Microbial Diversity and Nutritional Properties of Persian “Yellow Curd” (Kashk zard), a Promising Functional Fermented Food





Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Processing
2.2. pH Measurement
2.3. 16S rRNA Gene Amplicon Target Sequencing
2.4. Chemical and Mineral Compositions of YC Samples
2.5. Quantification of Sugar Content
2.6. Statistical Analysis
3. Results and Discussion
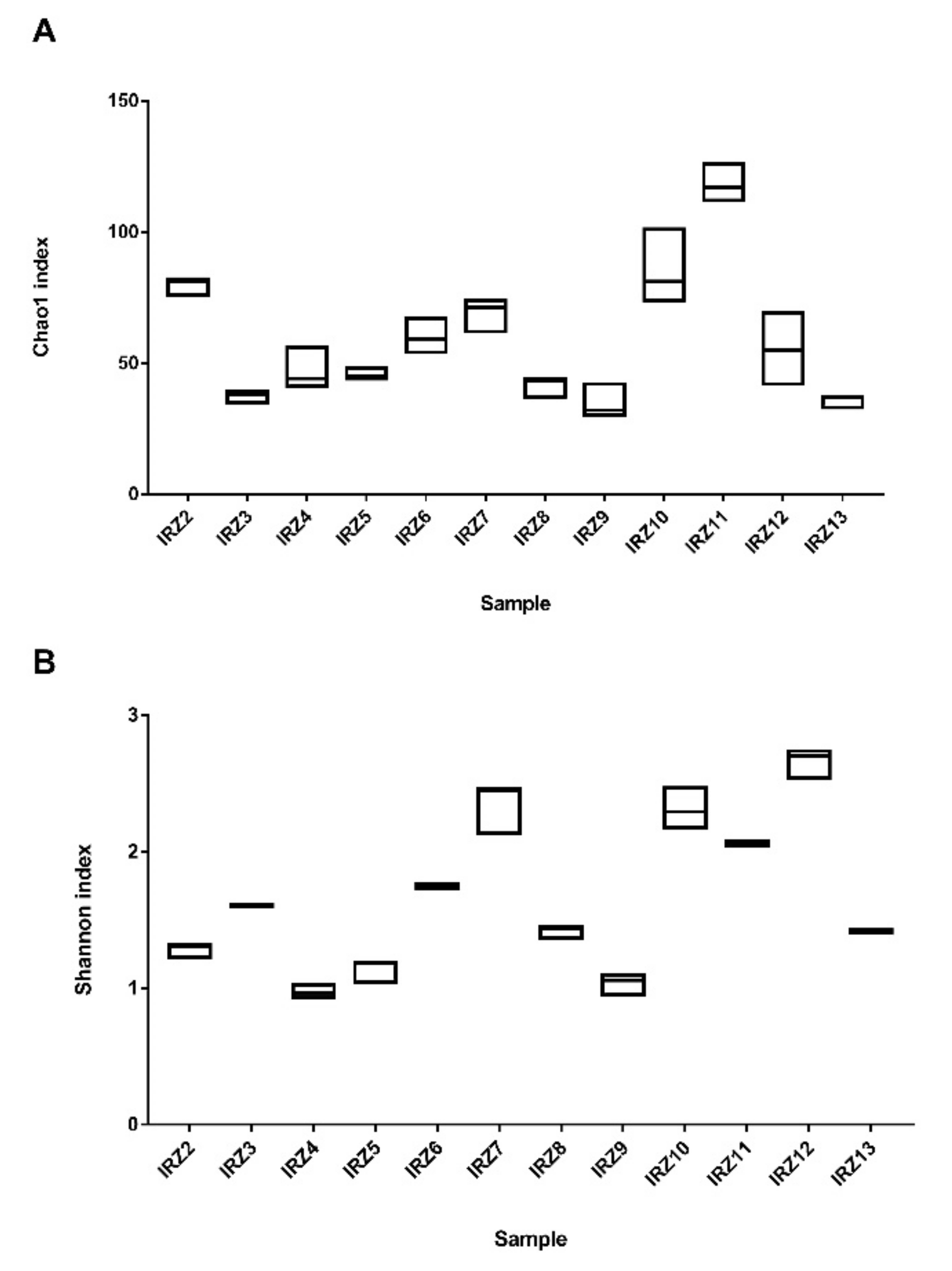
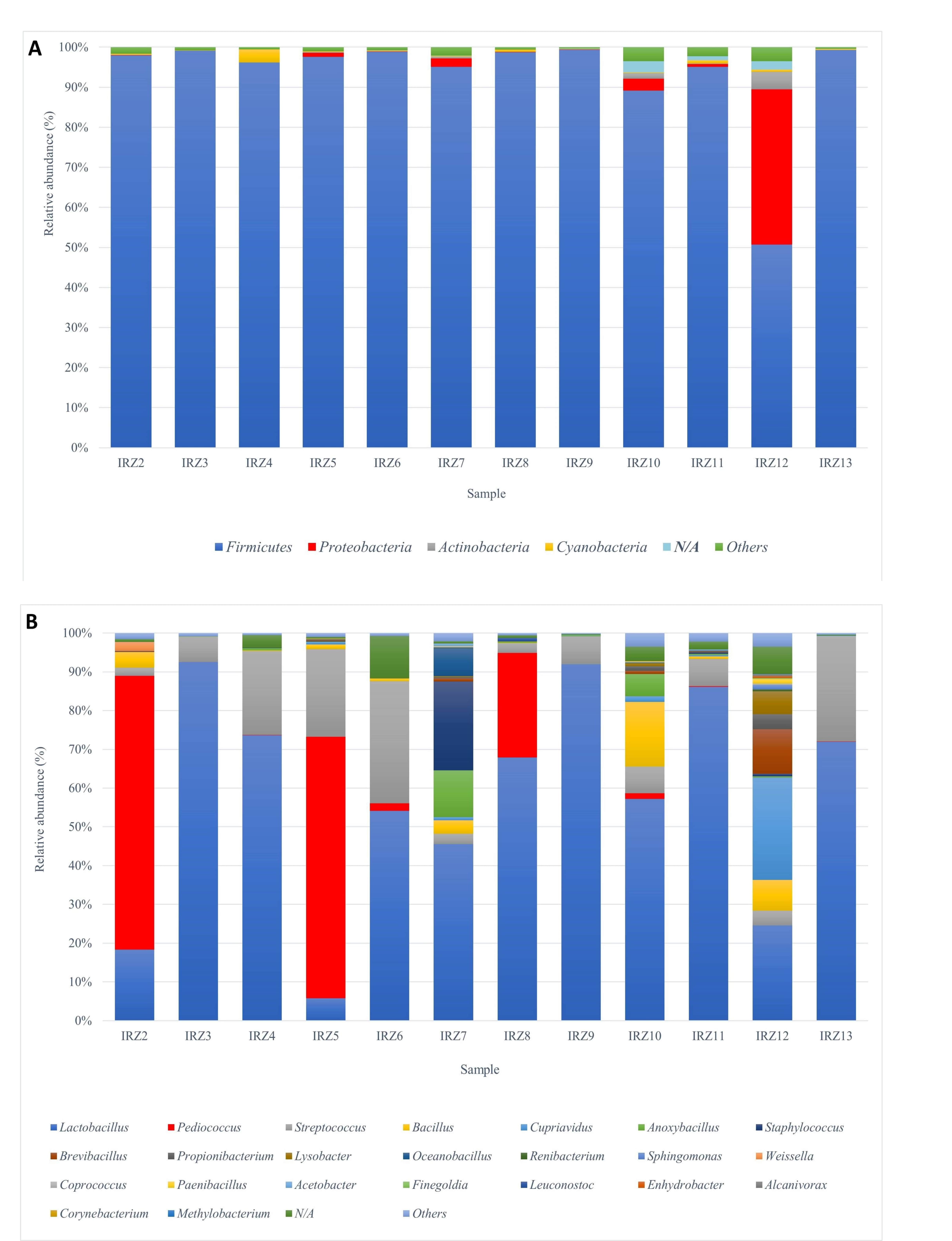
3.1. 16S rRNA Gene Amplicon Target Sequencing
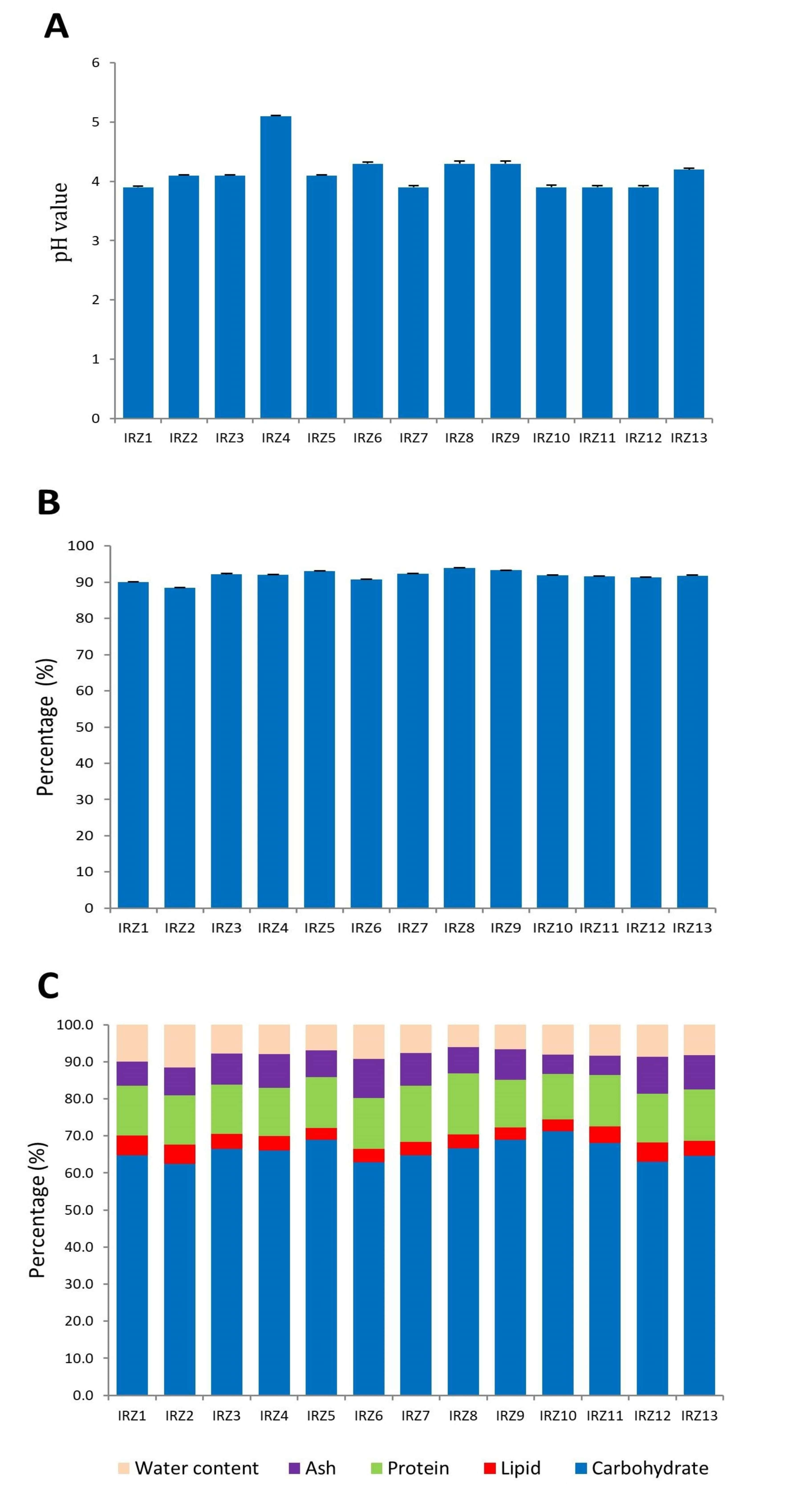
3.2. Chemical Properties of YC Samples
3.3. Mineral Content of YC Samples
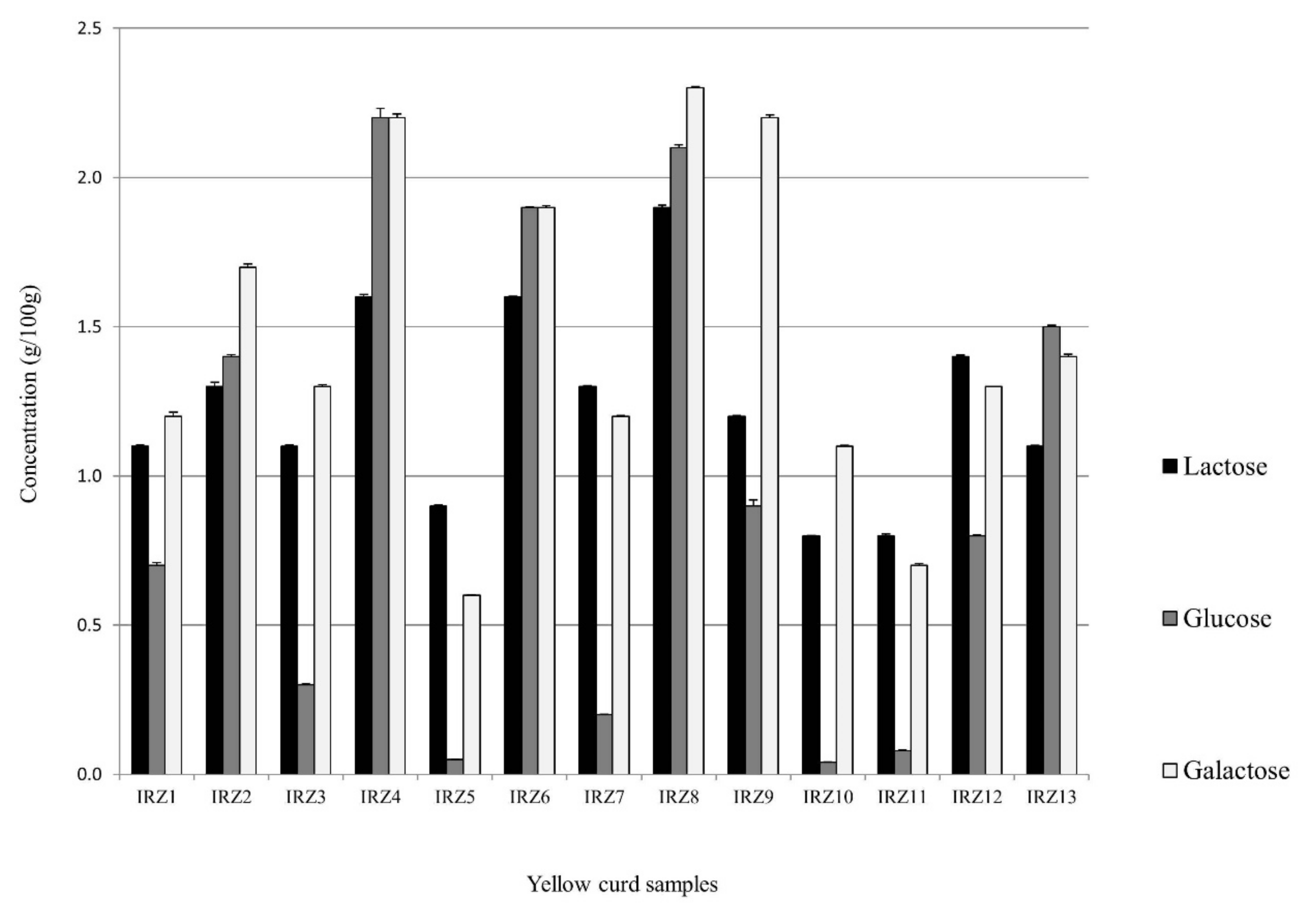
3.4. Sugar Content
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zaika, L.L.; Zell, T.E.; Smith, J.L.; Palumbo, S.A.; Kissinger, J.C. The role of nitrite and nitrate in Lebanon Bologna, a fermented sausage. J. Food Sci. 1976, 41, 1457–1460. [Google Scholar] [CrossRef]
- William, H. Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 2000. [Google Scholar]
- Poitevin, E. Official Methods for the Determination of Minerals and Trace Elements in Infant Formula and Milk Products: A Review. J. AOAC Int. 2016, 99, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Kot, M.; Daniel, W.A. The relative contribution of human cytochrome P450 isoforms to the four caffeine oxidation pathways: An in vitro comparative study with cDNA-expressed P450s including CYP2C isoforms. Biochem. Pharmacol. 2008, 76, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]
