Impact of Binge Alcohol Intoxication on the Humoral Immune Response during Burkholderia spp. Infections



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals for Study
2.2. Bacterial Preparation
2.3. Experimental Design, Alcohol Administration, and Blood Collection
2.4. Lipopolysaccharide (LPS), Capsular Polysaccharide (CPS), and Whole Cell Lysate (WCL) Preparation
2.5. Enzyme-Linked Immunosorbent Assay
2.6. MagPix Detection Assays: Coupling of Antigen to Beads
2.7. Bead-Based Assay for IgM and IgG Antibodies
2.8. Data Analysis
3. Results
3.1. Effect of Binge Alcohol Intoxication on Mouse Survival Following Infection with Burkholderia Species
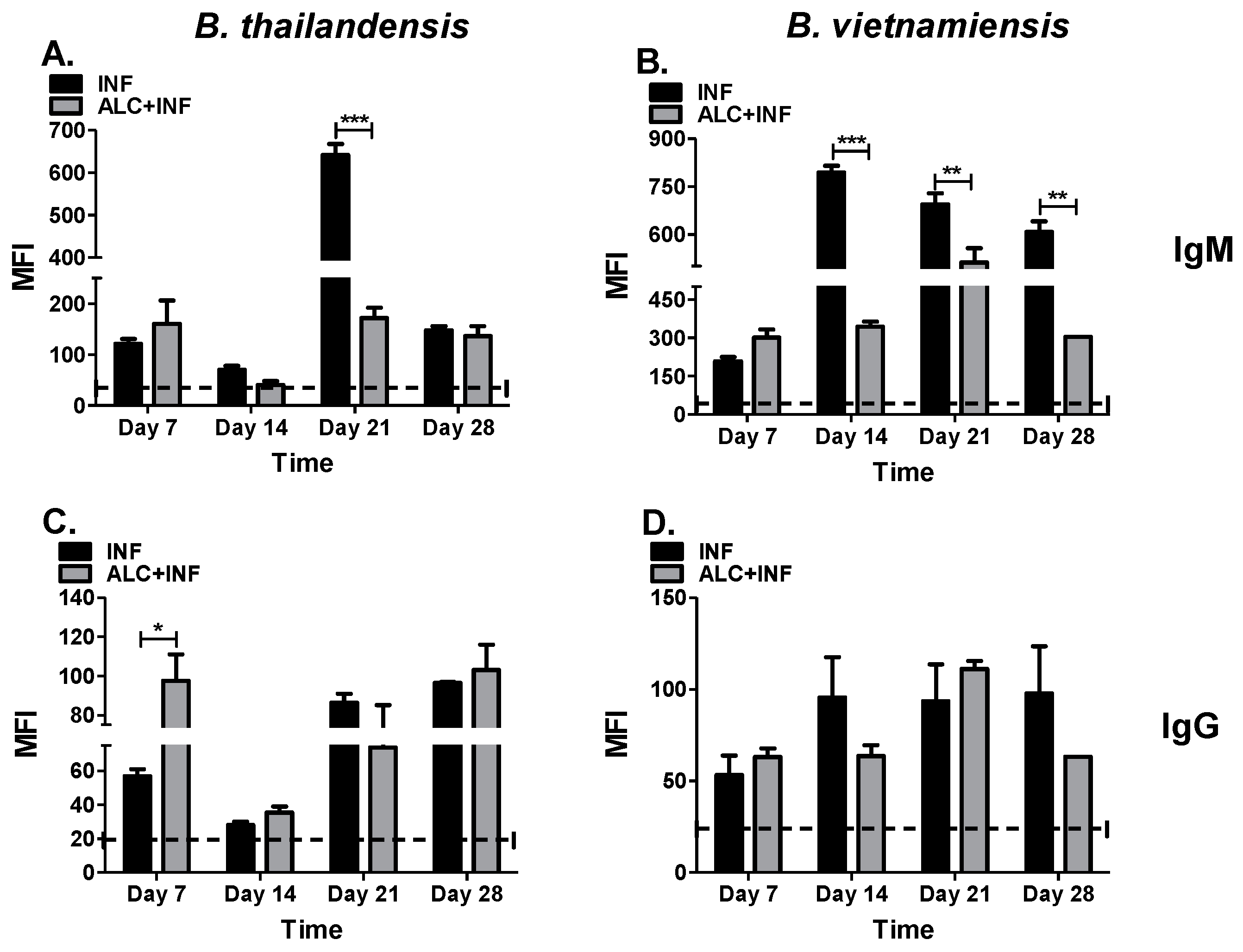
3.2. Binge Alcohol Selectively Suppresses Total IgM Concentration in B. thailandensis but Not in B. vietnamiensis Infections
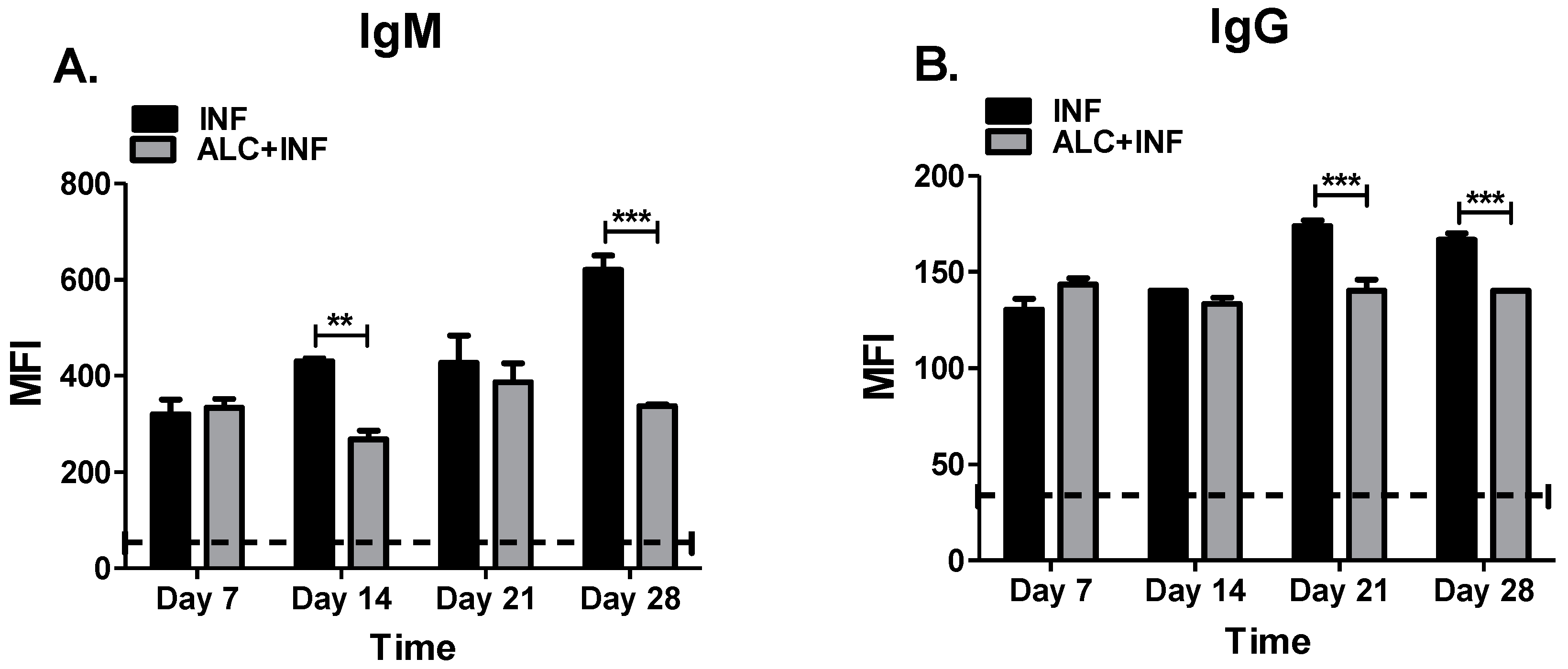
3.3. Binge Alcohol Delays Total IgG Concentrations during Burkholderia Infection
3.4. Binge Alcohol Elevates IgM and Delays IgG Specific to the Whole Cell Lysate (WCL) of B. vietnamiensis but Not B. thailandensis Infections
3.5. Binge Alcohol Suppresses IgM and IgG Specific to the Capsular Polysaccharide (CPS) of B. vietnamiensis
3.6. Binge Alcohol Suppresses IgM Specific to the Lipopolysaccharide-A (LPS-A) of B. thailandensis and B. vietnamiensis, While IgG Remained Unchanged
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Limmathurotsakul, D.; Golding, N.; Dance, D.A.B.; Messina, J.P.; Pigott, D.M. Predicted global distribution of Burkholderia pseudomallei and burden of melioidosis. Nat. Microbiol. 2016, 1, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Currie, B.J.; Fisher, D.A.; Howard, D.M.; Burrow, J.N.C.; Lo, D.; Selva-nayagam, S.; Krause, V.L. Endemic melioidosis in tropical northern Australia: A 10-Year prospective study and review of the literature. Clin. Infect. Dis. 2000, 31, 981–986. [Google Scholar] [CrossRef]
- Currie, B.J. Melioidosis: Evolving concepts in epidemiology, pathogenesis, and treatment. Semin. Respir. Crit. Care Med. 2015, 36, 111–125. [Google Scholar] [CrossRef]
