Z/I1 Hybrid Virulence Plasmids Carrying Antimicrobial Resistance genes in S. Typhimurium from Australian Food Animal Production




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Materials
2.1. Strains
2.2. DNA Extraction, Sequencing and Genome Assembly
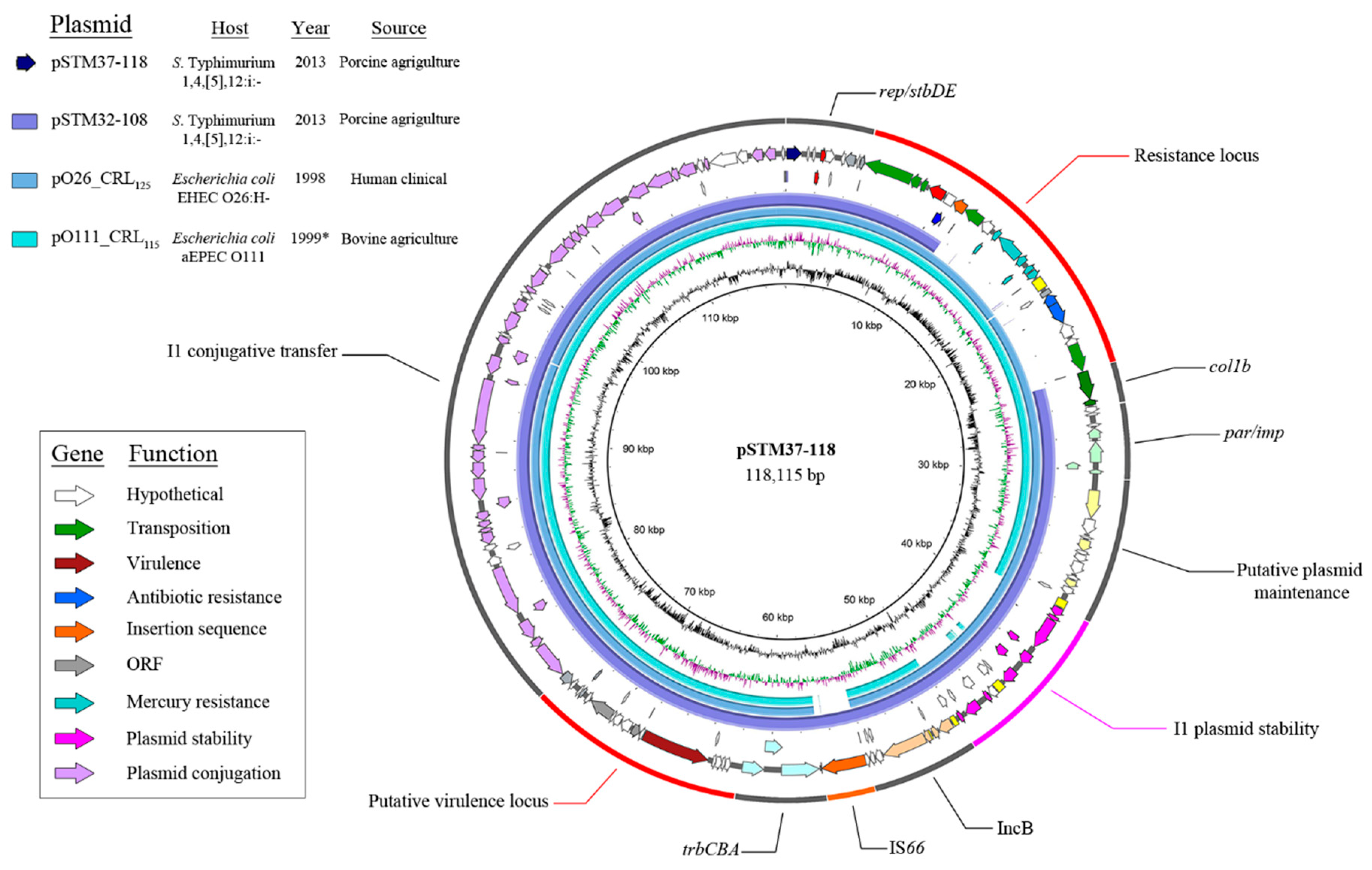
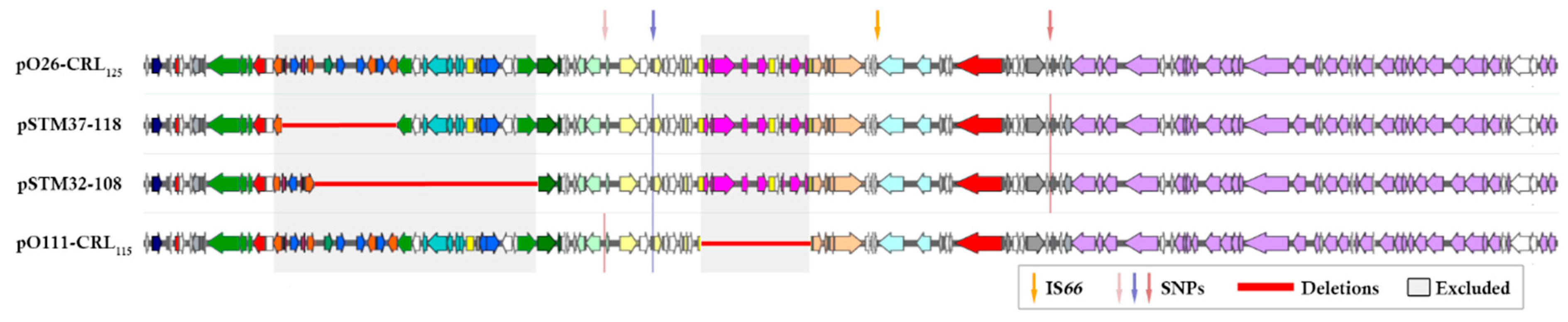
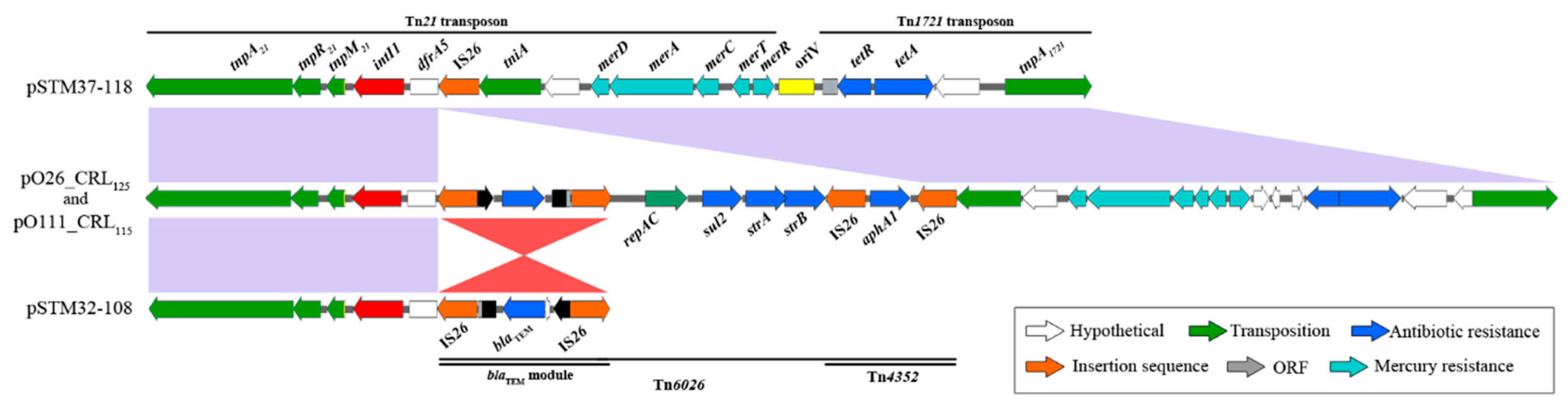
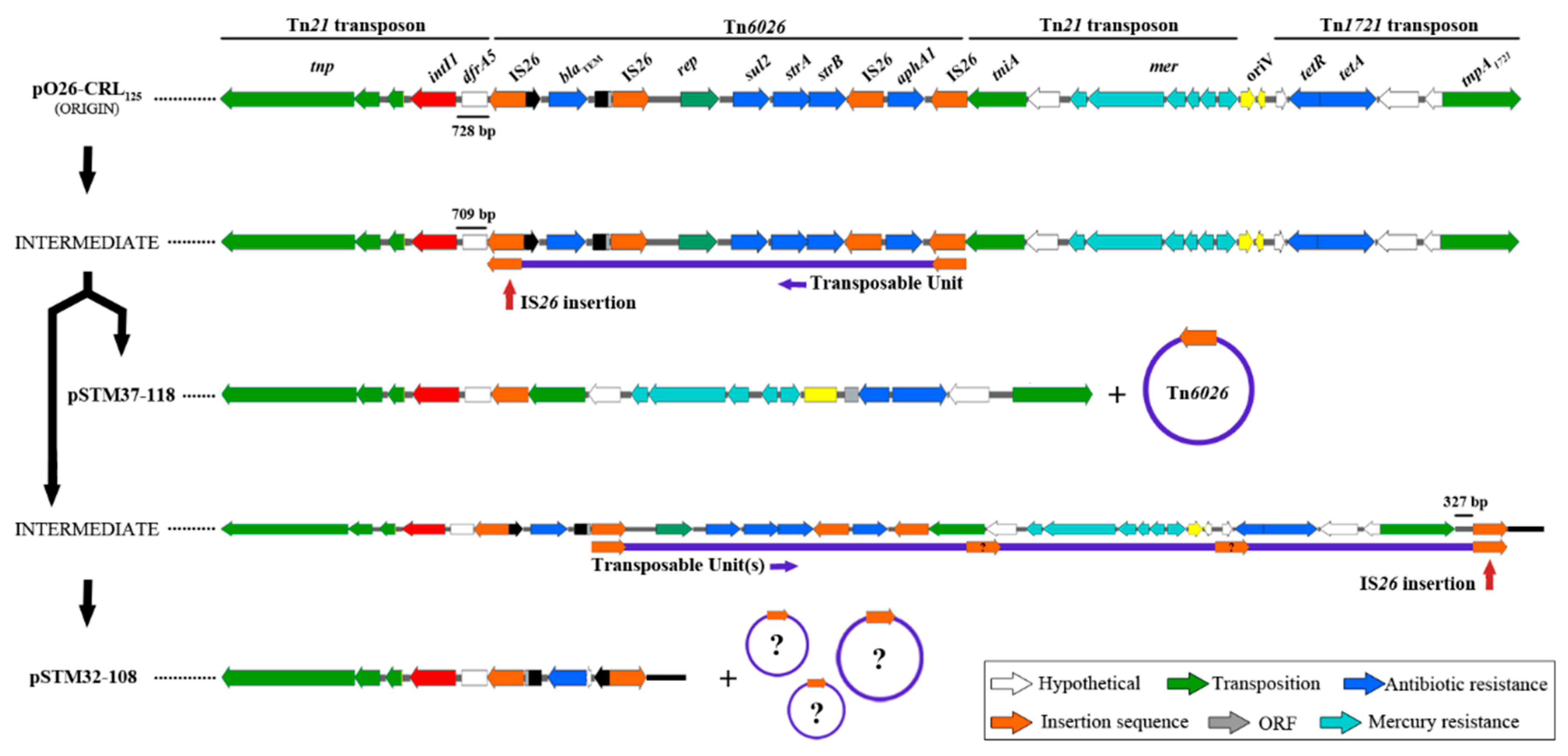
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Bannas, P.; Fraedrich, K.; Treszl, A.; Bley, T.A.; Herrmann, J.; Habermann, C.R.; Derlin, T.; Henes, F.O.; Wenzel, U.; Adam, G.; et al. Shiga Toxin-Producing E. Coli O104:H4 Outbreak 2011 in Germany: Radiological Features of Enterohemorrhagic Colitis. Rofo 2013, 185, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Pitout, J.D. Extraintestinal Pathogenic Escherichia coli: A Combination of Virulence with Antibiotic Resistance. Front. Microbiol. 2012, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Pierard, D.; De Greve, H.; Haesebrouck, F.; Mainil, J. O157:H7 and O104:H4 Vero/Shiga toxin-producing Escherichia coli outbreaks: Respective role of cattle and humans. Vet. Res. 2012, 43, 13. [Google Scholar] [CrossRef] [PubMed]
- Hardefeldt, L.; Gilkerson, J.; Billman-Jacobe, H.; Stevenson, M.; Thursky, K.; Browning, G.; Bailey, K. Antimicrobial labelling in Australia: a threat to antimicrobial stewardship? Aust. Vet. J. 2018, 96, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Bettelheim, K.A.; Hornitzky, M.A.; Djordjevic, S.P.; Kuzevski, A. Antibiotic resistance among verocytotoxigenic Escherichia coli (VTEC) and non-VTEC isolated from domestic animals and humans. J. Med. Microbiol. 2003, 52, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.; Jordan, D.; Wong, H.S.; Johnson, J.R.; Toleman, M.A.; Wakeham, D.L.; Gordon, D.M.; Turnidge, J.D.; Mollinger, J.L.; Gibson, J.S.; et al. First detection of extended-spectrum cephalosporin- and fluoroquinolone-resistant Escherichia coli in Australian food-producing animals. J. Glob. Antimicrob. Resist. 2015, 3, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Reid, C.J.; Wyrsch, E.R.; Roy Chowdhury, P.; Zingali, T.; Liu, M.; Darling, A.E.; Chapman, T.A.; Djordjevic, S.P. Porcine commensal Escherichia coli: A reservoir for class 1 integrons associated with IS26. Microb. Genom. 2017, 3, e000143. [Google Scholar] [CrossRef]