- Shangpliang, H.N.J.; Rai, R.; Keisam, S.; Jeyaram, K.; Tamang, J.P. Bacterial community in naturally fermented milk products of Arunachal Pradesh and Sikkim of India analysed by high-throughput amplicon sequencing. Sci. Rep. 2018, 8, 1532. [Google Scholar] [CrossRef]
- Zhong, Z.; Hou, Q.; Kwok, L.; Yu, Z.; Zheng, Y.; Sun, Z.; Menghe, B.; Zhang, H. Bacterial microbiota compositions of naturally fermented milk are shaped by both geographic origin and sample type. J. Dairy Sci. 2016, 99, 7832–7841. [Google Scholar] [CrossRef]
- Li, N.; Wang, Y.; You, C.; Ren, J.; Chen, W.; Zheng, H.; Liu, Z. Variation in Raw Milk Microbiota Throughout 12 Months and the Impact of Weather Conditions. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, W.; Bao, Q.; Zhang, J.; Hou, Q.; Kwok, L.; Sun, T.; Zhang, H. Investigation of bacterial and fungal diversity in tarag using high-throughput sequencing. J. Dairy Sci. 2014, 97, 6085–6096. [Google Scholar] [CrossRef]
- Guo, L.; Ya, M.; Guo, Y.-S.; Xu, W.; Li, C.-D.; Sun, J.-P.; Zhu, J.-J.; Qian, J.-P. Study of bacterial and fungal community structures in traditional koumiss from Inner Mongolia. J. Dairy Sci. 2019, 102, 1972–1984. [Google Scholar] [CrossRef]
- Lew, L.-C.; Hor, Y.-Y.; Yusoff, N.A.A.; Choi, S.-B.; Yusoff, M.S.B.; Roslan, N.S.; Ahmad, A.; Mohammad, J.A.; Abdullah, M.F.I.; Zakaria, N.; et al. Probiotic Lactobacillus plantarum P8 alleviated stress and anxiety while enhancing memory and cognition in stressed adults: A randomised, double-blind, placebo-controlled study. Clin. Nutr. 2019, 38, 2053–2064. [Google Scholar] [CrossRef]