- Currie, B.J.; Jacups, S.P.; Cheng, A.C.; Fisher, D.A.; Anstey, N.M.; Huffam, S.E.; Krause, V.L. Melioidosis epidemiology and risk factors from a prospective whole-population study in northern Australia. Trop. Med. Int. Health 2004, 9, 1167–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currie, B.J.; Ward, L.; Cheng, A.C. The epidemiology and clinical spectrum of melioidosis: 540 cases from the 20 year darwin prospective study. PLoS Negl. Trop. Dis. 2010, 4, e900. [Google Scholar] [CrossRef]
- Trevejo-Nunez, G.; Kolls, J.K.; de Wit, M. Alcohol use as a risk factor in infections and healing: A clinician’s perspective. Alcohol Res. Curr. Rev. 2015, 37, 177–184. [Google Scholar]
- National Institutes of Alcohol Abuse and Alcoholism. Drinking Levels Defined. Available online: https://www.niaaa.nih.gov/alcohol-health/overview-alcohol-consumption/moderate-binge-drinking (accessed on 1 March 2019).
- Simet, S.M.; Sisson, J.H. Alcohol’s effects on lung health and immunity. Alcohol Res. 2015, 37, 199–208. [Google Scholar]
- Mehta, A.J.; Guidot, D.M. Alcohol and the lung. Alcohol Res. 2017, 38, 243–254. [Google Scholar]
- Jimenez, V.; Moreno, R.; Kaufman, E.; Hornstra, H.; Settles, E.; Currie, B.J.; Monroy, F.P. Effects of binge alcohol exposure on Burkholderia thailandensis–alveolar macrophage interaction. Alcohol 2017, 64, 55–63. [Google Scholar] [CrossRef]
- Boé, D.M.; Nelson, S.; Zhang, P.; Bagby, G.J. Acute ethanol intoxication suppresses lung chemokine production following Infection with Streptococcus pneumoniae. J. Infect. Dis. 2001, 184, 1134–1142. [Google Scholar] [CrossRef] [PubMed]
- Szabo, G.; Saha, B. Alcohol′s effect on host defense. Alcohol Res. 2015, 37, 159–170. [Google Scholar] [PubMed]
- Happel, K.I.; Nelson, S. Alcohol, immunosuppression, and the lung. Proc. Am. Thorac. Soc. 2005, 2, 428–432. [Google Scholar] [CrossRef]
- Cook, R.T.; Ballas, Z.K.; Waldschmidt, T.J.; Vandersteen, D.; LaBrecque, D.R.; Cook, B.L. Modulation of T-cell adhesion markers, and the CD45R and CD57 antigens in human alcoholics. Alcohol. Clin. Exp. Res. 1995, 19, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Maruya, M.; Kawamoto, S.; Fagarasan, S. Roles of B-1 and B-2 cells in innate and acquired IgA-mediated immunity. Immunol. Rev. 2010, 237, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Haraga, A.; West, T.E.; Brittnacher, M.J.; Skerrett, S.J.; Miller, S.I. Burkholderia thailandensis as a model system for the study of the virulence-associated type III secretion system of Burkholderia pseudomallei. Infect. Immun. 2008, 76, 5402–5411. [Google Scholar] [CrossRef] [PubMed]
