Effects of Chestnut Tannin Extract, Vescalagin and Gallic Acid on the Dimethyl Acetals Profile and Microbial Community Composition in Rumen Liquor: An In Vitro Study

 ,
,  , ,
, , 
 ,
, 
Abstract
1. Introduction
2. Material and Method
2.1. Diets
2.2. Proximate Analyses of Feed Samples
2.3. Rumen Inoculum
2.4. DMA Analysis
2.5. Rumen Degradability
2.6. Quantification of the 16S rRNA Gene by Quantitative PCR
2.7. Amplification of 16S rRNA Gene, Sequencing and Sequence Analysis
2.8. Statistical Analysis
3. Results
3.1. Rumen Degradability and DMA Profile
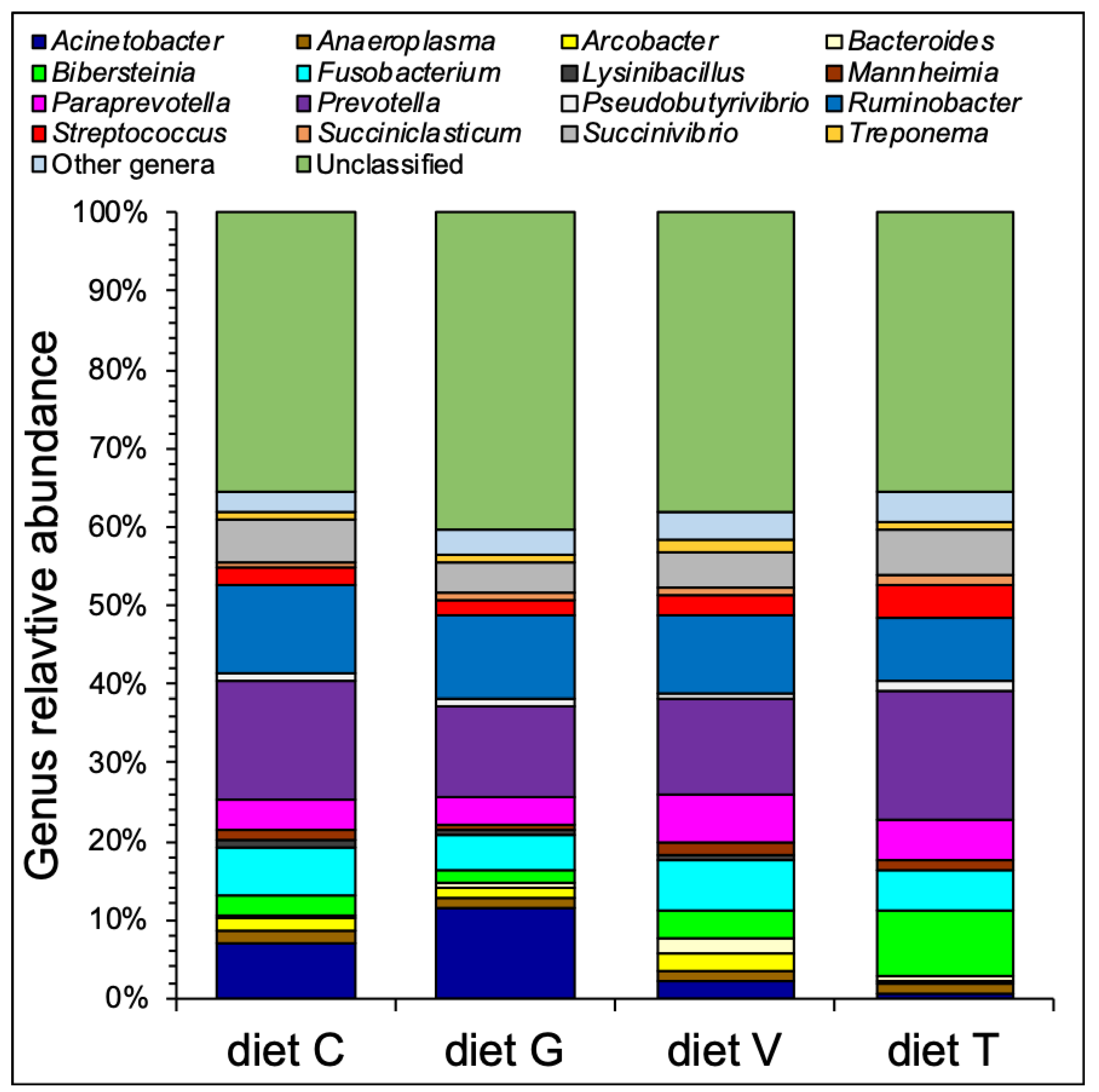
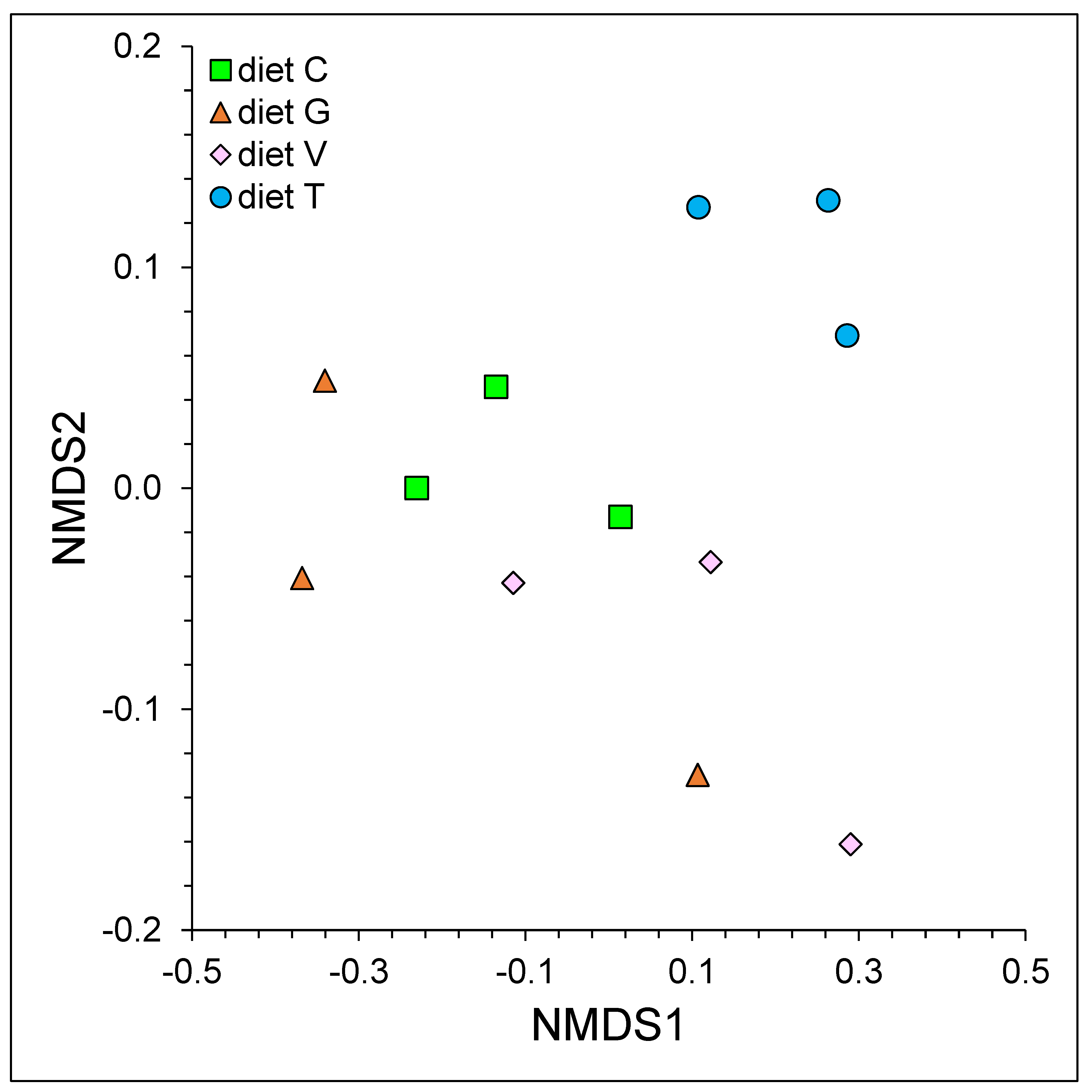
3.2. Microbial Community Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADF | acid detergent fibre |
| ADL | acid detergent lignin |
| ASV | amplicon sequence variant |
| C | control |
| CHT | chestnut tannins |
| CP | crude protein |
| DM | dry matter |
| DMA | dimethyl acetals |
| DMI | DM intake |
| EE | ether extract |
| GAL | gallic acid |
| HTS | high-throughput sequencing |
| NDF | neutral detergent fibre |
| NDFdeg | NDF degradability |
| NDFfeed | NDF of feeds |
| NDFundeg | NDF residuals |
| OM | organic matter |
| qPCR | quantitative PCR |
| RL | rumen liquor |
| VES | vescalagin |
References
- Pulina, G.; Francesconi, A.H.D.; Stefanon, B.; Sevi, A.; Calamari, L.; Lacetera, N.; Dell’Orto, V.; Pilla, F.; Marsan, P.M.; Mele, M.; et al. Sustainable ruminant production to help feed the planet. Ital. J. Anim. Sci. 2017, 16, 140–171. [Google Scholar] [CrossRef]
- Buccioni, A.; Cappucci, A.; Mele, M. Climate Change Impact on Livestock: Adaptation and Mitigation; Cap. Methane Emission from Enteric Fermentation: Methanogenesis and Fermentation; Springer: New Delhi, India, 2015; pp. 171–186. ISBN 9788132222651. [Google Scholar]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef] [PubMed]
- Oskoueian, E.; Abdullah, N.; Oskoueian, A. Effects of flavonoids on rumen fermentation activity, methane production, and microbial population. Biomed Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- Seradj, A.R.; Abecia, L.; Crespo, J.; Villalba, D.; Fondevila, M.; Balcells, J. The effect of Bioflavex® and its pure flavonoid components on in vitro fermentation parameters and methane production in rumen fluid from steers given high concentrate diets. Anim. Feed Sci. Technol. 2014, 197, 85–91. [Google Scholar] [CrossRef]