- Joghataei, M.; Shahidi, F.; Pouladfar, G.; Mortazavi, S.A.; Ghaderi, A. Probiotic potential comparison of Lactobacillus strains isolated from Iranian traditional food products and human feces with standard probiotic strains. J. Sci. Food Agric. 2019, 99, 6680–6688. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G.; Zheng, J. Lifestyles of sourdough lactobacilli—Do they matter for microbial ecology and bread quality? Int. J. Food Microbiol. 2019, 302, 15–23. [Google Scholar] [CrossRef]
- Howe, E.; Holton, K.; Nair, S.; Schlauch, D.; Sinha, R.; Quackenbush, J. MeV: MultiExperiment Viewer. In Biomedical Informatics for Cancer Research; Ochs, M.F., Casagrande, J.T., Davuluri, R.V., Eds.; Springer: Boston, MA, USA, 2010; pp. 267–277. [Google Scholar]
- Bartkiene, E.; Bartkevics, V.; Krungleviciute, V.; Juodeikiene, G.; Zadeike, D.; Baliukoniene, V.; Bakutis, B.; Zelvyte, R.; Santini, A.; Cizeikiene, D. Application of hydrolases and probiotic Pediococcus acidilactici BaltBio01 strain for cereal by-products conversion to bioproduct for food/feed. Int. J. Food Sci. Nutr. 2017, 69, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Fleming, H.P.; Etchells, J.L.; Costilow, R.N. Microbial Inhibition by an Isolate of Pediococcus from Cucumber Brines 1. Appl. Microbiol. 1975, 30, 1040–1042. [Google Scholar] [CrossRef][Green Version]
- Papagianni, M.; Anastasiadou, S. Pediocins: The bacteriocins of Pediococci. Sources, production, properties and applications. Microb. Cell Factories 2009, 8, 3. [Google Scholar] [CrossRef]
