Genomic Virulence Features of Two Novel Species Nocardia barduliensis sp. nov. and Nocardia gipuzkoensis sp. nov., Isolated from Patients with Chronic Pulmonary Diseases


 , , and
, , and
Abstract
:1. Introduction
2. Case Reports
2.1. Case 1
2.2. Case 2
3. Materials and Methods
3.1. Isolation and Phenotypic Characterization
3.2. Whole Genome Sequencing
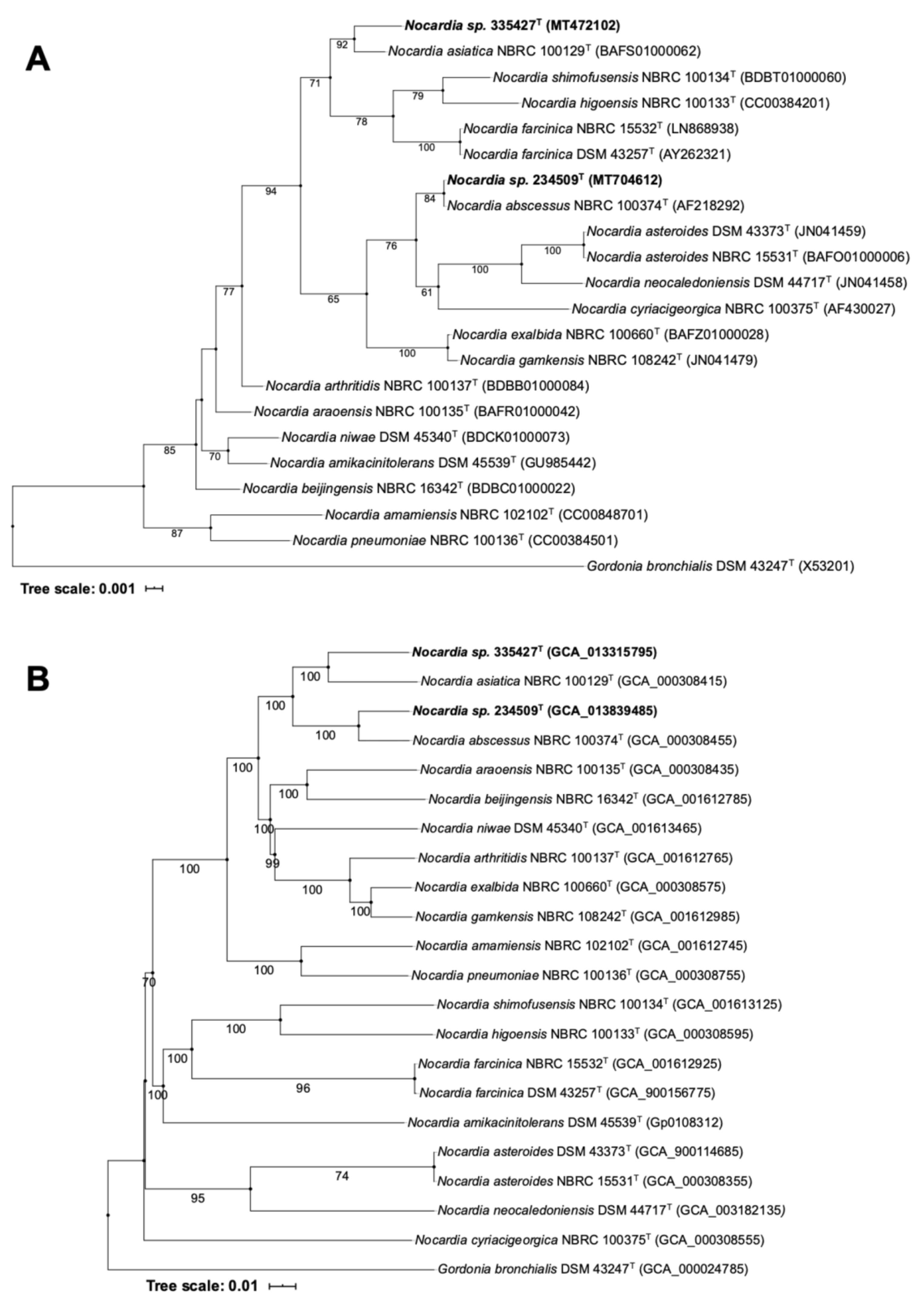
3.3. Identification and Phylogenetic Studies
3.4. Virulence Factors
- Pathogen species: Mycobacterium tuberculosis
- Disease: tuberculosis
- Host species: Homo sapiens (related to human) and Mus musculus (house mouse)
- Experimental technique: partial and full gene deletions, complementation and disruption.
- Mutant phenotype: increased virulence (hypervirulence); loss of pathogenicity; reduced virulence.