- Cain, A.K.; Liu, X.; Djordjevic, S.P.; Hall, R.M. Transposons related to Tn1696 in IncHI2 plasmids in multiply antibiotic resistant Salmonella enterica serovar Typhimurium from Australian animals. Microb. Drug Resist. 2010, 16, 197–202. [Google Scholar] [CrossRef]
- Dawes, F.E.; Kuzevski, A.; Bettelheim, K.A.; Hornitzky, M.A.; Djordjevic, S.P.; Walker, M.J. Distribution of class 1 integrons with IS26-mediated deletions in their 3’-conserved segments in Escherichia coli of human and animal origin. PLoS ONE 2010, 5, e12754. [Google Scholar] [CrossRef]
- Abraham, S.; Kirkwood, R.N.; Laird, T.; Saputra, S.; Mitchell, T.; Singh, M.; Linn, B.; Abraham, R.J.; Pang, S.; Gordon, D.M.; et al. Dissemination and persistence of extended-spectrum cephalosporin-resistance encoding IncI1-blaCTXM-1 plasmid among Escherichia coli in pigs. The ISME J. 2018, 12, 2352–2362. [Google Scholar] [CrossRef]
- Billman-Jacobe, H.; Liu, Y.; Haites, R.; Weaver, T.; Robinson, L.; Marenda, M.; Dyall-Smith, M. pSTM6-275, a conjugative IncHI2 plasmid of Salmonella that confers antibiotic and heavy metal resistance under changing physiological conditions. Antimicrob Agents Chemother. 2018, 62, e02357-17. [Google Scholar] [CrossRef] [PubMed]
- Moran, R.A.; Holt, K.E.; Hall, R.M. pCERC3 from a commensal ST95 Escherichia coli: A ColV virulence-multiresistance plasmid carrying a sul3-associated class 1 integron. Plasmid 2016, 84, 11–19. [Google Scholar] [CrossRef]
- Dyall-Smith, M.L.; Liu, Y.; Billman-Jacobe, H. Genome Sequence of an Australian Monophasic Salmonella enterica subsp. enterica Typhimurium Isolate (TW-Stm6) Carrying a Large Plasmid with Multiple Antimicrobial Resistance Genes. Genome Announc. 2017, 5, e00793-17. [Google Scholar] [CrossRef] [PubMed]