- Moate, P.J.; Williams, S.R.O.; Torok, V.A.; Hannah, M.C.; Ribaux, B.E.; Tavendale, M.H.; Eckard, R.J.; Jacobs, J.L.; Auldist, M.J.; Wales, W.J. Grape marc reduces methane emissions when fed to dairy cows. J. Dairy Sci. 2014, 97, 5073–5087. [Google Scholar] [CrossRef] [PubMed]
- Jayanegara, A.; Goel, G.; Makkar, H.P.S.; Becker, K. Divergence between purified hydrolysable and condensed tannin effects on methane emission, rumen fermentation and microbial population in vitro. Anim. Feed Sci. Technol. 2015, 209, 60–68. [Google Scholar] [CrossRef]
- Liu, H.; Vaddella, V.; Zhou, D. Effects of chestnut tannins and coconut oil on growth performance, methane emission, ruminal fermentation, and microbial populations in sheep. J. Dairy Sci. 2011, 94, 6069–6077. [Google Scholar] [CrossRef]
- Buccioni, A.; Serra, A.; Minieri, S.; Mannelli, F.; Cappucci, A.; Benvenuti, D.; Rapaccini, S.; Conte, G.; Mele, M. Milk production, composition, and milk fatty acid profile from grazing sheep fed diets supplemented with chestnut tannin extract and extruded linseed. Small Rumin. Res. 2015, 130, 200–207. [Google Scholar] [CrossRef]
- Bargiacchi, E.; Campo, M.; Romani, A.; Milli, G.; Miele, S. Hydrolysable tannins from sweet chestnut (Castanea sativa Mill.) to improve tobacco and food/feed quality. Note 1: Fraction characterization, and tobacco biostimulant effect for gall-nematode resistance. AIMS Agric. Food 2017, 2, 324–338. [Google Scholar] [CrossRef]
- Buccioni, A.; Pauselli, M.; Viti, C.; Minieri, S.; Pallara, G.; Roscini, V.; Rapaccini, S.; Marinucci, M.T.; Lupi, P.; Conte, G.; et al. Milk fatty acid composition, rumen microbial population, and animal performances in response to diets rich in linoleic acid supplemented with chestnut or quebracho tannins in dairy ewes. J. Dairy Sci. 2015, 98, 1145–1156. [Google Scholar] [CrossRef]