3.5. Antimicrobial Gene Resistance and Toxic Compound Tolerance
3.6. Comparative Genomic of VF-Associated Genes
4. Results and Discussion
4.1. Phenotypic Characterization
4.2. Molecular Characterization and Genome-Based Taxonomy
4.3. Virulence Factors
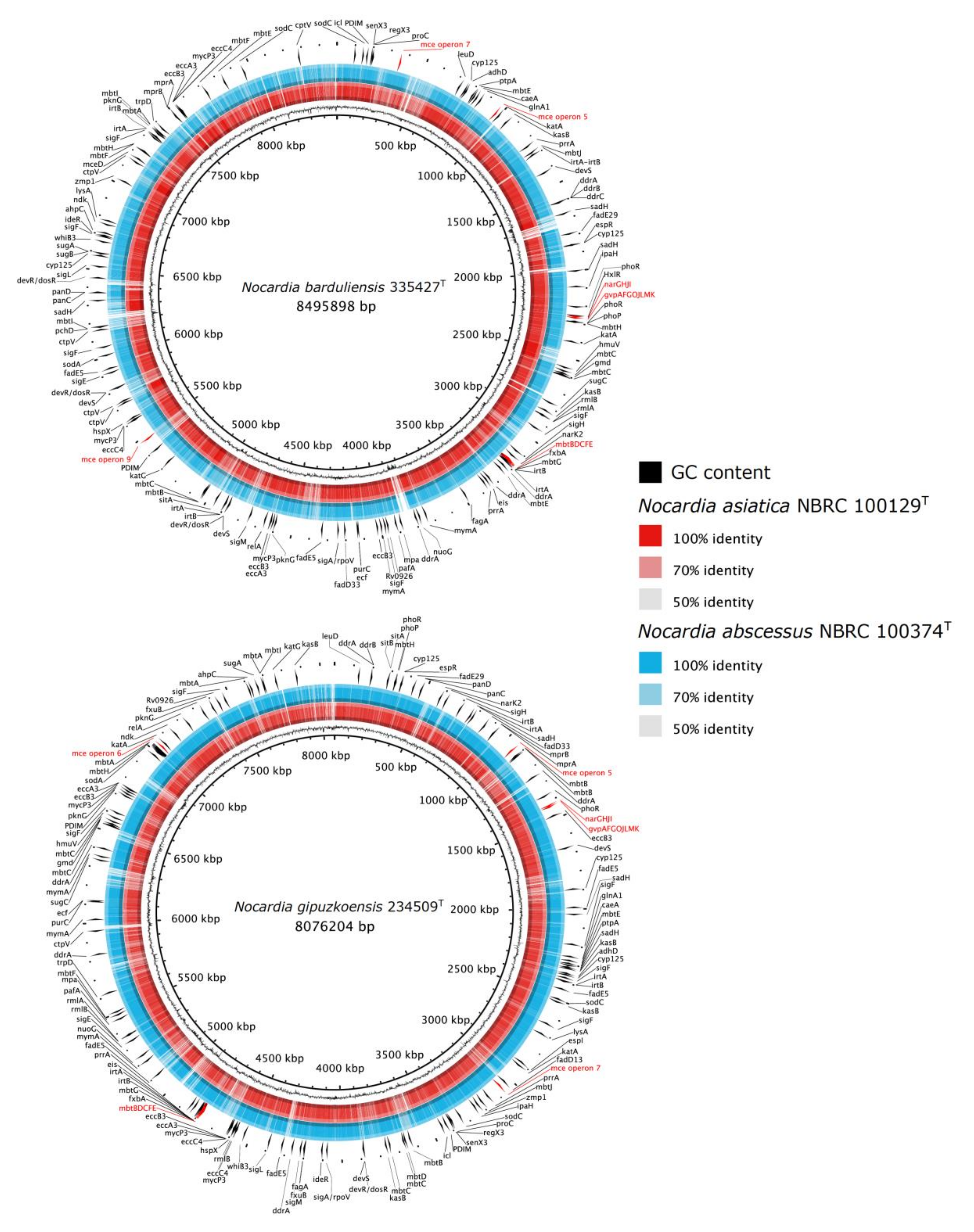
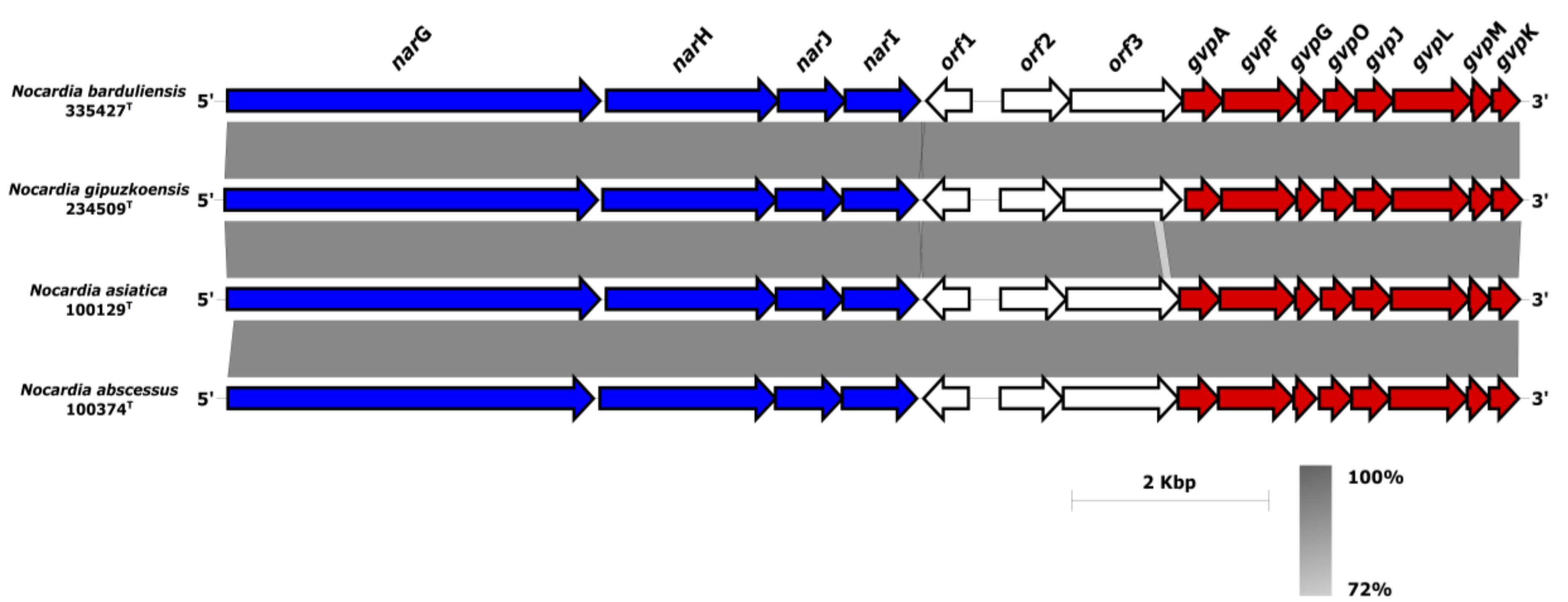
4.4. Comparative Genomic of VF-Encoding Genes
4.5. Estimation of Being Human Pathogen
4.6. Antimicrobial Gene Resistance and Toxic Compound Tolerance
4.7. Description of Nocardia barduliensis sp. nov.
4.8. Description of Nocardia gipuzkoensis sp. nov.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trevisan, V.I. Generi e le Specie delle Batteriacee; L. Zanaboni e Gabuzzi: Milano, Italy, 1889. [Google Scholar]
- Brown-Elliott, B.A.; Brown, J.M.; Conville, P.S.; Wallace, R.J. Clinical and laboratory features of the Nocardia spp. based on current molecular taxonomy. Clin. Microbiol. Rev. 2006, 19, 259–282. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, R. Parasites végétaux à l’exclusion des bactéries. In Traité de Pathologie Générale; Bouchard, C., Ed.; G. Masson: Paris, France, 1986; Volume 2, pp. 1–932. [Google Scholar]
- Poonwan, N.; Kusum, M.; Mikami, Y.; Yazawa, K.; Tanaka, Y.; Gonoi, T.; Hasegawa, S.; Konyama, K. PathogenicNocardia isolated from clinical specimens including those of AIDS patients in Thailand. Eur. J. Epidemiol. 1995, 11, 507–512. [Google Scholar] [CrossRef]
- J Javaly, K.; Horowitz, H.W.; Wormser, G.P. Nocardiosis in Patients with Human Immunodeficiency Virus Infection Report of 2 Cases and Review of the Literature. Medicine 1992, 71, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Kandi, V. Human Nocardia Infections: A Review of Pulmonary Nocardiosis. Cureus 2015, 7, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zia, K.; Nafees, T.; Faizan, M.; Salam, O.; Asad, S.I.; Khan, A.Y.; Altaf, A.; Saad, S.I. Ten Year Review of Pulmonary Nocardiosis: A Series of 55 Cases. Cureus 2019, 11, e4759. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.; McNeil, M.M.; Brown, J.M. Nocardia species (Nocardiosis). Antimicrobe. Available online: http://www.antimicrobe.org/b117.asp (accessed on 18 August 2020).