- Mizoguchi, T.; Kasahara, K.; Yamashita, T.; Sasaki, N.; Yodoi, K.; Matsumoto, T.; Emoto, T.; Hayashi, T.; Kitano, N.; Yoshida, N.; et al. Oral administration of the lactic acid bacterium Pediococcus acidilactici attenuates atherosclerosis in mice by inducing tolerogenic dendritic cells. Heart Vessel. 2017, 32, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, H.; Zhao, Z.; Xiao, S.; Zhao, Y.; Duan, C.; Gao, L.; Li, S.; Wang, J. Pediococcus acidilactici AS185 attenuates early atherosclerosis development through inhibition of lipid regulation and inflammation in rats. J. Funct. Foods 2019, 60, 103424. [Google Scholar] [CrossRef]
- Linares, D.M.; O’Callaghan, T.F.; O’Connor, P.M.; Ross, R.P.; Stanton, C. Streptococcus thermophilus APC151 Strain Is Suitable for the Manufacture of Naturally GABA-Enriched Bioactive Yogurt. Front. Microbiol. 2016, 7, 1876. [Google Scholar] [CrossRef]
- Vendramin, V.; Treu, L.; Campanaro, S.; Lombardi, A.; Corich, V.; Giacomini, A. Genome comparison and physiological characterization of eight Streptococcus thermophilus strains isolated from Italian dairy products. Food Microbiol. 2017, 63, 47–57. [Google Scholar] [CrossRef]
- Klein, B.G. Cunningham’s Textbook of Veterinary Physiology-E-Book; Elsevier Health Sciences: Berlin, Germany, 2013. [Google Scholar]
- Fish, J.; Devries, T. Short communication: Varying dietary dry matter concentration through water addition: Effect on nutrient intake and sorting of dairy cows in late lactation. J. Dairy Sci. 2012, 95, 850–855. [Google Scholar] [CrossRef]