- Tuanyok, A.; Mayo, M.; Scholz, H.; Hall, C.M.; Allender, C.J.; Kaestli, M.; Ginther, J.; Spring-Pearson, S.; Bollig, M.C.; Stone, J.K.; et al. Burkholderia humptydooensis sp. nov., A new species related to Burkholderia thailandensis and the fifth member of the Burkholderia pseudomallei Complex. Appl. Environ. Microbiol. 2017, 83, e02802–e02816. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, V.; Moreno, R.; Settles, E.; Currie, B.J.; Keim, P.; Monroy, F.P. A mouse model of binge alcohol consumption and Burkholderia infection. PLoS ONE 2018, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Burtnick, M.N.; Heiss, C.; Roberts, R.A.; Schweizer, H.P.; Azadi, P.; Brett, P.J. Development of capsular polysaccharide-based glycoconjugates for immunization against melioidosis and glanders. Front. Cell. Infect. Microbiol. 2012, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.R., Jr.; Goldberg, J.B. Purification and visualization of lipopolysaccharide from Gram-negative bacteria by hot aqueous-phenol extraction. J. Vis. Exp. JoVE 2012, 63, 3916. [Google Scholar]
- Schlottmann, S.A.; Jain, N.; Chirmule, N.; Esser, M.T. A novel chemistry for conjugating pneumococcal polysaccharides to Luminex microspheres. J. Immunol. Methods 2006, 309, 75–85. [Google Scholar] [CrossRef]
- Perraut, R.; Richard, V.; Varela, M.L.; Trape, J.F.; Guillotte, M.; Tall, A.; Toure, A.; Sokhna, C.; Vigan-Womas, I.; Mercereau-Puijalon, O. Comparative analysis of IgG responses to Plasmodium falciparum MSP1p19 and PF13-DBL1α1 using ELISA and a magnetic bead-based duplex assay (MAGPIX®-Luminex) in a Senegalese meso-endemic community. Malar. J. 2014, 13, 410. [Google Scholar] [CrossRef] [Green Version]
- Sim, B.M.; Chantratita, N.; Ooi, W.F.; Nandi, T.; Tewhey, R.; Wuthiekanun, V.; Thaipadungpanit, J.; Tumapa, S.; Ariyaratne, P.; Sung, W.K.; et al. Genomic acquisition of a capsular polysaccharide virulence cluster by nonpathogenic. Burkholderia isolates. Genome Biol. 2010, 11, 89. [Google Scholar] [CrossRef] [PubMed]
- Sarovich, D.S.; Price, E.P.; Webb, J.R.; Ward, L.M.; Voutsinos, M.Y.; Tuanyok, A.; Currie, B.J. Variable virulence factors in Burkholderia pseudomallei (Melioidosis) associated with human disease. PLoS ONE 2014, 9, 1–4. [Google Scholar] [CrossRef]