- Yıldız, O.; Doganay, M.; Yildiz, O. Actinomycoses and Nocardia pulmonary infections. Curr. Opin. Pulm. Med. 2006, 12, 228–234. [Google Scholar] [CrossRef]
- Rahdar, H.A.; Azadi, D.; Shojaei, H.; Daei-Naser, A. Molecular analysis and species diversity of Nocardia in the hospital environment in a developing country, a potential health hazard. J. Med. Microbiol. 2017, 66, 334–341. [Google Scholar] [CrossRef]
- Beaman, B.L.; Burnside, J.; Edwards, B.; Causey, W. Nocardial Infections in the United States, 1972–1974. J. Infect. Dis. 1976, 134, 286–289. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, A.M.; Fida, M.; Deml, S.M.; Abu Saleh, O.M.; Wengenack, N.L. Retrospective Analysis of Antimicrobial Susceptibility Profiles of Nocardia Species from a Tertiary Hospital and Reference Laboratory, 2011 to 2017. Antimicrob. Agents Chemother. 2019, 64, e01868-19. [Google Scholar] [CrossRef]
- Davis-Scibienski, C.; Beaman, B.L. Interaction of Nocardia asteroides with Rabbit Alveolar Macrophages: Association of Virulence, Viability, Ultrastructural Damage, and Phagosome-Lysosome Fusion. Infect. Immun. 1980, 28, 610–619. [Google Scholar]
- Vera-Cabrera, L.; Ortiz-Lopez, R.; Elizondo-Gonzalez, R.; Ocampo-Candiani, J. Complete Genome Sequence Analysis of Nocardia brasiliensis HUJEG-1 Reveals a Saprobic Lifestyle and the Genes Needed for Human Pathogenesis. PLoS ONE 2013, 8, e65425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaman, B.L.; Black, C.M.; Doughty, F.; Beaman, L. Role of superoxide dismutase and catalase as determinants of pathogenicity of Nocardia asteroides: Importance in resistance to microbicidal activities of human polymorphonuclear neutrophils. Infect. Immun. 1985, 47, 135–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 1–6. [Google Scholar] [CrossRef]
- Metchock, B.G.; Nolte, F.S.; Wallace, R.J. Mycobacterium. In Manual of Clinical Microbiology; Murray, P.M., Baron, E.J., Pfaller, M.A., Tenover, F.C., Yolken, R.H., Eds.; American Society for Microbiology: Washington, DC, USA, 1999; pp. 399–437. [Google Scholar]
- Idigoras, P.; Beristain, X.; Iturzaeta, A.; Vicente, D.; Pérez-Trallero, E. Comparison of the automated nonradiometric Bactec MGIT 960 system with Löwenstein-Jensen, Coletsos, and Middlebrook 7H11 solid media for recovery of mycobacteria. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Shirling, E.B.; Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef] [Green Version]
- Atlas, R.M.; Snyder, J.W. Handbook of Media for Clinical Microbiology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Siddiqi, S.H.; Rüsch-Gerdes, S. Procedure Manual for BACTECTM MGIT 960 TB System. Find Foundation for Innovative New Diagnostics; FIND: Geneva, Switzerland, 2006. [Google Scholar]
- MacFaddin, J.F. Media for Isolation–Cultivation–Identification–Maintenance of Medical Bacteria; Williams and Wilkins: Baltimore, MD, USA, 1985. [Google Scholar]
- Tomioka, H.; Saito, H.; Sato, K.; Dawson, D.J. Arylsulfatase activity for differentiating Mycobacterium avium and Mycobacterium intracellulare. J. Clin. Microbiol. 1990, 28, 2104–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, P.T.; Kubica, G.P. Public Health Mycobacteriology a Guide for the Level III Laboratory; Centers for Disease Control and Prevention: Atlanta, GA, USA, 1985. [Google Scholar]
- Kilburn, J.O.; Silcox, V.A.; Kubica, G.P. Differential identification of Mycobacteria. V. The tellurite reduction test. Am. Rev. Res. Dis. 1969, 99, 94–100. [Google Scholar]
- Vaas, L.A.I.; Sikorski, J.; Hofner, B.; Fiebig, A.; Buddruhs, N.; Klenk, H.-P.; Göker, M. opm: An R package for analysing OmniLog(R) phenotype microarray data. Bioinformatics 2013, 29, 1823–1824. [Google Scholar] [CrossRef] [Green Version]
- Woods, G.L.; Brown-Elliott, B.A.; Conville, P.S.; Desmond, E.P.; Hall, G.S.; Lin, G.; Pfyffer, G.E.; Ridderhof, J.C.; Siddiqi, S.H.; Wallace, R.J.; et al. Susceptibility Testing of Mycobacteria, Nocardiae, and Other Aerobic Actinomycetes, 2nd ed.; No. 31.5; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011. [Google Scholar]
- Minnikin, D.; O’Donnell, A.; Goodfellow, M.; Alderson, G.; Athalye, M.; Schaal, A.; Parlett, J. An integrated procedure for the extraction of bacterial isoprenoid quinones and polar lipids. J. Microbiol. Methods 1984, 2, 233–241. [Google Scholar] [CrossRef]
- Ambaye, A.; Kohner, P.C.; Wollan, P.C.; Roberts, K.L.; Roberts, G.D.; Cockerill, F.R. Comparison of agar dilution, broth microdilution, disk diffusion, E-test, and BACTEC radiometric methods for antimicrobial susceptibility testing of clinical isolates of the Nocardia asteroides complex. J. Clin. Microbiol. 1997, 35, 847–852. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.T. Single derivatization method for routine analysis of bacterial whole-cell fatty acid methyl esters, including hydroxy acids. J. Clin. Microbiol. 1982, 16, 584–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minnikin, D.E.; Goodfellow, M. Lipid composition in the classification and identification of nocardiae and related taxa. In The Biology of the Nocardiae; Goodfellow, M., Brownell, G.H., Serrano, J.A., Eds.; Academic Press: London, UK, 1976; pp. 160–219. [Google Scholar]
- Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids, Technical note 101, DE, MID, 1990.