- Venturini, C.; Beatson, S.A.; Djordjevic, S.P.; Walker, M.J. Multiple antibiotic resistance gene recruitment onto the enterohemorrhagic Escherichia coli virulence plasmid. FASEB J. 2010, 24, 1160–1166. [Google Scholar] [CrossRef]
- Venturini, C.; Hassan, K.A.; Roy Chowdhury, P.; Paulsen, I.T.; Walker, M.J.; Djordjevic, S.P. Sequences of Two Related Multiple Antibiotic Resistance Virulence Plasmids Sharing a Unique IS26-Related Molecular Signature Isolated from Different Escherichia coli Pathotypes from Different Hosts. PLoS ONE 2013, 8, e78862. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.; Trott, D.J.; Jordan, D.; Gordon, D.M.; Groves, M.D.; Fairbrother, J.M.; Smith, M.G.; Zhang, R.; Chapman, T.A. Phylogenetic and molecular insights into the evolution of multidrug-resistant porcine enterotoxigenic Escherichia coli in Australia. Int. J. Antimicrob. Agents 2014, 44, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Saputra, S.; Jordan, D.; Mitchell, T.; Wong, H.S.; Abraham, R.J.; Kidsley, A.; Turnidge, J.; Trott, D.J.; Abraham, S. Antimicrobial resistance in clinical Escherichia coli isolated from companion animals in Australia. Vet. Microbiol. 2017, 211, 43–50. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, J.; Roy Chowdhury, P.; Djordjevic, S.P. Genomic analysis of multidrug-resistant Escherichia coli ST58 causing urosepsis. Int. J. Antimicrob. Agents 2018, 52, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Roy Chowdhury, P.; McKinnon, J.M.; Liu, M.Y.; Djordjevic, S.P. Multidrug resistant uropathogenic Escherichia coli ST405 with a novel, composite IS26 transposon in a unique chromosomal location. Front. Microbiol. 2018, 9, 3212. [Google Scholar] [CrossRef] [PubMed]
- Cummins, M.L.; Reid, C.J.; Roy Chowdhury, P.; Bushell, R.N.; Esbert, N.; Tivendale, K.A.; Noormohammadi, A.H.; Islam, S.; Marenda, M.S.; Browning, G.F.; et al. Whole genome sequence analysis of Australian avian pathogenic Escherichia coli that carry the class 1 integrase gene. Microb. Genom. 2019, 5, e000250. [Google Scholar] [CrossRef] [PubMed]
- OzFoodNet. OzFoodNet quarterly report, 1 January to 31 March 2015. Commun. Dis. Intell. Q. Rep. 2017, 41, E186–E193. [Google Scholar]