- Lai, C.Y.; Fatimah, A.B.; Mahyudin, N.A.; Saari, N.; Zaman, M.Z. Physico-chemical and microbiological qualities of locally produced raw goat milk. Int. Food Res. J. 2016, 23, 739–750. [Google Scholar]
- Bergamo, P.; Fedele, E.; Iannibelli, L.; Marzillo, G. Fat-soluble vitamin contents and fatty acid composition in organic and conventional Italian dairy products. Food Chem. 2003, 82, 625–631. [Google Scholar] [CrossRef]
- Muehlhoff, E.; Bennett, A.; McMahon, D. Milk and Dairy Products in Human Nutrition; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar]
- Jafari, M.; Rezaei, M.; Gheisari, H.R.; Abhari, K.; Khaniki, G.J.; Noori, N.; Khaneghah, A.M. Application of cultivable lactic acid bacteria isolated from Iranian traditional dairy products for the production of liquid and dried kashks. LWT 2019, 116, 108519. [Google Scholar] [CrossRef]
- González-Martín, I.; Hernández-Hierro, J.M.; Revilla, I.; Vivar-Quintana, A.; Lobos-Ortega, I.; González-Pérez, C. Changes in the Mineral Content in Cheeses of Different Compositions during 6 Months of Ripening. Czech J. Food Sci. 2009, 27, S114–S118. [Google Scholar] [CrossRef]
- Levkov, V.; Stafilov, T.; Pacinovski, N.; Andonovska, K.B.; Mateva, N.; Gjorgovska, N.; Eftimova, E.; Kostadinov, T. The Content of Macro and Trace Elements in Curd and Traditional White Brined Cheese. Veterinarija ir Zootechnika 2017, 75, 36–42. [Google Scholar]
- Milind, P.; Jyoti, M. Curd: A sedative with a bonus bowl of useful side effect. Int. Res. J. Pharm. 2014, 5, 131–135. [Google Scholar]