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.Y.; Glass, E.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genomics 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; Da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.-W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M.; Spröer, C.; Klenk, H.-P. When should a DDH experiment be mandatory in microbial taxonomy? Arch. Microbiol. 2013, 195, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Hahnke, R.L.; Petersen, J.; Scheuner, C.; Michael, V.; Fiebig, A.; Rohde, C.; Rohde, M.; Fartmann, B.; Goodwin, L.A.; et al. Complete genome sequence of DSM 30083T, the type strain (U5/41T) of Escherichia coli, and a proposal for delineating subspecies in microbial taxonomy. Stand. Genomic Sci. 2014, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis--10 years on. Nucleic Acids Res. 2015, 44, D694–D697. [Google Scholar] [CrossRef] [PubMed]
- Wassenaar, T.M.; Gaastra, W. Bacterial virulence: Can we draw the line? FEMS Microbiol. Lett. 2001, 201, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.; Cuzick, A.; Seager, J.; Wood, V.; Rutherford, K.; Venkatesh, S.Y.; De Silva, N.; Martinez, M.C.; Pedro, H.; Yates, A.; et al. PHI-base: The pathogen–host interactions database. Nucleic Acids Res. 2019, 48, D613–D620. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Zheng, C.; Chitsaz, F.; Derbyshire, M.K.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Lanczycki, C.J.; et al. CDD: Conserved domains and protein three-dimensional structure. Nucleic Acids Res. 2012, 41, D348–D352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosentino, S.; Voldby, M.V.; Aarestrup, F.M.; Lund, O. PathogenFinder—Distinguishing friend from foe using bacterial whole genome sequence data. PLoS ONE 2013, 8, e77302. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2013, 42, D206–D214. [Google Scholar] [CrossRef]
- Berriman, M.; Rutherford, K.M. Viewing and annotating sequence data with Artemis. Brief. Bioinform. 2003, 4, 124–132. [Google Scholar] [CrossRef]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genomics 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C.; Patel, P.V. The Isolation, Properties and Taxonomic Relevance of Lipid-soluble, Iron-binding Compounds (the Nocobactins) from Nocardia. J. Gen. Microbiol. 1976, 93, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, M.; Maldonado, L.A.; Genus, I. Nocardia Trevisan 1889AL. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Whitman, W.B., Goodfellow, M., Kaempfer, P., Busse, H.-J., Trujillo, M.E., Ludwig, W., Suzuki, K.-I., Eds.; Springer: New York, NY, USA, 2012; Volume 5, pp. 376–421. [Google Scholar]
- Kageyama, A.; Yazawa, K.; Mukai, A.; Kohara, T.; Nishimura, K.; Kroppenstedt, R.M.; Mikami, Y. Nocardia araoensis sp. nov. and Nocardia pneumoniae sp. nov., isolated from patients in Japan. Int. J. Syst. Evol. Microbiol. 2004, 54, 2025–2029. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Y.; Lu, Z.; Shi, Y.; Liu, Z.; Maldonado, L.; Goodfellow, M. Nocardia beijingensis sp. nov., a novel isolate from soil. Int. J. Syst. Evol. Microbiol. 2001, 51, 1783–1788. [Google Scholar] [CrossRef]
- Reann Wai-Po, C.; David, L.; Sik-Nin, W. Pulmonary abscess caused by Nocardia beijingensis: The second report of human infection. Pediatr. Infect. Dis. J. 2008, 27, 572–573. [Google Scholar]
- Moser, B.D.; Klenk, H.-P.; Schumann, P.; Pötter, G.; Lasker, B.A.; Steigerwalt, A.G.; Hinrikson, H.P.; Brown, J.M. Nocardia niwae sp. nov., isolated from human pulmonary sources. Int. J. Syst. Evol. Microbiol. 2011, 61, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, A.; Torikoe, K.; Iwamoto, M.; Masuyama, J.-I.; Shibuya, Y.; Okazaki, H.; Yazawa, K.; Minota, S.; Kroppenstedt, R.M.; Mikami, Y. Nocardia arthritidis sp. nov., a New Pathogen Isolated from a Patient with Rheumatoid Arthritis in Japan. J. Clin. Microbiol. 2004, 42, 2366–2371. [Google Scholar] [CrossRef] [Green Version]