- Weaver, T.; Valcanis, M.; Mercoulia, K.; Sait, M.; Tuke, J.; Kiermeier, A.; Hogg, G.; Pointon, A.; Hamilton, D.; Billman-Jacobe, H. Longitudinal study of Salmonella 1,4,[5],12:i:- shedding in five Australian pig herds. Prev. Vet. Med. 2017, 136, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Hauser, E.; Tietze, E.; Helmuth, R.; Junker, E.; Blank, K.; Prager, R.; Rabsch, W.; Appel, B.; Fruth, A.; Malorny, B. Pork Contaminated with Salmonella enterica Serovar 4,[5],12:i:-, an Emerging Health Risk for Humans. Appl. Environ. Microbiol. 2010, 76, 4601–4610. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.L.; Kirchner, M.; Guerra, B.; Granier, S.A.; Lucarelli, C.; Porrero, M.C.; Jakubczak, A.; Threlfall, E.J.; Mevius, D.J. Multiresistant Salmonella enterica serovar 4,[5],12:i:- in Europe: a new pandemic strain? Eurosurveillance 2010, 15, 19580. [Google Scholar] [CrossRef] [PubMed]
- Switt, A.I.; Soyer, Y.; Warnick, L.D.; Wiedmann, M. Emergence, distribution, and molecular and phenotypic characteristics of Salmonella enterica serotype 4,5,12:i. Foodborne Pathog. Dis. 2009, 6, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, D.; Holds, G.; Hogg, G.; Valcanis, M.; Kiermeier, A. Longitudinal study of an Australian pig farm infected with monophasic Salmonella Typhimurium-like PT193 (1, 4,[5], 12: i:-PT193) using MLVA. In Proceedings of the Fourth International Symposium on the Epidemiology and Control of Hazards in Pork Production Chain, Porto, Portugal, 7–10 September 2015; pp. 127–130. [Google Scholar]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Moller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol 2014, 15, 524. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fernandez, A.; Chiaretto, G.; Bertini, A.; Villa, L.; Fortini, D.; Ricci, A.; Carattoli, A. Multilocus sequence typing of IncI1 plasmids carrying extended-spectrum beta-lactamases in Escherichia coli and Salmonella of human and animal origin. J. Antimicrob. Chemother. 2008, 61, 1229–1233. [Google Scholar] [CrossRef] [PubMed]
