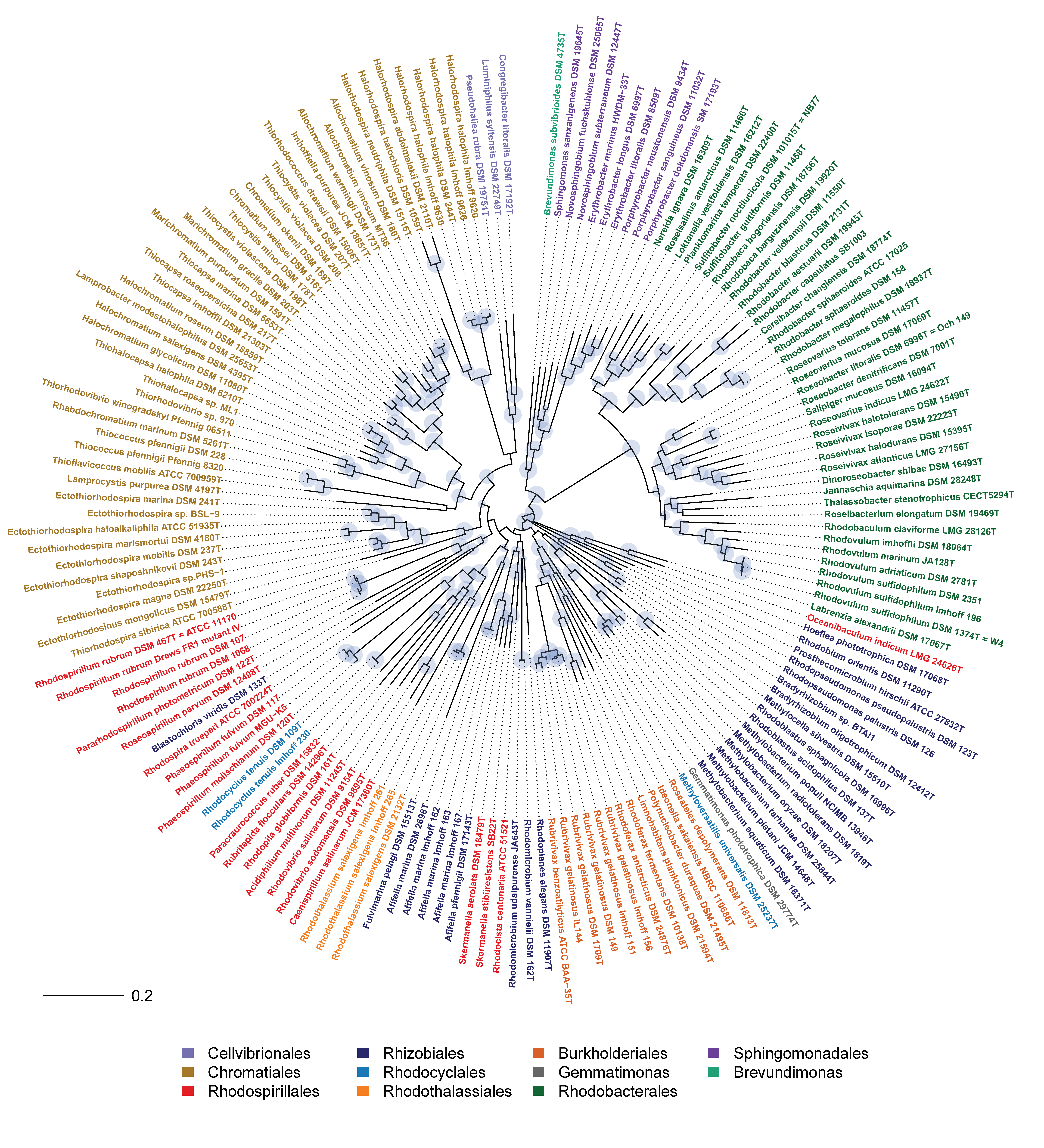
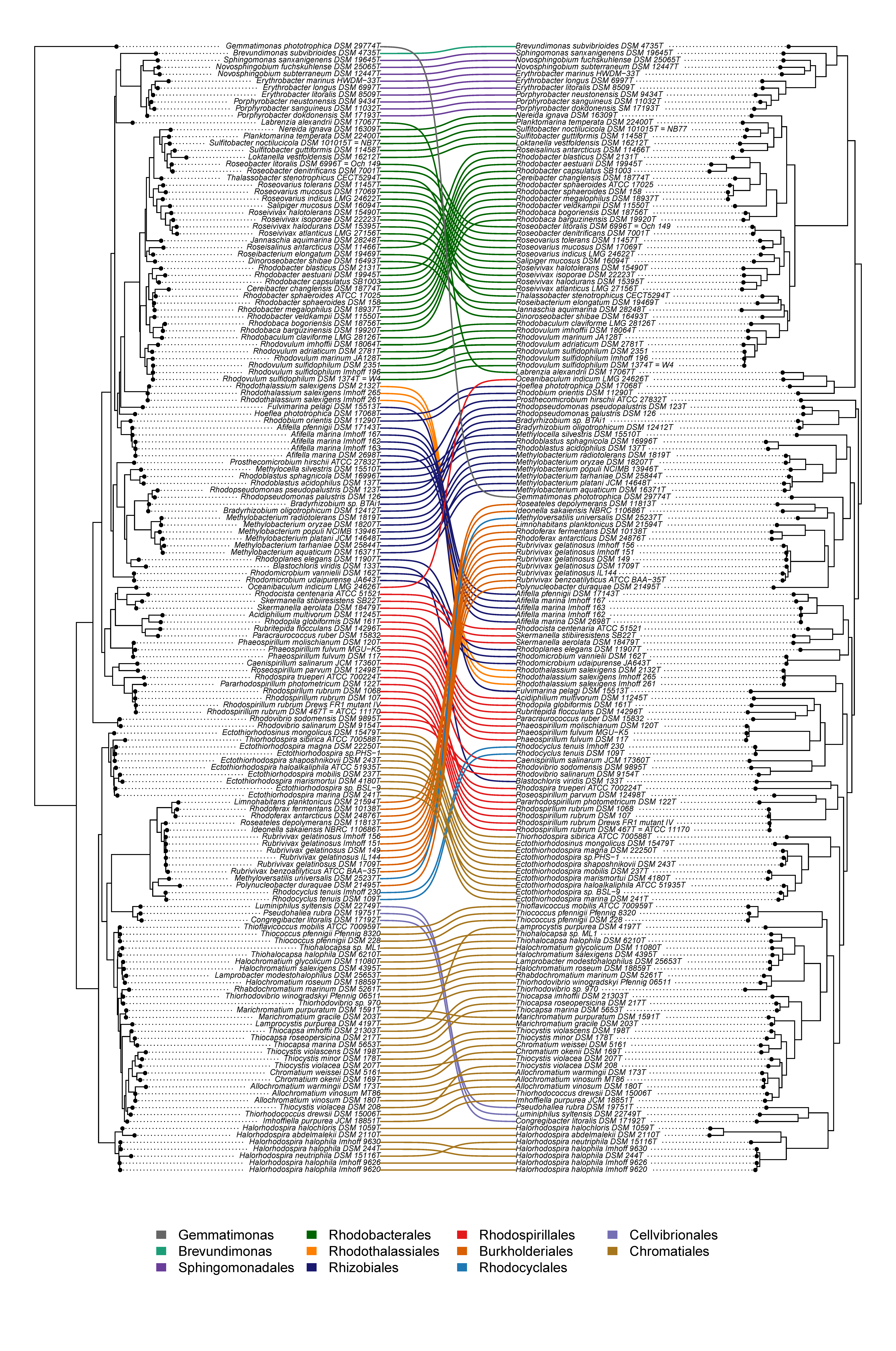
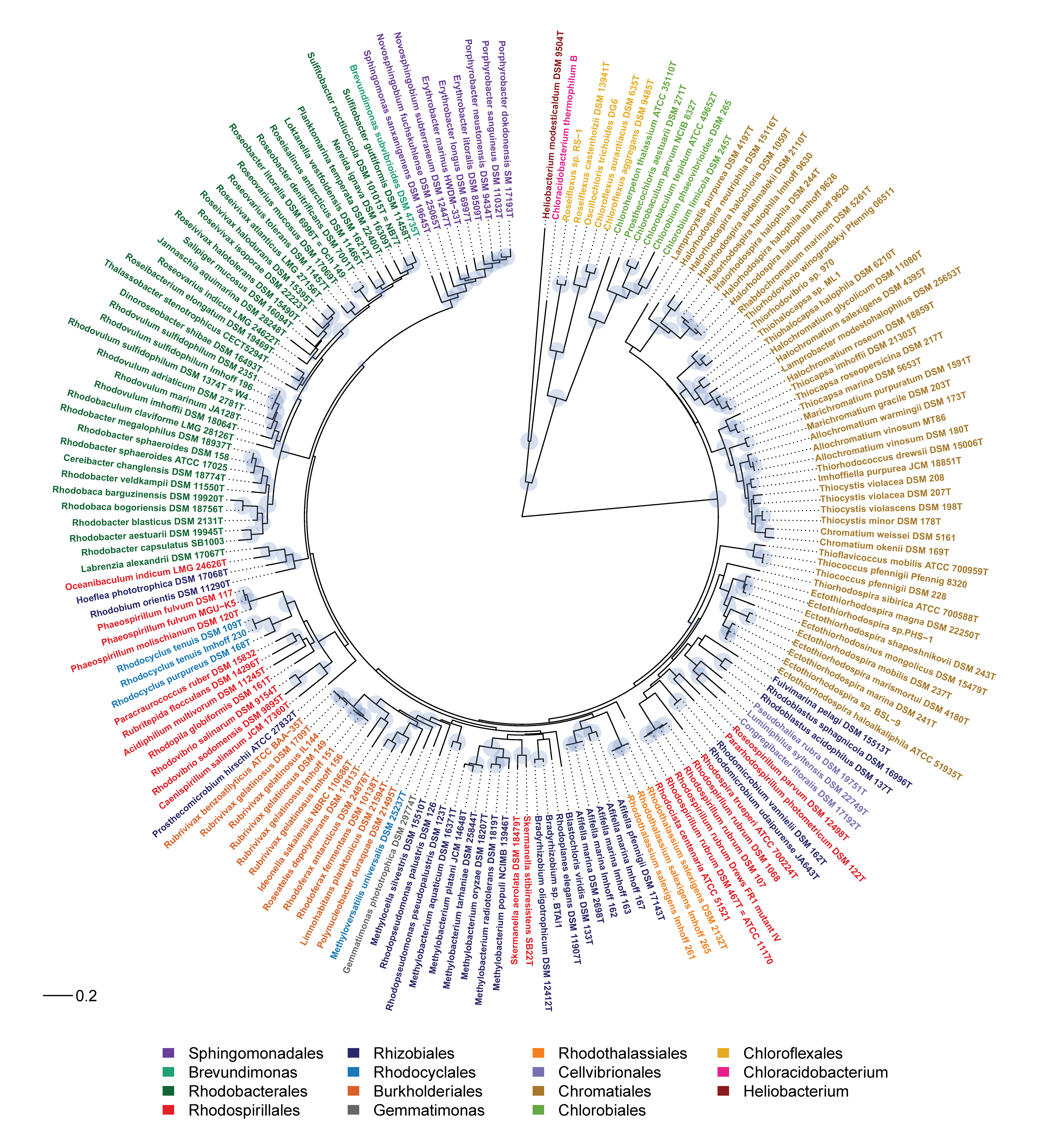
3.2. Phylogeny According to 16S rRNA Gene Sequences
- (i)
- Rhodospirillum, Roseospirillum/Caenispirillum, Rhodospira, Pararhodospirillum,
- (ii)
- Phaeospirillum, Oceanibaculum, Rhodocista, Skermanella,
- (iii)
- Rhodopila, Rubritepida, Paracraurococcus, Acidiphilum, Acidisphaera,
- (iv)
- Erythrobacter, Porphyrobacter, Novosphingobium, Sphingomonas,
- (v)
- Rhodopseudomonas/Bradyrhizobium, Methylobacterium.