- Panomket, P. Immune response to Burkholderia pseudomallei. J. Med. Assoc. Thail. 2011, 94, 1410–1417. [Google Scholar]
- Liu, B.; Koo, G.C.; Yap, E.H.; Chua, K.L.; Gan, Y.-H. Model of differential susceptibility to mucosal Burkholderia pseudomallei Infection. Infect. Immun. 2002, 70, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Leitão, J.H.; Sousa, S.A.; Ferreira, A.S.; Ramos, C.G.; Silva, I.N.; Moreira, L.M. Pathogenicity, virulence factors, and strategies to fight against Burkholderia cepacia complex pathogens and related species. Appl. Microbiol. Biotechnol. 2010, 87, 31–40. [Google Scholar] [CrossRef]
- Drevinek, P.; Mahenthiralingam, E. Burkholderia cenocepacia in cystic fibrosis: Epidemiology and molecular mechanisms of virulence. Clin. Microbiol. Infect. 2010, 16, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Conway, B.-A.D.; Chu, K.K.; Bylund, J.; Altman, E.; Speert, D.P. Production of exopolysaccharide by Burkholderia cenocepacia results in altered cell-surface interactions and altered bacterial clearance in mice. J. Infect. Dis. 2004, 190, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.M.; Busch, J.D.; Shippy, K.; Allender, C.J.; Kaestli, M.; Mayo, M.; Sahl, J.W.; Schupp, J.M.; Colman, R.E.; Keim, P.; et al. Diverse Burkholderia species isolated from Soils in the Southern United States with no Evidence of B. pseudomallei. PLoS ONE 2015, 10, e0143254. [Google Scholar] [CrossRef]
- Nelson, M.; Prior, J.L.; Lever, M.S.; Jones, H.E.; Atkins, T.P.; Titball, R.W. Evaluation of lipopolysaccharide and capsular polysaccharide as subunit vaccines against experimental melioidosis. J. Med. Microbiol. 2004, 53, 1177–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houghton, R.L.; Reed, D.E.; Hubbard, M.A.; Dillon, M.J.; Chen, H.; Currie, B.J.; Mayo, M.; Sarovich, D.S.; Theobald, V.; Limmathurotsakul, D.; et al. Development of a prototype lateral flow immunoassay (LFI) for the rapid diagnosis of melioidosis. PLoS Negl. Trop. Dis. 2014, 8, e2727. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.E.; Burtnick, M.N.; Stokes, M.G.; Whelan, A.O.; Williamson, E.D.; Atkins, T.P.; Prior, J.L.; Brett, P.J. Burkholderia pseudomallei capsular polysaccharide conjugates provide protection against acute melioidosis. Infect. Immun. 2014, 82, 3206–3213. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Gregory, A.E.; Hatcher, C.L.; Vinet-Oliphant, H.; Morici, L.A.; Titball, R.W.; Roy, C.J. Protection of non-human primates against glanders with a gold nanoparticle glycoconjugate vaccine. Vaccine 2015, 33, 686–692. [Google Scholar] [CrossRef]