- Reid, C.J.; Roy Chowdhury, P.; Djordjevic, S.P. Tn6026 and Tn6029 are found in complex resistance regions mobilised by diverse plasmids and chromosomal islands in multiple antibiotic resistant Enterobacteriaceae. Plasmid 2015, 80, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Roy Chowdhury, P.; Charles, I.G.; Djordjevic, S.P. A role for Tn6029 in the evolution of the complex antibiotic resistance gene loci in genomic island 3 in enteroaggregative hemorrhagic Escherichia coli O104:H4. PLoS ONE 2015, 10, e0115781. [Google Scholar] [CrossRef] [PubMed]
- Dionisi, A.M.; Lucarelli, C.; Owczarek, S.; Luzzi, I.; Villa, L. Characterization of the plasmid-borne quinolone resistance gene qnrB19 in Salmonella enterica serovar Typhimurium. Antimicrob. Agents Chemother. 2009, 53, 4019–4021. [Google Scholar] [CrossRef] [PubMed]
- Liakopoulos, A.; Mevius, D.; Ceccarelli, D. A Review of SHV Extended-Spectrum beta-Lactamases: Neglected Yet Ubiquitous. Front Microbiol. 2016, 7, 1374. [Google Scholar] [CrossRef] [PubMed]
- Harmer, C.J.; Moran, R.A.; Hall, R.M. Movement of IS26-Associated Antibiotic Resistance Genes Occurs via a Translocatable Unit That Includes a Single IS26 and Preferentially Inserts Adjacent to Another IS26. MBio 2014, 5, e01801–e01814. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, W.; Xu, X.; Zhou, X.; Shi, C. Transmissible ST3-IncHI2 Plasmids Are Predominant Carriers of Diverse Complex IS26-Class 1 Integron Arrangements in Multidrug-Resistant Salmonella. Front Microbiol. 2018, 9, 2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangat, C.S.; Bekal, S.; Irwin, R.J.; Mulvey, M.R. A Novel Hybrid Plasmid Carrying Multiple Antimicrobial Resistance and Virulence Genes in Salmonella enterica Serovar Dublin. Antimicrob. Agents Chemother. 2017, 61, e02601–e02616. [Google Scholar] [CrossRef]
- Lee, K.Y.; Hopkins, J.D.; Syvanen, M. Direct involvement of IS26 in an antibiotic resistance operon. J. Bacteriol. 1990, 172, 3229–3236. [Google Scholar] [CrossRef] [Green Version]
- Porse, A.; Schonning, K.; Munck, C.; Sommer, M.O. Survival and Evolution of a Large Multidrug Resistance Plasmid in New Clinical Bacterial Hosts. Mol. Biol. Evol. 2016, 33, 2860–2873. [Google Scholar] [CrossRef] [PubMed]