3.3. Phylogeny of Photosynthesis
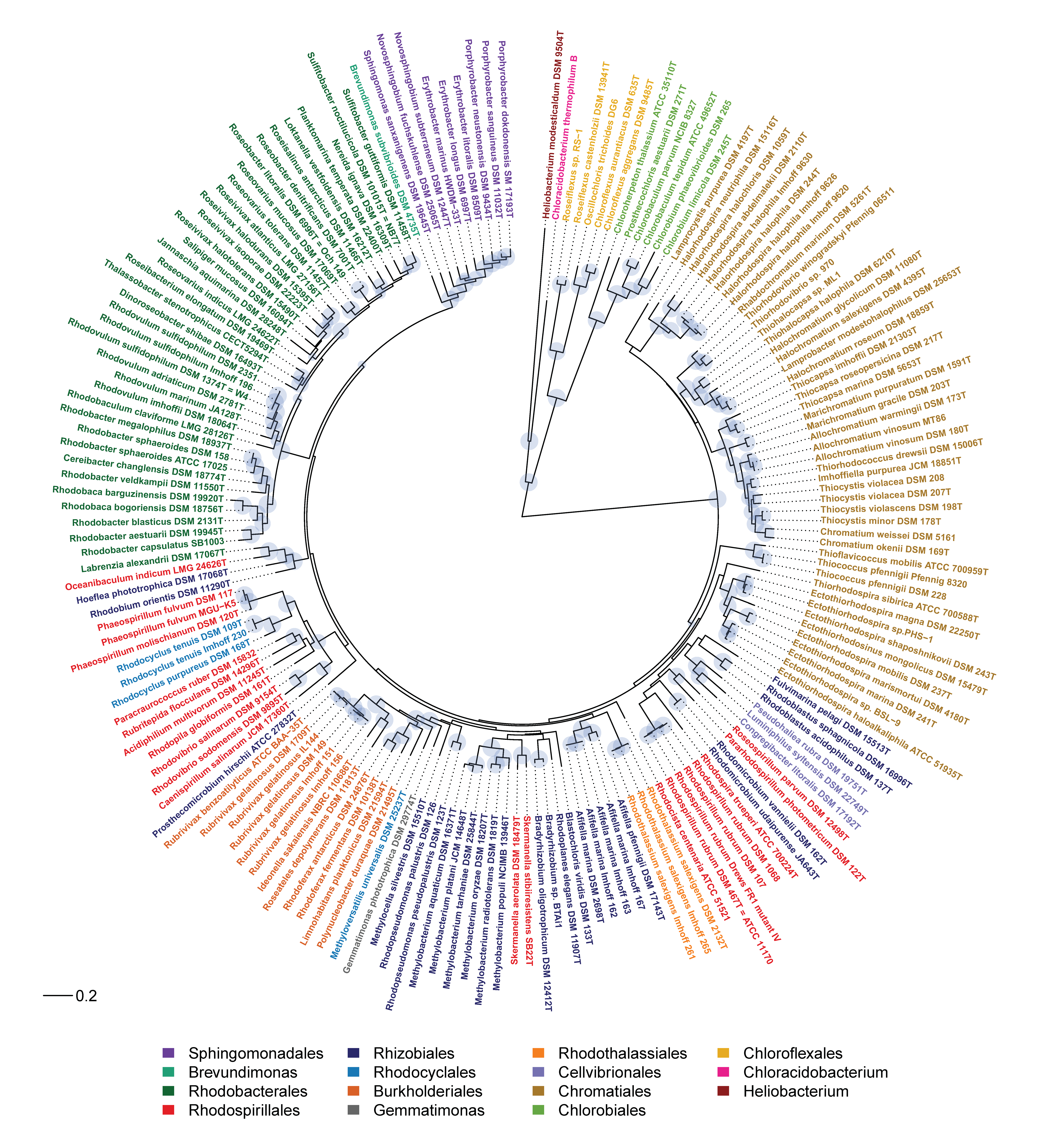
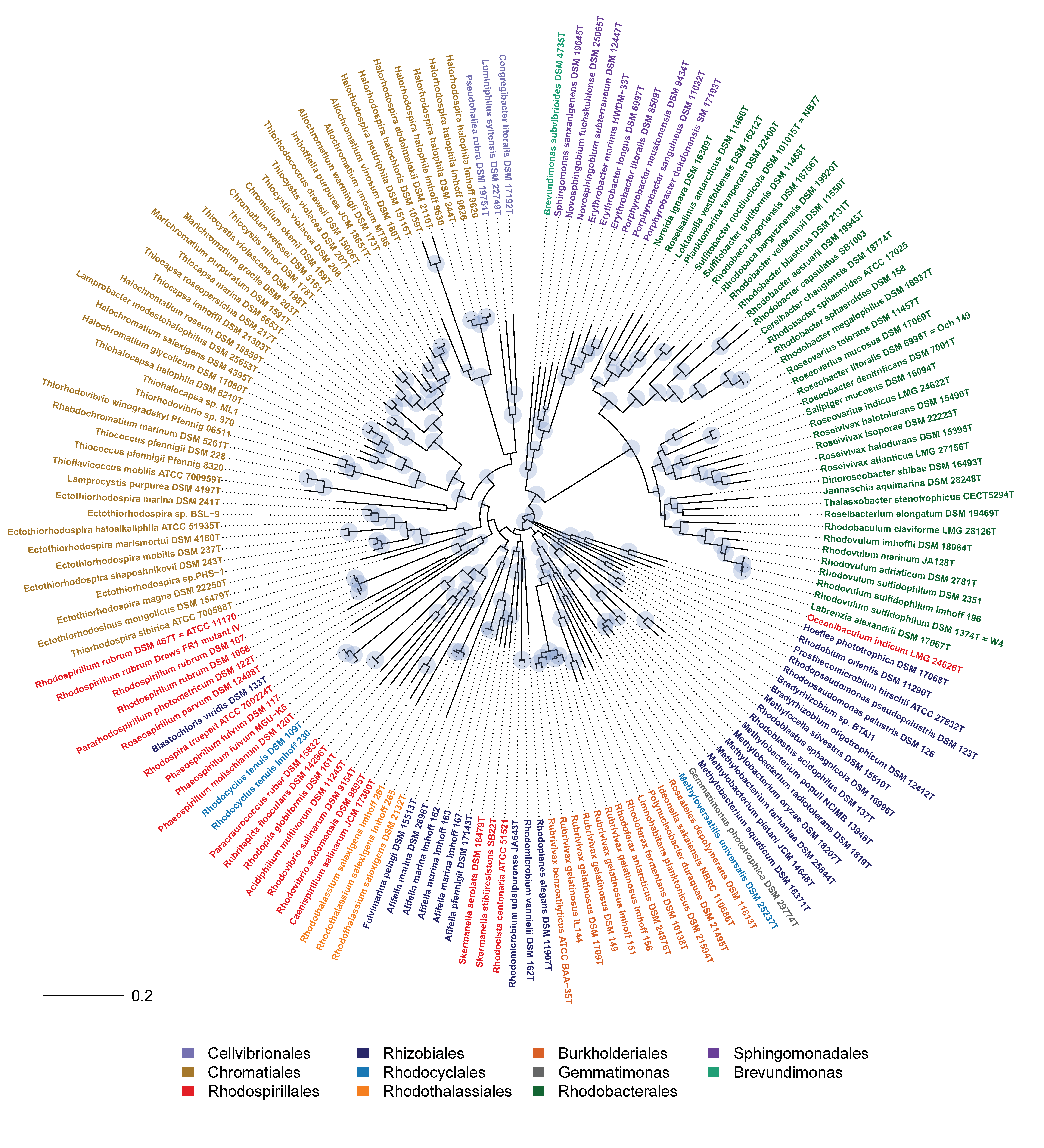
3.3.1. Phylogeny according to BchXYZ Sequences
3.3.2. Phylogeny of BchXYZ-PufHLM and Comparison with 16S rRNA Phylogeny
Gammaproteobacteria (Chromatiales and Cellvibrionales)
- (i)
- the Halorhodospira species,
- (ii)
- the Ectothiorhodospiraceae, including Ectothiorhodospira and Ectothiorhodosinus species, with Thiorhodospira being associated more distantly, but excluding the Halorhodospira species,
- (iii)
- the Chromatiaceae with subgroups of a) the Thiococcus group of bacteriochlorophyll-b containing Chromatiaceae, including species of Thiococcus and Thioflavicoccus; b) the Halochromatium group with halophilic species of the genera Halochromatium, Lamprobacter, Rhabdochromatium, Thiorhodovibrio, and Thiohalocapsa; c) the Chromatium group with species of Chromatium, Thiocapsa, Marichromatium, Allochromatium, Thiorhodococcus, Imhoffiella, and Thiocystis; d) Lamprocystis purpurea as an outsider among the Chromatiaceae with distant relationship to others and no statistical support of its position. Most significantly, Lamprocystis purpurea formed a deeply branching line in the Chromatium-group according to both 16S rRNA phylogeny and PS phylogeny. Therefore, it is likely to be an ancient representative of the Chromatiaceae,
- (iv)
- the Cellvibrionales (Haliaceae) with Congregibacter litoralis, Luminiphilus syltensis, and Pseudohaliea rubra (most likely including Chromatocurvus halotolerans [3]), which were linked with low confidence to the Halorhodospira group. The Cellvibrionales formed a group distant to other Gammaproteobacteria according to both trees. In the RNA tree, they were linked to the Betaproteobacteria (in this tree within the frame of the Gammaproteobacteria), and in the PS tree, associated with the Halorhodospira group. Apparently, they represent an ancient phylogenetic lineage of the Gammaproteobacteria without clearly resolved roots.