- Tang, M.; Odejinmi, S.; Vankayalapati, H.; Wierenga, K.; Lai, K. Innovative therapy for Classic Galactosemia—Tale of two HTS. Mol. Genet. Metab. 2012, 105, 44–55. [Google Scholar] [CrossRef]
- McCorvie, T.J.; Timson, D.J. The structural and molecular biology of type I galactosemia: Enzymology of galactose 1-phosphate uridylyltransferase. IUBMB Life 2011, 63, 694–700. [Google Scholar] [CrossRef]
- Tarrah, A.; Noal, V.; Treu, L.; Giaretta, S.; Duarte, V.D.S.; Corich, V.; Giacomini, A. Short communication: Comparison of growth kinetics at different temperatures of Streptococcus macedonicus and Streptococcus thermophilus strains of dairy origin. J. Dairy Sci. 2018, 101, 7812–7816. [Google Scholar] [CrossRef]
- Tarrah, A.; Noal, V.; Giaretta, S.; Treu, L.; Duarte, V.D.S.; Corich, V.; Giacomini, A. Effect of different initial pH on the growth of Streptococcus macedonicus and Streptococcus thermophilus strains. Int. Dairy J. 2018, 86, 65–68. [Google Scholar] [CrossRef]
- Tarrah, A.; Treu, L.; Giaretta, S.; Duarte, V.D.S.; Corich, V.; Giacomini, A. Differences in Carbohydrates Utilization and Antibiotic Resistance Between Streptococcus macedonicus and Streptococcus thermophilus Strains Isolated from Dairy Products in Italy. Curr. Microbiol. 2018, 75, 1334–1344. [Google Scholar] [CrossRef] [PubMed]