- Enne, V.I.; Livermore, D.M.; Stephens, P.; Hall, L.M. Persistence of sulphonamide resistance in Escherichia coli in the UK despite national prescribing restriction. Lancet 2001, 357, 1325–1328. [Google Scholar] [CrossRef]
- Grape, M.; Sundstrom, L.; Kronvall, G. Sulphonamide resistance gene sul3 found in Escherichia coli isolates from human sources. J. Antimicrob. Chemother. 2003, 52, 1022–1024. [Google Scholar] [CrossRef] [PubMed]
- Hammerum, A.M.; Sandvang, D.; Andersen, S.R.; Seyfarth, A.M.; Porsbo, L.J.; Frimodt-Moller, N.; Heuer, O.E. Detection of sul1, sul2 and sul3 in sulphonamide resistant Escherichia coli isolates obtained from healthy humans, pork and pigs in Denmark. Int. J. Food Microbiol. 2006, 106, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Razavi, M.; Marathe, N.P.; Gillings, M.R.; Flach, C.F.; Kristiansson, E.; Joakim Larsson, D.G. Discovery of the fourth mobile sulfonamide resistance gene. Microbiome. 2017, 5, 160. [Google Scholar] [CrossRef] [PubMed]
- Jordan, D.; Chin, J.J.; Fahy, V.A.; Barton, M.D.; Smith, M.G.; Trott, D.J. Antimicrobial use in the Australian pig industry: results of a national survey. Aust. Vet. J. 2009, 87, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.H.; Liu, L.; Yan, M.; Chan, E.W.; Chen, S. Dissemination of IncI2 Plasmids That Harbor the blaCTX-M Element among Clinical Salmonella Isolates. Antimicrob. Agents Chemother. 2015, 59, 5026–5028. [Google Scholar] [CrossRef]
- Wang, J.; Zhi, C.P.; Chen, X.J.; Guo, Z.W.; Liu, W.L.; Luo, J.; Huang, X.Y.; Zeng, L.; Huang, J.W.; Xia, Y.B.; et al. Characterization of oqxAB in Escherichia coli Isolates from Animals, Retail Meat, and Human Patients in Guangzhou, China. Front Microbiol. 2017, 8, 1982. [Google Scholar] [CrossRef]
- Freire Martín, I.; AbuOun, M.; La Ragione, R.M.; Reichel, R.; Woodward, M.J. Sequence analysis of a CTX-M-1 IncI1 plasmid found in Salmonella 4,5,12:i:-, Escherichia coli and Klebsiella pneumoniae on a UK pig farm. J. Antimicrob. Chemother. 2014, 69, 2098–2101. [Google Scholar] [CrossRef]