- Buccioni, A.; Pallara, G.; Pastorelli, R.; Bellini, L.; Cappucci, A.; Mannelli, F.; Minieri, S.; Roscini, V.; Rapaccini, S.; Mele, M.; et al. Effect of dietary chestnut or quebracho tannin supplementation on microbial community and fatty acid profile in the rumen of dairy ewes. Biomed Res. Int. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Quideau, S.; Varadinova, T.; Karagiozova, D.; Jourdes, M.; Pardon, P.; Baudry, C.; Genova, P.; Diakov, T.; Petrova, R. Main structural and stereochemical aspects of the antiherpetic activity of nonahydroxyterphenoyl-containing C-glycosidic ellagitannins. Chem. Biodivers. 2004, 1, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Murdiati, T.B.; Mcsweeney, C.S.; Lowry, J.B. Metabolism in sheep of gallic acid, tannic acid and hydrolyzable tannin from Terminalia oblongata. Aust. J. Agric. Res. 1992, 43, 1307–1319. [Google Scholar] [CrossRef]
- Ekambaram, S.P.; Perumal, S.S.; Balakrishnan, A. Scope of hydrolysable tannins as possible antimicrobial agent. Phyther. Res. 2016, 30, 1035–1045. [Google Scholar] [CrossRef]
- Panizzi, L.; Caponi, C.; Catalano, S.; Cioni, P.L.; Morelli, I. In vitro antimicrobial activity of extracts and isolated constituents of Rubus ulmifolius. J. Ethnopharmacol. 2002, 79, 165–168. [Google Scholar] [CrossRef]
- Buccioni, A.; Pauselli, M.; Minieri, S.; Roscini, V.; Mannelli, F.; Rapaccini, S.; Lupi, P.; Conte, G.; Serra, A.; Cappucci, A.; et al. Chestnut or quebracho tannins in the diet of grazing ewes supplemented with soybean oil: Effects on animal performances, blood parameters and fatty acid composition of plasma and milk lipids. Small Rumin. Res. 2017, 153, 23–30. [Google Scholar] [CrossRef]
- Burns, R.E. Methods of tannin analysis for forage crop evaluation. Ga. Agric. Exp. Stn. Tech. Bull. 1963, 32, 4–14. [Google Scholar]
- AOAC International. Official Methods of Analysis of AOAC International, 16th ed.; The Association: Washington, DC, USA, 1995; ISBN 9780935584547. [Google Scholar]
- van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Denek, N.; Can, A.; Koncagul, S. Usage of slaughtered animal rumen fluid for dry matter digestibility of ruminant feeds. J. Anim. Vet. Adv. 2006, 5, 459–461. [Google Scholar]
- Lutakome, P.; Kabi, F.; Tibayungwa, F.; Laswai, G.H.; Kimambo, A.; Ebong, C. Rumen liquor from slaughtered cattle as inoculum for feed evaluation. Anim. Nutr. 2017, 3, 300–308. [Google Scholar] [CrossRef]