Betaproteobacteria (Burkholderiales and Rhodocyclales)
Gemmatimonas (Gemmatimonadales)
Alphaproteobacteria
- -
- Rhodovulum group: According to BchXYZ and BchXYZ-PufHLM, the Rhodovulum group was well recognized. Rhodobaculum claviforme appeared distantly associated with this group and, like the Rhodovulum species, had PufC (Supplementary Table S1). However, in the RNA tree, Rhodobaculum claviforme clustered with Rhodobacter species.
- -
- Rhodobacter/Rhodobaca group: This group contained Rhodobacter and Rhodobaca species together with Cereibacter changlensis and was supported by all considered trees (BchXYZ-PufHLM, BchXYZ, RNA tree). The reaction center cytochrome PufC was absent (Supplementary Table S1). Quite remarkable Rhodobaculum claviforme was included in this group according to the RNA tree only.
- -
- Loktanella/Sulfitobacter group: This group included species of Loktanella, Sulfitobacter, Planktomarina, and Roseisalinus and distantly linked also Nereida ignava. It was supported by BchXYZ-PufHLM and lacked PufC (Supplementary Table S1). According to the RNA tree, this group was not well supported, and Roseobacter but not Roseisalinus was included.
- -
- Roseobacter/Roseivivax group: This group contained species of Roseobacter, Roseivivax, Salipiger, and Roseovarius. In line with the PS tree, PufC was present in all species, including Roseobacter. The RNA tree excluded Roseobacter from this group.
- -
- Dinoroseobacter/Jannaschia group: Dinoroseobacter shibae, Jannaschia aquamarina, Thalassobacter stenotrophicus, and Roseibacterium elongatum formed a group of poorly linked bacteria, which did not fit into any of the aforementioned groups. All four species had PufC. Within the RNA tree, there was only weak support for this group (Figure 1).
- -
- Rhodocista centenaria and Skermanella species showed up jointly in the RNA tree with the Acetobacteraceae as a sister branch, while both formed a deeply rooted unsupported branch in the PS tree.
- -
- Rhodomicrobium formed a distinct lineage within the Rhizobiales in the RNA tree but, according to the PS tree, separated from other Rhizobiales together with Rhodoplanes in a distinct deeply branching but unsupported line.
- -
- Blastochloris separated from other Rhizobiales in the PS tree and formed an unsupported isolated line together with the bacteriochlorophyll-b containing Rhodospira trueperi, while it was included in a major branch of Rhizobiales in the RNA tree.
3.4. Distribution of PufC
3.5. Phylogenetic Aspects of Aerobic Anoxygenic Photosynthesis
3.6. General Aspects
4. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Madigan, M.T.; Schaaf, N.A.V.; Sattley, W.M. The Chlorobiaceae, Chloroflexaceae, and Heliobacteriaceae. In Modern Topics in the Phototrophic Prokaryotes: Environmental and Applied Aspects; Hallenbeck, P.C., Ed.; Springer: Cham, Switzerland, 2017; pp. 139–161. [Google Scholar]
- Imhoff, J.F. Diversity of anaerobic anoxygenic phototrophic purple bacteria. In Modern Topics in the Phototrophic Prokaryotes: Environmental and Applied Aspects; Hallenbeck, P.C., Ed.; Springer: Cham, Switzerland, 2017; pp. 47–85. [Google Scholar]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Neulinger, S.C. Photosynthesis is widely distributed among proteobacteria as revealed by the phylogeny of PufLM reaction center proteins. Front. Microbiol. 2018, 8, 2679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sekiguchi, Y.; Hanada, S.; Hugenholtz, P.; Kim, H.; Kamagata, Y.; Nakamura, K. Gemmatimonas aurantiaca gen. nov., sp. nov., a Gram-negative, aerobic, polyphosphate-accumulating microorganism, the first cultured representative of the new bacterial phylum Gemmatimonadetes phyl. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Zeng, A.; Koblížek, M. Phototrophic Gemmatimonadetes: A new “purple” branch of the bacterial tree of life. In Modern Topics in the Phototrophic Prokaryotes: Environmental and Applied Aspects; Hallenbeck, P.C., Ed.; Springer: Cham, Switzerland, 2017; pp. 163–192. [Google Scholar]