- Iida, S.; Kageyama, A.; Yazawa, K.; Uchiyama, N.; Toyohara, T.; Chohnabayashi, N.; Suzuki, S.-I.; Nomura, F.; Kroppenstedt, R.M.; Mikami, Y. Nocardia exalbida sp. nov., isolated from Japanese patients with nocardiosis. Int. J. Syst. Evol. Microbiol. 2006, 56, 1193–1196. [Google Scholar] [CrossRef]
- Le Roes, M.; Meyers, P.R.; Le Roes-Hill, M. Nocardia gamkensis sp. nov. Antonie Leeuwenhoek 2006, 90, 291–298. [Google Scholar] [CrossRef]
- Roberts, A.L.; Davidson, R.M.; Freifeld, A.G.; Iwen, P.C. Nocardia arthritidis as a cause of disseminated nocardiosis in a patient with chronic lymphocytic leukemia. IDCases 2016, 6, 68–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuchibiro, T.; Ikeda, T.; Nakanishi, H.; Morishita, Y.; Houdai, K.; Ito, J.; Gonoi, T. First case report of pulmonary nocardiosis caused by Nocardia mexicana. JMM Case Rep. 2016, 3, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wayne, L.G.; Moore, W.E.C.; Stackebrandt, E.; Kandler, O.; Colwell, R.R.; Krichevsky, M.I.; Trüper, H.G.; Murray, R.G.E.; Grimont, P.A.D.; Brenner, D.J.; et al. Report of the Ad Hoc Committee on Reconciliation of Approaches to Bacterial Systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Carrillo, C.; Millan-Sauceda, C.; Lozano-Garza, H.G.; Ortiz-Lopez, R.; Elizondo-Gonzalez, R.; Welsh, O.; Ocampo-Candiani, J.; Vera-Cabrera, L. Genomic Changes Associated with the Loss of Nocardia brasiliensis Virulence in Mice after 200In VitroPassages. Infect. Immun. 2016, 84, 2595–2606. [Google Scholar] [CrossRef] [Green Version]
- Yassin, A.; Rainey, F.A.; Mendrock, U.; Brzezinka, H.; Schaal, K.P. Nocardia abscessus sp. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 1487–1493. [Google Scholar] [CrossRef]
- Lella, R.K.; Sharma, C. Eis (Enhanced Intracellular Survival) Protein ofMycobacterium tuberculosisDisturbs the Cross Regulation of T-cells. J. Biol. Chem. 2007, 282, 18671–18675. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Rao, X.; Zhang, K. Nucleoside diphosphate kinase (Ndk): A pleiotropic effector manipulating bacterial virulence and adaptive responses. Microbiol. Res. 2017, 205, 125–134. [Google Scholar] [CrossRef]
- Velmurugan, K.; Chen, B.; Miller, J.L.; Azogue, S.; Gurses, S.; Hsu, T.; Glickman, M.; Jacobs, W.R., Jr.; Porcelli, S.A.; Briken, V. Mycobacterium tuberculosis nuoG Is a Virulence Gene That Inhibits Apoptosis of Infected Host Cells. PLoS Pathog. 2007, 3, e110. [Google Scholar] [CrossRef] [Green Version]
- Master, S.S.; Springer, B.; Sander, P.; Boettger, E.C.; Deretic, V.; Timmins, G.S. Oxidative stress response genes in Mycobacterium tuberculosis: Role of ahpC in resistance to peroxynitrite and stage-specific survival in macrophages. Microbiology 2002, 148, 3139–3144. [Google Scholar] [CrossRef] [Green Version]
- Fenn, K.; Wong, C.T.; Darbari, V.C. Mycobacterium tuberculosis Uses Mce Proteins to Interfere with Host Cell Signaling. Front. Mol. Biosci. 2020, 6, 149. [Google Scholar] [CrossRef] [Green Version]
- Yasuike, M.; Nishiki, I.; Iwasaki, Y.; Nakamura, Y.; Fujiwara, A.; Shimahara, Y.; Kamaishi, T.; Yoshida, T.; Nagai, S.; Kobayashi, T.; et al. Analysis of the complete genome sequence of Nocardia seriolae UTF1, the causative agent of fish nocardiosis: The first reference genome sequence of the fish pathogenic Nocardia species. PLoS ONE 2017, 12, e0173198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.; Sarkar, D. Nitrate reduction pathways in mycobacteria and their implications during latency. Microbiology 2012, 158, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Khan, A.; Sohaskey, C.D.; Jagannath, C.; Sarkar, D. Nitrite Reductase NirBD Is Induced and Plays an Important Role during In Vitro Dormancy of Mycobacterium tuberculosis. J. Bacteriol. 2013, 195, 4592–4599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeifer, F. Gas Vesicles of Archaea and Bacteria. In Complex Intracellular Structures in Prokaryotes, Microbiology Monographs; Shively, J.M., Ed.; Springer: Berlin, Germany, 2006; Volume 2, pp. 115–140. [Google Scholar]