- Buccioni, A.; Minieri, S.; Rapaccini, S.; Antongiovanni, M.; Mele, M. Effect of chestnut and quebracho tannins on fatty acid profile in rumen liquid- and solid-associated bacteria: An in vitro study. Animal 2011, 5, 1521–1530. [Google Scholar] [CrossRef] [PubMed]
- McDougall, E.I. Studies on Ruminant Saliva 1. The composition and output of sheep’s saliva. Biochem. J. 1947, 43, 99–109. [Google Scholar] [CrossRef]
- Alves, S.P.; Santos-Silva, J.; Cabrita, A.R.J.; Fonseca, A.J.M.; Bessa, R.J.B. Detailed dimethylacetal and fatty acid composition of rumen content from lambs fed lucerne or concentrate supplemented with soybean oil. PLoS ONE 2013, 8, e58386. [Google Scholar] [CrossRef] [PubMed]
- Tilley, J.M.A.; Terry, R.A. A two-stage technique for the in vitro digestion of forage crops. J. Br. Grassl. Soc. 1980, 18, 104–111. [Google Scholar] [CrossRef]
- Maeda, H.; Fujimoto, C.; Haruki, Y.; Maeda, T. Quantitative real-time PCR using TaqMan and SYBR Green for Actinobacillus actinomycetemcomitans, Porphyromonas gingivalis, Prevotella intermedia, tetQ gene and total bacteria. FEMS Immunol. Med. Microbiol. 2003, 39, 81–86. [Google Scholar] [CrossRef]
- Mannelli, F.; Cappucci, A.; Pini, F.; Pastorelli, R.; Decorosi, F.; Giovannetti, L.; Mele, M.; Minieri, S.; Conte, G.; Pauselli, M.; et al. Effect of different types of olive oil pomace dietary supplementation on the rumen microbial community profile in Comisana ewes. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- O’Callaghan, K.A.M.; Kerry, J.P. Preparation of low- and medium-molecular weight chitosan nanoparticles and their antimicrobial evaluation against a panel of microorganisms, including cheese-derived cultures. Food Control 2016, 69, 256–261. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef]
- Oksanen, A.J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; Hara, R.B.O.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package R Package Version 2.5-4. Available online: https://CRAN.R-project.org/package=vegan (accessed on 9 July 2019).
- SAS Institute User’s Guide: Statistics; SAS ® Documentation: Khlong Toei, Thailand, 2008.