- Nagashima, S.; Nagashima, K.V.P. Comparison of photosynthrtic gene clusters retrieved from total genome sequences of purple bacteria. In Genome Evolution of Photosynthetic Bacteria; Beatty, T.J., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 151–178. [Google Scholar]
- Tank, M.; Blümel, M.; Imhoff, J.F. Communities of purple sulfur bacteria in a Baltic Sea coastal lagoon analyzed by pufLM gene libraries and the impact of temperature and NaCl concentration in experimental enrichment cultures. FEMS Microbiol. Ecol. 2011, 78, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Tank, M.; Thiel, V.; Imhoff, J.F. Phylogenetic relationship of phototrophic purple sulfur bacteria according to pufL and pufM genes. Intern. Microbiol. 2009, 12, 175–185. [Google Scholar]
- Thiel, V.; Tank, M.; Neulinger, S.C.; Gehrmann, L.; Dorador, C.; Imhoff, J.F. Unique communities of anoxygenic phototrophic bacteria in saline lakes of Salar de Atacama (Chile). Evidence for a new phylogenetic lineage of phototrophic Gammaproteobacteria from pufLM gene analyses. FEMS Microbiol. Ecol. 2010, 74, 510–522. [Google Scholar] [CrossRef]
- Willows, R.D.; Kriegel, A.M. Biosynthesis of bacteriochlorophylls in purple bacteria. In Advances in Photosynthesis and Respiration. The Purple Phototrophic Bacteria; Hunter, C.N., Daldal, F., Thurnauer, M.C., Beatty, J.T., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 57–79. [Google Scholar]
- Pfennig, N.; Trüper, H.G. Isolation of members of the families Chromatiaceae and Chlorobiaceae. In The Prokaryotes, A Handbook on Habitats, Isolation and Identification of Bacteria; Starr, M.P., Stolp, H., Trüper, H.G., Balows, A., Schlegel, H.G., Eds.; Springer: New York, NY, USA, 1981; pp. 279–289. [Google Scholar]
- Imhoff, J.F. The Chromatiaceae. In The Prokaryotes. A Handbook on the Biology of Bacteria, 3rd ed.; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 6, pp. 846–873. [Google Scholar]
- Imhoff, J.F. Anoxygenic phototrophic bacteria. In Methods in Aquatic Bacteriology; Austin, B., Ed.; John Wiley & Sons: Chichester, UK, 1988; pp. 207–240. [Google Scholar]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Neulinger, S.C. New insights into the metabolic potential of the phototrophic purple bacterium Rhodopila globiformis DSM 161T from its draft genome sequence and evidence for a vanadium-dependent nitrogenase. Arch. Microbiol. 2018, 200, 847–857. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.-I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Schliep, K.P. Phangorn: Phylogenetic analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef]
- Revell, L.J. Phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Woese, C.R. Bacterial evolution. Microbiol. Rev. 1987, 51, 4673–4680. [Google Scholar]
- Zeng, A.; Feng, F.; Medova, H.; Dean, J.; Koblizek, M. Functional type 2 photosynthetic reaction centers found in the rare bacterial phylum Gemmatimonadetes. Proc. Natl. Acad. Sci. USA 2014, 111, 7795–7800. [Google Scholar] [CrossRef]
- Zeng, A.; Selyanin, V.; Lukes, M.; Dean, J.; Kaftan, D.; Feng, F.; Koblizek, M. Characterization of the microaerophilic, bacteriochlorophyll a-containing bacterium Gemmatimonas phototrophica sp. nov., and emended description of the genus Gemmatimonas and Gemmatimonas auranttiaca. Int. J. Syst. Evol. Microbiol. 2015, 65, 2410–2419. [Google Scholar] [CrossRef]
- Imhoff, J.F. Genus Roseospirillum. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; Volume 2, p. 39. [Google Scholar]