- Van Keulen, G.; Hopwood, D.A.; Dijkhuizen, L.; Sawers, R.G. Gas vesicles in actinomycetes: Old buoys in novel habitats? Trends Microbiol. 2005, 13, 350–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.-J.; Kwak, M.-J.; Ha, S.-M.; Yang, S.-J.; Kim, J.D.; Cho, K.-H.; Kim, T.-W.; Cho, M.Y.; Kim, B.-Y.; Jung, S.-H.; et al. Genomic characterization of Nocardia seriolae strains isolated from diseased fish. Microbiology 2018, 8, e00656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Iglesia, P.; Viejo, G.; Gómez, B.; De Miguel, D.; Del Valle, A.; Otero, L. Fatal Pulmonary Nocardia farcinica Infection. J. Clin. Microbiol. 2002, 40, 1098–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Zhang, X.; Du, P.; Li, G.; Li, L.; Li, Z. Susceptibility profiles of Nocardia spp. to antimicrobial and antituberculotic agents detected by a microplate Alamar Blue assay. Sci. Rep. 2017, 7, 43660. [Google Scholar] [CrossRef]
- Cornforth, D.M.; Dees, J.L.; Ibberson, C.B.; Huse, H.K.; Mathiesen, I.H.; Kirketerp-Møller, K.; Wolcott, R.D.; Rumbaugh, K.P.; Bjarnsholt, T.; Whiteley, M. Pseudomonas aeruginosa transcriptome during human infection. Proc. Natl. Acad. Sci. USA 2018, 115, E5125–E5134. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.H.; Butler, M.S.; Ranzoni, A.; Cooper, M.A. Detection and quantification of the heterogeneity of S. aureus bacterial populations to identify antibiotic-induced persistence. bioRxiv 2018. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | Isolate 335427T | N. asiatica DSM 44668T | Isolate 234509T | N. abscessus DSM 44432T |
|---|---|---|---|---|
| Carbon source utilization | ||||
| β-Gentiobiose and L-Fucose | + | + | - | - |
| β-Methyl-D-Glucoside | + | w | w | + |
| Dextrin | - | + | - | + |
| D-Arabitol | w | + | - | - |
| D-Cellobiose | - | - | - | w |
| D-Fructose, D-Fructose-6-Phosphate, N-Acetyl-β-D-Mannosamine and L-Rhamnose | + | + | - | + |
| D-Fucose; D-Galactose, D-Mannose, Inosine, myo-Inositol, 3-O-Methyl-D-Glucose and N-Acetyl-D-Galactosamine | + | - | - | - |
| D-Galacturonic Acid and Glucuronamide | - | w | - | - |
| D-Glucose and Turanose | w | + | + | + |
| D-Glucose-6-Phosphate | + | - | + | + |
| D-Maltose | w | - | + | + |
| D-Melibiose | w | + | - | + |
| D-Saccharic Acid and N-Acetyl-D-Glucosamine | + | w | + | + |
| D-Salicin | w | w | + | + |
| D-Trehalose | - | - | + | w |
| Gelatin | - | + | - | - |
| N-Acetyl-Neuraminic Acid | + | w | - | - |
| Pectin | + | w | - | w |
| Aminoacids | ||||
| L-Histidine | + | + | - | + |
| L-Serine | w | - | - | + |
| Organic acids | ||||
| Acetoacetic Acid | + | w | + | + |
| α-Hydroxy-Butyric Acid | + | + | - | + |
| α-Keto-Butyric Acid | + | - | - | + |
| Bromo-Succinic Acid | - | - | + | + |
| Butyric Acid | w | - | w | + |
| D-Gluconic Acid | w | + | - | - |
| D-Glucuronic Acid | + | w | + | - |
| D-Lactic Acid Methyl Ester | + | - | w | - |
| D-Malic Acid | + | + | + | w |
| L-Lactic Acid | + | + | w | - |
| Mucic Acid | + | + | - | - |
| Methyl Pyruvate | w | - | + | + |
| p-Hydroxy-Phenylacetic Acid | - | - | - | + |
| Inhibitory compounds | ||||
| Lithium Chloride | - | - | - | w |
| Sodium Bromate | - | - | - | + |
| Tetrazolium Violet | w | - | + | - |
| Vancomycin | - | - | - | + |
| 1% Sodium Lactate | w | - | + | + |
| Gene Identity (%) | |||||||
|---|---|---|---|---|---|---|---|
| Isolate | Hit Taxon | 16S rRNA | hsp65 | secA | gyrB | rpoB | dDDH |
| 335427T | |||||||
| Isolate 234509T | 99.0 | 98.7 | 97.1 | 92.9 | 99.8 | 44.2 | |
| N. asiatica | 99.5 | 98.1 | 98.5 | 94.4 | 99.7 | 53.5 | |
| N. arthritidis | 99.0 | 96.2 | 97.0 | 92.8 | 96.0 | 36.6 | |