- Vasta, V.; Luciano, G. The effects of dietary consumption of plants secondary compounds on small ruminants’ products quality. Small Rumin. Res. 2011, 101, 150–159. [Google Scholar] [CrossRef]
- Zimmer, N.; Cordesse, R. Digestibility and ruminal digestion of non-nitrogenous compounds in adult sheep and goats: Effects of chestnut tannins. Anim. Feed Sci. Technol. 1996, 61, 259–273. [Google Scholar] [CrossRef]
- Tabacco, E.; Borreani, G.; Crovetto, G.M.; Galassi, G.; Colombo, D.; Cavallarin, L. Effect of chestnut tannin on fermentation quality, proteolysis, and protein rumen degradability of alfalfa silage. J. Dairy Sci. 2010, 89, 4736–4746. [Google Scholar] [CrossRef]
- Smith, A.H.; Zoetendal, E.; Mackie, R.I. Bacterial mechanisms to overcome inhibitory effects of dietary tannins. Microb. Ecol. 2005, 50, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Alves, S.P.; Cappucci, A.; Cook, S.R.; Duarte, A.; Caldeira, R.M.; McAllister, T.A.; Bessa, R.J.B. Effects of condensed and hydrolyzable tannins on rumen metabolism with emphasis on the biohydrogenation of unsaturated fatty acids. J. Agric. Food Chem. 2018, 66, 3367–3377. [Google Scholar] [CrossRef]
- Deaville, E.R.; Givens, D.I.; Mueller-Harvey, I. Chestnut and mimosa tannin silages: Effects in sheep differ for apparent digestibility, nitrogen utilisation and losses. Anim. Feed Sci. Technol. 2010, 157, 129–138. [Google Scholar] [CrossRef]
- McSweeney, C.S.; Palmer, B.; Bunch, R.; Krause, D.O. Isolation and characterization of proteolytic ruminal bacteria from sheep and goats fed the tannin-containing shrub legume Calliandra calothyrsus. Appl. Environ. Microbiol. 1999, 65, 3075–3083. [Google Scholar] [PubMed]
- Reed, J.D. Relationships among soluble phenolics, insoluble proanthocyanidins and fiber in East African browse species. J. Range Manag. 1986, 39, 5–7. [Google Scholar] [CrossRef]
- Reed, J.D. Nutritional toxicology of tannins and related polyphenols in forage legumes. J. Anim. Sci. 1995, 73, 1516–1528. [Google Scholar] [CrossRef]
- Li, F.; Henderson, G.; Sun, X.; Cox, F.; Janssen, P.H.; Guan, L.L. Taxonomic assessment of rumen microbiota using total RNA and targeted amplicon sequencing approaches. Front. Microbiol. 2016, 7, 987. [Google Scholar] [CrossRef]
- Kaneda, T. Iso- and anteiso-fatty acids in bacteria: Biosynthesis, function, and taxonomic significance. Microbiol Mol. Biol. Rev. 1991, 55, 288–302. [Google Scholar]
- Goldfine, H. The appearance, disappearance and reappearance of plasmalogens in evolution. Prog. Lipid Res. 2010, 49, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Collaborators, G.R.C.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Baldwin, R.L.; Li, W.; Li, C.; Connor, E.E.; Li, R.W. The bacterial community composition of the bovine rumen detected using pyrosequencing of 16S rRNA genes. Metagenomics 2012, 1, 1–11. [Google Scholar] [CrossRef]
- Seshadri, R.; Leahy, S.C.; Attwood, G.T.; Teh, K.H.; Lambie, S.C.; Cookson, A.L.; Eloe-Fadrosh, E.A.; Pavlopoulos, G.A.; Hadjithomas, M.; Varghese, N.J.; et al. Cultivation and sequencing of rumen microbiome members from the Hungate1000 Collection. Nat. Biotechnol. 2018, 36, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2017, 45, D37–D42. [Google Scholar] [CrossRef] [PubMed]