- Igarashi, N.; Harada, J.; Nagashima, S.; Matsuura, K.; Shimada, K.; Nagashima, K.V.P. Horizontal transfer of photosynthesis gene cluster and operon rearrangement in purple bacteria. J. Mol. Evol. 2001, 52, 333–341. [Google Scholar] [CrossRef]
- Holland, H.D. The oxygenation of the atmosphere and oceans. Phil. Trans. R. Soc. B 2006, 361, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, R.E. Early evolution of photosynthesis. Plant Physiol. 2010, 154, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Canfield, D.E. The early history of atmospheric oxygen: Homage to Robert, M. Garrels. Annu. Rev. Earth Planet. Sci. 2005, 33, 1–36. [Google Scholar] [CrossRef]
- Drews, G.; Imhoff, J.F. Phototrophic purple bacteria. In Variations in Autotrophic Life; Shively, J.M., Barton, L.L., Eds.; Academic Press: London, UK, 1991; pp. 51–97. [Google Scholar]
- Imhoff, J.F. Genus Rhodobacter. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; Volume 2, pp. 161–167. [Google Scholar]
- Imhoff, J.F. Genus Rhodovulum. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; Volume 2, pp. 205–209. [Google Scholar]
- Imhoff, J.F. The phototrophic Alphaproteobacteria. In The Prokaryotes. A Handbook on the Biology of Bacteria., 3rd ed.; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 5, pp. 41–64. [Google Scholar]
- Shimada, K. Aerobic anoxygenic phototrophs. In Anoxygenic Photosynthetic Bacteria; Blankenship, R.E., Madigan, M.T., Bauer, C.E., Eds.; Kluwer: Dordrecht, The Netherlands, 1995; pp. 105–122. [Google Scholar]
- Shiba, T. Roseobacter litoralis gen. nov., sp. nov., and Roseobacter denitrificans sp. nov., aerobic pink-pigmented bacteria which contain bacteriochlorophyll a. Syst. Appl. Microbiol. 1991, 14, 140–145. [Google Scholar] [CrossRef]
- Yurkov, V. Aerobic phototrophic Proteobacteria. In The Prokaryotes. A Handbook on the Biology of Bacteria, 3rd ed.; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 5, pp. 562–584. [Google Scholar]
- Koblížek, M.; Zeng, Y.; Horák, A.; Oborník, M. Regressive evolution of photosynthesis in the Roseobacter clade. In Genome Evolution of Photosynthetic Bacteria; Beatty, T.J., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 1385–1405. [Google Scholar]
- Brinkmann, H.; Göker, M.; Koblížek, M.; Wagner-Döbler, I.; Petersen, J. Horizontal operon transfer, plasmids, and the evolution of photosynthesis in Rhodobacteraceae. ISME J. 2018, 12, 1994–2010. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imhoff, J.F.; Rahn, T.; Künzel, S.; Neulinger, S.C. Phylogeny of Anoxygenic Photosynthesis Based on Sequences of Photosynthetic Reaction Center Proteins and a Key Enzyme in Bacteriochlorophyll Biosynthesis, the Chlorophyllide Reductase. Microorganisms 2019, 7, 576. https://doi.org/10.3390/microorganisms7110576
Imhoff JF, Rahn T, Künzel S, Neulinger SC. Phylogeny of Anoxygenic Photosynthesis Based on Sequences of Photosynthetic Reaction Center Proteins and a Key Enzyme in Bacteriochlorophyll Biosynthesis, the Chlorophyllide Reductase. Microorganisms. 2019; 7(11):576. https://doi.org/10.3390/microorganisms7110576
Chicago/Turabian StyleImhoff, Johannes F., Tanja Rahn, Sven Künzel, and Sven C. Neulinger. 2019. "Phylogeny of Anoxygenic Photosynthesis Based on Sequences of Photosynthetic Reaction Center Proteins and a Key Enzyme in Bacteriochlorophyll Biosynthesis, the Chlorophyllide Reductase" Microorganisms 7, no. 11: 576. https://doi.org/10.3390/microorganisms7110576
APA StyleImhoff, J. F., Rahn, T., Künzel, S., & Neulinger, S. C. (2019). Phylogeny of Anoxygenic Photosynthesis Based on Sequences of Photosynthetic Reaction Center Proteins and a Key Enzyme in Bacteriochlorophyll Biosynthesis, the Chlorophyllide Reductase. Microorganisms, 7(11), 576. https://doi.org/10.3390/microorganisms7110576