- Poole, T.L.; Callaway, T.R.; Norman, K.N.; Scott, H.M.; Loneragan, G.H.; Ison, S.A.; Beier, R.C.; Harhay, D.M.; Norby, B.; Nisbet, D.J. Transferability of antimicrobial resistance from multidrug-resistant Escherichia coli isolated from cattle in the USA to E. coli and Salmonella Newport recipients. J. Glob. Antimicrob. Resist. 2017, 11, 123–132. [Google Scholar] [CrossRef]
- Liu, M.Y.; Worden, P.; Monahan, L.G.; DeMaere, M.Z.; Burke, C.M.; Djordjevic, S.P.; Charles, I.G.; Darling, A.E. Evaluation of ddRADseq for reduced representation metagenome sequencing. PeerJ. 2017, 5, e3837. [Google Scholar] [CrossRef] [PubMed]
- DeMaere, M.Z.; Darling, A.E. Sim3C: Simulation of Hi-C and Meta3C proximity ligation sequencing technologies. Gigascience 2018, 7, gix103. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wyrsch, E.R.; Hawkey, J.; Judd, L.M.; Haites, R.; Holt, K.E.; Djordjevic, S.P.; Billman-Jacobe, H. Z/I1 Hybrid Virulence Plasmids Carrying Antimicrobial Resistance genes in S. Typhimurium from Australian Food Animal Production. Microorganisms 2019, 7, 299. https://doi.org/10.3390/microorganisms7090299
Wyrsch ER, Hawkey J, Judd LM, Haites R, Holt KE, Djordjevic SP, Billman-Jacobe H. Z/I1 Hybrid Virulence Plasmids Carrying Antimicrobial Resistance genes in S. Typhimurium from Australian Food Animal Production. Microorganisms. 2019; 7(9):299. https://doi.org/10.3390/microorganisms7090299
Chicago/Turabian StyleWyrsch, Ethan R., Jane Hawkey, Louise M. Judd, Ruth Haites, Kathryn E. Holt, Steven P. Djordjevic, and Helen Billman-Jacobe. 2019. "Z/I1 Hybrid Virulence Plasmids Carrying Antimicrobial Resistance genes in S. Typhimurium from Australian Food Animal Production" Microorganisms 7, no. 9: 299. https://doi.org/10.3390/microorganisms7090299
APA StyleWyrsch, E. R., Hawkey, J., Judd, L. M., Haites, R., Holt, K. E., Djordjevic, S. P., & Billman-Jacobe, H. (2019). Z/I1 Hybrid Virulence Plasmids Carrying Antimicrobial Resistance genes in S. Typhimurium from Australian Food Animal Production. Microorganisms, 7(9), 299. https://doi.org/10.3390/microorganisms7090299