- Suttisunhakul, V.; Wuthiekanun, V.; Brett, P.J.; Khusmith, S.; Day, N.P.; Burtnick, M.N.; Limmathurotsakul, D.; Chantratita, N. Development of rapid Enzyme-Linked Immunosorbent Assays for detection of antibodies to Burkholderia pseudomallei. J. Clin. Microbiol. 2016, 54, 1259–1268. [Google Scholar] [CrossRef]
- Pasala, S.; Barr, T.; Messaoudi, I. Impact of alcohol abuse on the adaptive immune system. Alcohol Res. Curr. Rev. 2015, 37, 185–197. [Google Scholar]
- Dillon, M.J.; Loban, R.A.; Reed, D.E.; Thorkildson, P.; Pflughoeft, K.J.; Pandit, S.G.; Brett, P.J.; Burtnick, M.N.; AuCoin, D.P. Contribution of murine IgG Fc regions to antibody binding to the capsule of Burkholderia pseudomallei. Virulence 2016, 7, 691–701. [Google Scholar] [CrossRef] [Green Version]
- Aldo-Benson, M.; Pratt, L.; Hardwick, J. Alcohol can inhibit effect of IL-4 on activated murine B cells. Immun. Res. 1992, 11, 117–124. [Google Scholar] [CrossRef]
- Stavnezer, J.; Schrader, C.E. Ig heavy chain class switch recombination: Mechanism and regulation. J. Immun. 2015, 193, 5370–5378. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, R.M.; Jimenez, V., Jr; Monroy, F.P. Impact of Binge Alcohol Intoxication on the Humoral Immune Response during Burkholderia spp. Infections. Microorganisms 2019, 7, 125. https://doi.org/10.3390/microorganisms7050125
Moreno RM, Jimenez V Jr, Monroy FP. Impact of Binge Alcohol Intoxication on the Humoral Immune Response during Burkholderia spp. Infections. Microorganisms. 2019; 7(5):125. https://doi.org/10.3390/microorganisms7050125
Chicago/Turabian StyleMoreno, Ryan M., Victor Jimenez, Jr, and Fernando P. Monroy. 2019. "Impact of Binge Alcohol Intoxication on the Humoral Immune Response during Burkholderia spp. Infections" Microorganisms 7, no. 5: 125. https://doi.org/10.3390/microorganisms7050125
APA StyleMoreno, R. M., Jimenez, V., Jr, & Monroy, F. P. (2019). Impact of Binge Alcohol Intoxication on the Humoral Immune Response during Burkholderia spp. Infections. Microorganisms, 7(5), 125. https://doi.org/10.3390/microorganisms7050125