- Tarrah, A.; Pakroo, S.; Junior, W.J.F.L.; Guerra, A.F.; Corich, V.; Giacomini, A. Complete Genome Sequence and Carbohydrates-Active EnZymes (CAZymes) Analysis of Lactobacillus paracasei DTA72, a Potential Probiotic Strain with Strong Capability to Use Inulin. Curr. Microbiol. 2020, 77, 2867–2875. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ca | K | Mg | Mn | Na | P | Zn | |
|---|---|---|---|---|---|---|---|
| IRZ1 | 2414.5 ± 78.5 | 8062.3 ± 129.2 | 1706 ± 8.3 | 27.6 ± 0.1 | 14,277.9 ± 150.5 | 3544.4 ± 27.4 | 27.4 ± 0.1 |
| IRZ2 | 1627.1 ± 42 | 9082.2 ± 219.7 | 1824.1 ± 49.5 | 33.6 ± 1 | 18,235.1 ± 569.1 | 3938.2 ± 147.1 | 26.7 ± 0.5 |
| IRZ3 | 2256.9 ± 59.5 | 7961.4 ± 45.1 | 1565.8 ± 33.4 | 22.4 ± 0.2 | 21,761.1 ± 280.6 | 3838.9 ± 51.5 | 25.1 ± 0.4 |
| IRZ4 | 1709.6 ± 79.3 | 8581.8 ± 265.8 | 1690 ± 57.5 | 30.1 ± 1.4 | 24,106.6 ± 1051.1 | 3789.4 ± 153.8 | 19.6 ± 0.8 |
| IRZ5 | 1404.6 ± 40.9 | 7244.9 ± 73.1 | 1658 ± 12.3 | 24.7 ± 0.2 | 17,747.6 ± 187.1 | 4304.7 ± 38.5 | 32.7 ± 0.3 |
| IRZ6 | 1368.4 ± 43.7 | 7387.5 ± 138.2 | 1524.7 ± 42.8 | 27.4 ± 0.4 | 30,069.1 ± 734.7 | 3520.3 ± 87.8 | 32.4 ± 0.7 |
| IRZ7 | 1965.5 ± 31.3 | 7675.4 ± 169.5 | 1594.6 ± 45.9 | 26.3 ± 0.8 | 22,176.2 ± 846.1 | 4500.7 ± 156.1 | 28.7 ± 0.7 |
| IRZ8 | 1359.1 ± 13.6 | 7923.2 ± 41.6 | 1835.7 ± 6.9 | 27.8 ± 0.3 | 15,896.9 ± 67.1 | 4633.5 ± 34.5 | 28.7 ± 0.2 |
| IRZ9 | 2280.1 ± 133.4 | 8703.7 ± 402.5 | 1582.6 ± 83.4 | 25.3 ± 1.4 | 19,659.3 ± 890.3 | 4204.5 ± 225.7 | 26.9 ± 1.5 |
| IRZ10 | 1476.1 ± 73 | 7379.1 ± 48.6 | 1666.8 ± 72.2 | 33.8 ± 0.7 | 10,168.6 ± 69.7 | 4297.6 ± 47.6 | 24.3 ± 1.1 |
| IRZ11 | 1860.1 ± 38.9 | 7554.7 ± 167 | 1660.1 ± 58.2 | 30.9 ± 0.7 | 8911.9 ± 240.4 | 4400.6 ± 119.7 | 28.6 ± 0.8 |