- Cantalapiedra-Hijar, G.; Abo-Ismail, M.; Carstens, G.E.; Guan, L.L.; Hegarty, R.; Kenny, D.A.; McGee, M.; Plastow, G.; Relling, A.; Ortigues-Marty, I. Review: Biological determinants of between-animal variation in feed efficiency of growing beef cattle. Animal 2018, 12, s321–s335. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| diet C | diet T | diet G | diet V | SEM 1 | p2 | |
|---|---|---|---|---|---|---|
| NDF g/100g DM | 41.73 b | 41.88 b | 44.79 a | 43.71 a | 0.75 | 0.0380 * |
| Degradability g/100g DM | 10.49 a | 10.17 a | 3.94 c | 6.25 b | 1.61 | 0.0380 * |
| DMA | diet C | diet T | diet G | diet V | SEM 1 | p2 |
|---|---|---|---|---|---|---|
| DMA 12:0 | 0.052 b | 0.257 a | 0.266 a | 0.234 a | 0.044 | 0.0294 * |
| DMA 13:0 iso | 0.025 | 0.124 | 0.103 | 0.095 | 0.024 | 0.0868 |
| DMA 13:0 | 0.044 b | 0.128 a | 0.124 a | 0.132 a | 0.020 | 0.0458 * |
| DMA 14:0 iso | 0.365 b | 0.244 b | 1.318 a | 1.132 a | 0.152 | 0.0075 * |
| DMA 14:0 | 0.281 b | 0.824 a | 0.921 a | 0.800 a | 0.122 | 0.0238 * |
| DMA 15:0 iso | 0.257 b | 0.652 a | 0.816 a | 0.700 a | 0.110 | 0.0329 * |
| DMA 15:0 | 0.280 | 0.593 | 0.477 | 0.530 | 0.223 | 0.7784 |
| DMA 16:0 | 1.957 | 5.225 | 5.628 | 5.454 | 0.956 | 0.0767 |
| DMA 16:1 | 0.074 c | 0.316 a | 0.212 b | 0.190 b | 0.035 | 0.0088 * |
| DMA 17:0 iso | 0.041 | 0.122 | 0.145 | 0.144 | 0.029 | 0.1061 |
| DMA 17:0 ante | 0.054 c | 0.212 a | 0.214 a | 0.168 b | 0.024 | 0.0054 * |
| DMA 17:0 | 0.030 d | 0.181 a | 0.114 b | 0.083 c | 0.025 | 0.0200 * |
| DMA 18:0 | 0.186 b | 0.518 a | 0.572 a | 0.567 a | 0.093 | 0.0544 |
| DMA 18:1 trans-11 | 0.030 b | 0.122 a | 0.087 a | 0.094 a | 0.015 | 0.0208 * |
| DMA 18:1 cis-9 | 0.173 b | 0.571 a | 0.408 b | 0.417 a,b | 0.077 | 0.0389 * |
| DMA 18:1 cis-11 | 0.087 | 0.294 | 0.166 | 0.227 | 0.048 | 0.0751 |
| DMA 18:1 cis-12 | 0.024 | 0.081 | 0.026 | 0.040 | 0.020 | 0.2493 |
| DMA 17:1 | 0.097 | 0.360 | 0.276 | 0.247 | 0.098 | 0.3558 |
| DMA 18:2 | 0.099 | 0.904 | 0.250 | 0.185 | 0.284 | 0.2489 |
| DMA 26:0 | 0.552 | 1.3700 | 0.961 | 1.3570 | 0.290 | 0.2283 |
| diet C | diet T | diet G | diet V | |
|---|---|---|---|---|
| ASVs | 156 ± 3 | 154 ± 12 | 147 ± 16 | 148 ± 2 |
| Shannon Index | 4.30 ± 0.03 | 4.3 ± 0.1 | 4.2 ± 0.1 | 4.29 ± 0.06 |
| Phylum | Class | Order | Family | Genus |
|---|---|---|---|---|
| Bacteroidetes | Bacilli | Aeromonadales | Acidaminococcaceae | Acinetobacter |
| Candidatus_Saccharibacteria | Bacteroidia | Anaeroplasmatales | Anaeroplasmataceae | Alloprevotella |
| Euryarchaeota | Betaproteobacteria | Bacteroidales | Bacteroidaceae | Anaeroplasma |
| Firmicutes | Clostridia | Burkholderiales | Campylobacteraceae | Anaerovibrio |
| Fusobacteria | Deltaproteobacteria | Campylobacterales | Clostridiales_Incertae_Sedis_XIII | Arcobacter |
| Proteobacteria | Epsilonproteobacteria | Clostridiales | Desulfovibrionaceae | Bacteroides |
| Spirochaetes | Erysipelotrichia | Desulfovibrionales | Erysipelotrichaceae | Bibersteinia |
| SR1 | Fusobacteriia | Erysipelotrichales | Fusobacteriaceae | Campylobacter |
| Tenericutes | Gammaproteobacteria | Fusobacteriales | Lachnospiraceae | Desulfovibrio |
| Verrucomicrobia | Methanobacteria | Lactobacillales | Methanobacteriaceae | Fusobacterium |
| Mollicutes | Methanobacteriales | Methanomassiliicoccaceae | Mannheimia | |
| Negativicutes | Methanomassiliicoccales | Moraxellaceae | Methanobrevibacter | |