| N. abscessus | 98.9 | 97.5 | 97.9 | 93.1 | 99.4 | 44.6 | |
| N. farcinica | 98.8 | 92.9 | 88.5 | 91.9 | 97.2 | 23.3 | |
| N. kroppenstedtii | 98.7 | - | - | - | - | N/A | |
| N. araoensis | 98.6 | 98.3 | 97.0 | 98.5 | 35.7 | ||
| N. beijingensis | 98.4 | 97.9 | 97.6 | 93.5 | 98.2 | 35.0 | |
| 234509T | |||||||
| Isolate 335427T | 99.0 | 98.7 | 97.1 | 92.9 | 99.8 | 44.2 | |
| N. abscessus | 100.0 | 98.5 | 98.5 | 99.8 | 99.4 | 67.9 | |
| N. asiatica | 99.0 | 98.3 | 98.2 | 97.9 | 99.2 | 43.0 | |
| N. exalbida | 99.0 | 98.8 | 96.4 | 97.9 | 99.0 | 37.6 | |
| N. cyriacigeorgica | 98.9 | 96.3 | 87.9 | 93.3 | 97.5 | 22.9 | |
| N. shimofusensis | 98.9 | 96.8 | 85.9 | 92.8 | 97.6 | 22.5 | |
| N. asteroides | 98.7 | 95.5 | 98.5 | 92.7 | 92.7 | 22.1 | |
| N. arthritidis | 98.5 | 98.5 | 96.5 | 91.9 | 99.1 | 37.6 | |
| N. neocaledoniensis | 98.3 | 95.9 | 86.5 | 92.6 | 96.4 | 22.1 | |
| Antibiotic | Susceptibility Criteria 1 | Isolate 234509T | Isolate 335427T |
|---|---|---|---|
| Penicillin 2 | ND | 0.25 | 0.25 |
| Amoxicillin 2 | ND | 0.38 | 0.75 |
| Amoxicillin-clavulanic acid | ≤8/4 | <2/1 | 16/8 |
| Ceftriaxone | ≤8 | <4 | <4 |
| Cefepime | ≤8 | <1 | 2 |
| Imipenem | ≤4 | <2 | <2 |
| Clarithromycin | ≤2 | 2 | 4 |
| Doxycycline | ≤1 | <0.12 | <0.12 |
| Minocycline | ≤1 | <1 | <1 |
| Trimethoprim/sulfamethoxazole | ≤2/38 | <0.25/4.75 | <0.25/4.75 |
| Ciprofloxacin | ≤1 | >4 | 1 |
| Levofloxacin | ≤1 | 4 | 0.5 |
| Tobramycin | ≤4 | <1 | <1 |
| Amikacin | ≤8 | <1 | <1 |
| Tigecycline | ND | 0.06 | 0.25 |
| Linezolid | ≤8 | <1 | <1 |
| Isolate | Gene | AMR Gene Family | Drug Class | Resistance Mechanism | Identity of Matching Region (%) |
|---|---|---|---|---|---|
| 335427T | |||||
| c | small multidrug resistance (SMR) antibiotic efflux pump | aminoglycoside antibiotic, tetracycline antibiotic, phenicol antibiotic | antibiotic efflux | 100 | |
| tet44 | tetracycline-resistant ribosomal protection protein | tetracycline | target protection | ||
| lpe | resistance-nodulation-cell division (RND) antibiotic efflux pump | - | efflux | 100 | |
| vanHM | glycopeptide antibiotic | - | target alteration | 100 | |
| 234509T | |||||
| - | EmrB/QacA subfamily drug resistance transporter | - | 85.9 | ||
| - | MurA regulator CwlM implicated in cell wall metabolism and antibiotic tolerance | - | 99.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nouioui, I.; Cortés-Albayay, C.; Neumann-Schaal, M.; Vicente, D.; Cilla, G.; Klenk, H.-P.; Marimón, J.M.; Ercibengoa, M. Genomic Virulence Features of Two Novel Species Nocardia barduliensis sp. nov. and Nocardia gipuzkoensis sp. nov., Isolated from Patients with Chronic Pulmonary Diseases. Microorganisms 2020, 8, 1517. https://doi.org/10.3390/microorganisms8101517
Nouioui I, Cortés-Albayay C, Neumann-Schaal M, Vicente D, Cilla G, Klenk H-P, Marimón JM, Ercibengoa M. Genomic Virulence Features of Two Novel Species Nocardia barduliensis sp. nov. and Nocardia gipuzkoensis sp. nov., Isolated from Patients with Chronic Pulmonary Diseases. Microorganisms. 2020; 8(10):1517. https://doi.org/10.3390/microorganisms8101517
Chicago/Turabian StyleNouioui, Imen, Carlos Cortés-Albayay, Meina Neumann-Schaal, Diego Vicente, Gustavo Cilla, Hans-Peter Klenk, Jose María Marimón, and Maria Ercibengoa. 2020. "Genomic Virulence Features of Two Novel Species Nocardia barduliensis sp. nov. and Nocardia gipuzkoensis sp. nov., Isolated from Patients with Chronic Pulmonary Diseases" Microorganisms 8, no. 10: 1517. https://doi.org/10.3390/microorganisms8101517
APA StyleNouioui, I., Cortés-Albayay, C., Neumann-Schaal, M., Vicente, D., Cilla, G., Klenk, H.-P., Marimón, J. M., & Ercibengoa, M. (2020). Genomic Virulence Features of Two Novel Species Nocardia barduliensis sp. nov. and Nocardia gipuzkoensis sp. nov., Isolated from Patients with Chronic Pulmonary Diseases. Microorganisms, 8(10), 1517. https://doi.org/10.3390/microorganisms8101517