| IRZ12 | 3035.0 ± 107.7 | 9689.4 ± 280.6 | 1770.3 ± 42.6 | 33.7 ± 0.9 | 23,286.5 ± 376.7 | 3939.3 ± 92.8 | 28.2 ± 0.4 |
| IRZ13 | 1894.7 ± 67.9 | 7253.5 ± 134.2 | 1446.4 ± 42.3 | 31.1 ± 0.9 | 22,817.1 ± 309.2 | 3862.7 ± 154.6 | 29.5 ± 0.7 |
| Mean | 1853.1 ± 490.9 | 8038.4 ± 766.1 | 1655.8 ± 114.0 | 28.8 ± 3.7 | 19,162.6 ± 5637.7 | 4059.6 ± 356.4 | 27.6 ± 3.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pakroo, S.; Tarrah, A.; da Silva Duarte, V.; Corich, V.; Giacomini, A. Microbial Diversity and Nutritional Properties of Persian “Yellow Curd” (Kashk zard), a Promising Functional Fermented Food. Microorganisms 2020, 8, 1658. https://doi.org/10.3390/microorganisms8111658
Pakroo S, Tarrah A, da Silva Duarte V, Corich V, Giacomini A. Microbial Diversity and Nutritional Properties of Persian “Yellow Curd” (Kashk zard), a Promising Functional Fermented Food. Microorganisms. 2020; 8(11):1658. https://doi.org/10.3390/microorganisms8111658
Chicago/Turabian StylePakroo, Shadi, Armin Tarrah, Vinícius da Silva Duarte, Viviana Corich, and Alessio Giacomini. 2020. "Microbial Diversity and Nutritional Properties of Persian “Yellow Curd” (Kashk zard), a Promising Functional Fermented Food" Microorganisms 8, no. 11: 1658. https://doi.org/10.3390/microorganisms8111658
APA StylePakroo, S., Tarrah, A., da Silva Duarte, V., Corich, V., & Giacomini, A. (2020). Microbial Diversity and Nutritional Properties of Persian “Yellow Curd” (Kashk zard), a Promising Functional Fermented Food. Microorganisms, 8(11), 1658. https://doi.org/10.3390/microorganisms8111658