| Spirochaetia | Neisseriales | Neisseriaceae | Methanomassiliicoccus | |
| Subdivision5 | Pasteurellales | Pasteurellaceae | Moraxella | |
| Thermoplasmata | Pseudomonadales | Porphyromonadaceae | Paraprevotella | |
| Selenomonadales | Prevotellaceae | Prevotella | ||
| Spirochaetales | Ruminococcaceae | Pseudobutyrivibrio | ||
| Spirochaetaceae | Roseburia | |||
| Streptococcaceae | Ruminobacter | |||
| Succinivibrionaceae | Saccharofermentans | |||
| Veillonellaceae | Selenomonas | |||
| Streptococcus | ||||
| Succiniclasticum | ||||
| Succinivibrio | ||||
| Treponema |
| Domain | Phylum | Class | Order | Family | Genus | diet C (%) | diet T (%) | diet G (%) | diet V (%) | SEM 1 | p2 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Archaea | Euryarchaeota | Methanobacteria | Methanobacteriales | Methanobacteriaceae | Methanobrevibacter | 0.34 | 0.37 | 0.20 | 0.48 | 0.15 | 0.6309 |
| Methanosphaera | <0.01 | <0.01 | 0.03 | <0.01 | 0.01 | 0.4411 | |||||
| Thermoplasmata | Methanomassiliicoccales | Methanomassiliicoccaceae | Methanomassiliicoccus | 0.19 | 0.29 | 0.54 | 0.48 | 0.13 | 0.2790 | ||
| Bacteria | Bacteroidetes | Bacteroidia | Bacteroidales | Bacteroidaceae | Bacteroides | 0.28 | 0.57 | 0.38 | 1.96 | 0.48 | 0.1218 |
| Porphyromonadaceae | Petrimonas | 0.03 | N.D. 3 | N.D. | 0.04 | 0.03 | 0.5880 | ||||
| Porphyromonas | <0.01 | <0.01 | <0.01 | 0.02 | 0.01 | 0.4411 | |||||
| Prevotellaceae | Alloprevotella | 0.04 | 0.05 | 0.07 | 0.09 | 0.03 | 0.6864 | ||||
| Paraprevotella | 3.81 | 5.25 | 3.67 | 6.16 | 0.91 | 0.2399 | |||||
| Prevotella | 15.15 | 16.16 | 11.46 | 11.97 | 1.53 | 0.1528 | |||||
| Elusimicrobia | Elusimicrobia | Elusimicrobiales | Elusimicrobiaceae | Elusimicrobium | 0.01 | N.D. | 0.01 | N.D. | 0.01 | 0.5948 | |
| Fibrobacteres | Fibrobacteria | Fibrobacterales | Fibrobacteraceae | Fibrobacter | 0.02 | 0.01 | 0.03 | <0.01 | 0.02 | 0.7279 | |
| Firmicutes | Bacilli | Bacillales | Planococcaceae | Kurthia | N.D. | N.D. | 0.18 | N.D. | 0.09 | 0.4411 | |
| Lysinibacillus | 1.16 | N.D. | 0.69 | 0.67 | 0.39 | 0.2896 | |||||
| Lactobacillales | Streptococcaceae | Streptococcus | 2.19 b | 4.22 a | 1.99 b | 2.51 b | 0.24 | 0.0006 * | |||
| Clostridia | Clostridiales | Lachnospiraceae | Butyrivibrio | <0.01 | 0.17 | <0.01 | <0.01 | 0.09 | 0.4411 | ||
| Clostridium_XlVa | N.D. | 0.13 | 0.09 | 0.12 | 0.09 | 0.7078 | |||||
| Clostridium_XlVb | N.D. | N.D. | N.D. | 0.03 | 0.01 | 0.4411 | |||||
| Pseudobutyrivibrio | 0.71 b | 1.31 a | 0.99 a,b | 0.68 b | 0.10 | 0.0086 * | |||||
| Roseburia | 0.21 | 0.14 | 0.22 | 0.22 | 0.10 | 0.9232 | |||||
| Peptostreptococcaceae | Peptostreptococcus | <0.01 | <0.01 | <0.01 | 0.02 | 0.01 | 0.4411 | ||||
| Ruminococcaceae | Anaerofilum | N.D. | N.D. | N.D. | 0.03 | 0.01 | 0.4411 | ||||
| Clostridium_IV | 0.06 | 0.02 | N.D. | 0.06 | 0.03 | 0.5830 | |||||
| Ruminococcus | 0.14 | 0.03 | 0.10 | <0.01 | 0.06 | 0.4272 | |||||
| Saccharofermentans | 0.04 | 0.11 | 0.11 | 0.06 | 0.06 | 0.7622 | |||||
| Negativicutes | Selenomonadales | Acidaminococcaceae | Succiniclasticum | 0.78 | 1.15 | 0.8 | 1.06 | 0.18 | 0.4620 | ||
| Veillonellaceae | Anaerovibrio | 0.12 b | 0.53 a | 0.11 b | 0.15 b | 0.09 | 0.0317 * | ||||
| Selenomonas | 0.58 | 0.94 | 0.47 | 0.56 | 0.12 | 0.1015 | |||||
| Fusobacteria | Fusobacteriia | Fusobacteriales | Fusobacteriaceae | Fusobacterium | 6.07 | 5.28 | 4.20 | 6.56 | 0.69 | 0.1647 | |
| Proteobacteria | Betaproteobacteria | Burkholderiales | Comamonadaceae | Brachymonas | N.D. | 0.03 | N.D. | N.D. | 0.01 | 0.4411 | |
| Comamonas | N.D. | 0.08 | N.D. | N.D. | 0.04 | 0.4411 | |||||
| Deltaproteobacteria | Desulfovibrionales | Desulfovibrionaceae | Desulfovibrio | 0.05 | 0.02 | 0.05 | 0.03 | 0.04 | 0.8945 | ||
| Epsilonproteobacteria | Campylobacterales | Campylobacteraceae | Arcobacter | 1.84 a | 0.29 b | 1.39 a | 2.23 a | 0.25 | 0.0028 * | ||
| Campylobacter | 0.42 | 0.24 | 0.50 | 0.51 | 0.12 | 0.3658 | |||||
| Gammaproteobacteria | Aeromonadales | Succinivibrionaceae | Ruminobacter | 11.28 | 8.12 | 10.59 | 9.91 | 1.36 | 0.4438 | ||
| Succinivibrio | 5.41 | 5.76 | 4.00 | 4.59 | 0.51 | 0.1443 | |||||
| Enterobacteriales | Enterobacteriaceae | Escherichia/Shigella | 0.02 b | 0.24 a | N.D. b | 0.07 b | 0.03 | 0.0009 * | |||
| Pasteurellales | Pasteurellaceae | Actinobacillus | 0.06 | 0.14 | N.D. | <0.01 | 0.04 | 0.1154 | |||
| Bibersteinia | 2.39 b | 8.30 a | 1.86 b | 3.49 b | 0.87 | 0.0030 * | |||||
| Mannheimia | 1.33 | 1.15 | 0.72 | 1.55 | 0.41 | 0.5536 | |||||
| Pseudomonadales | Moraxellaceae | Acinetobacter | 7.08 | 0.62 | 11.49 | 2.33 | 3.36 | 0.1752 | |||
| Moraxella | 0.17 | 0.13 | 0.21 | 0.18 | 0.09 | 0.9269 | |||||
| Spirochaetes | Spirochaetia | Spirochaetales | Spirochaetaceae | Treponema | 1.04 a,b | 1.19 a,b | 0.82 b | 1.56 a | 0.17 | 0.0778 * | |
| Synergistetes | Synergistia | Synergistales | Synergistaceae | Pyramidobacter | <0.01 | 0.13 | 0.22 | 0.19 | 0.07 | 0.2345 | |
| Tenericutes | Mollicutes | Anaeroplasmatales | Anaeroplasmataceae | Anaeroplasma | 1.40 | 1.37 | 1.33 | 1.16 | 0.19 | 0.8156 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannelli, F.; Daghio, M.; Alves, S.P.; Bessa, R.J.B.; Minieri, S.; Giovannetti, L.; Conte, G.; Mele, M.; Messini, A.; Rapaccini, S.; et al. Effects of Chestnut Tannin Extract, Vescalagin and Gallic Acid on the Dimethyl Acetals Profile and Microbial Community Composition in Rumen Liquor: An In Vitro Study. Microorganisms 2019, 7, 202. https://doi.org/10.3390/microorganisms7070202
Mannelli F, Daghio M, Alves SP, Bessa RJB, Minieri S, Giovannetti L, Conte G, Mele M, Messini A, Rapaccini S, et al. Effects of Chestnut Tannin Extract, Vescalagin and Gallic Acid on the Dimethyl Acetals Profile and Microbial Community Composition in Rumen Liquor: An In Vitro Study. Microorganisms. 2019; 7(7):202. https://doi.org/10.3390/microorganisms7070202
Chicago/Turabian StyleMannelli, Federica, Matteo Daghio, Susana P. Alves, Rui J. B. Bessa, Sara Minieri, Luciana Giovannetti, Giuseppe Conte, Marcello Mele, Anna Messini, Stefano Rapaccini, and et al. 2019. "Effects of Chestnut Tannin Extract, Vescalagin and Gallic Acid on the Dimethyl Acetals Profile and Microbial Community Composition in Rumen Liquor: An In Vitro Study" Microorganisms 7, no. 7: 202. https://doi.org/10.3390/microorganisms7070202
APA StyleMannelli, F., Daghio, M., Alves, S. P., Bessa, R. J. B., Minieri, S., Giovannetti, L., Conte, G., Mele, M., Messini, A., Rapaccini, S., Viti, C., & Buccioni, A. (2019). Effects of Chestnut Tannin Extract, Vescalagin and Gallic Acid on the Dimethyl Acetals Profile and Microbial Community Composition in Rumen Liquor: An In Vitro Study. Microorganisms, 7(7), 202. https://doi.org/10.3390/microorganisms7070202