Involvement of MexS and MexEF-OprN in Resistance to Toxic Ion Chelators in Pseudomonas putida KT2440

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Media
2.2. Colony Morphology Assay
2.3. Generation of Mutants and Complemented Strains
2.4. Growth Curves
2.5. Luciferase Activity Assay
2.6. Susceptibility Testing by Diffusion Method
2.7. Genomic Sequence Analysis
2.8. Statistical Analysis
3. Results
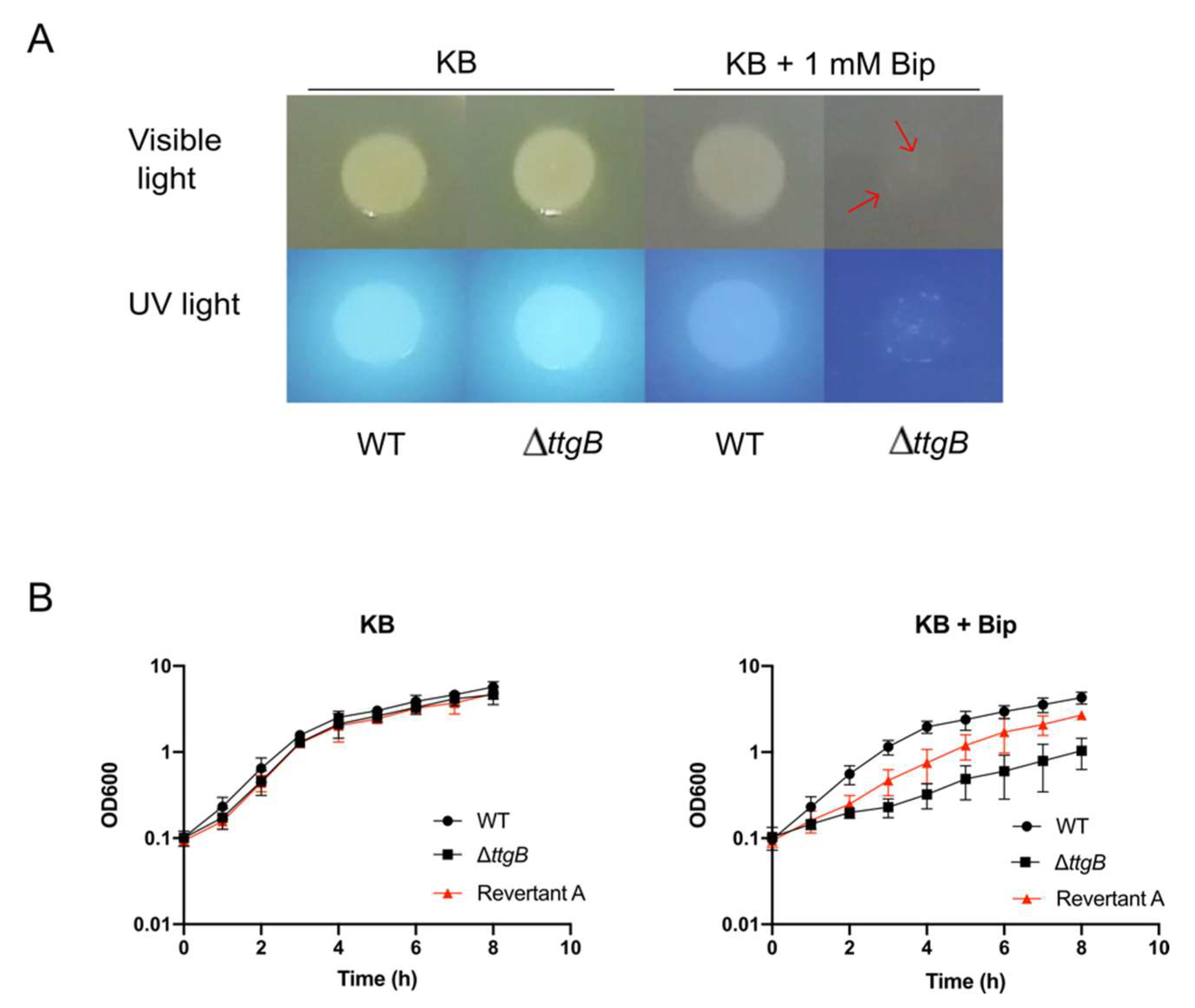
3.1. Bip-Driven Generation of Second Site Revertant Strains from the ttgB Mutant
3.2. Alteration in pp_2827 Rescues Growth of the ttgB Mutant in Presence of Bip
3.3. Alterations in pp_2827 Confer Bip Resistance by Stimulating Expression of mexEF-oprN
3.4. The mexEF-oprN Operon is also Upregulated in Revertants B and C
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Timmis, K.N. Pseudomonas putida: A cosmopolitan opportunist par excellence. Environ. Microbiol. 2002, 4, 779–781. [Google Scholar] [CrossRef] [PubMed]
- Palleroni, N.J. Pseudomonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Dreier, J.; Ruggerone, P. Interaction of antibacterial compounds with RND efflux pumps in Pseudomonas aeruginosa. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, K. Stress responses as determinants of antimicrobial resistance in Pseudomonas aeruginosa: Multidrug efflux and more. Can. J. Microbiol. 2014, 60, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Sobel, M.L.; Poole, K. Antibiotic inducibility of the MexXY multidrug efflux system of Pseudomonas aeruginosa: Involvement of the antibiotic-inducible PA5471 gene product. J. Bacteriol. 2006, 188, 1847–1855. [Google Scholar] [CrossRef] [Green Version]
- Fraud, S.; Campigotto, A.J.; Chen, Z.; Poole, K. MexCD-OprJ multidrug efflux system of Pseudomonas aeruginosa: Involvement in chlorhexidine resistance and induction by membrane-damaging agents dependent upon the AlgU stress response sigma factor. Antimicrob. Agents Chemother. 2008, 52, 4478–4482. [Google Scholar] [CrossRef] [Green Version]
- Köhler, T.; van Delden, C.; Curty, L.K.; Hamzehpour, M.M.; Pechere, J.C. Overexpression of the MexEF-OprN multidrug efflux system affects cell-to-cell signaling in Pseudomonas aeruginosa. J. Bacteriol. 2001, 183, 5213–5222. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, H.; Hosaka, M.; Hirai, K.; Iyobe, S. New norfloxacin resistance gene in Pseudomonas aeruginosa PAO. Antimicrob. Agents Chemother. 1990, 34, 1757–1761. [Google Scholar] [CrossRef] [Green Version]
- Fargier, E.; Mac Aogáin, M.; Mooij, M.J.; Woods, D.F.; Morrissey, J.P.; Dobson, A.D.; Adams, C.; O’Gara, F. MexT functions as a redox-responsive regulator modulating disulfide stress resistance in Pseudomonas aeruginosa. J. Bacteriol. 2012, 194, 3502–3511. [Google Scholar] [CrossRef] [Green Version]
- Westfall, L.W.; Carty, N.L.; Layland, N.; Kuan, P.; Colmer-Hamood, J.A.; Hamood, A.N. mvaT mutation modifies the expression of the Pseudomonas aeruginosa multidrug efflux operon mexEF-oprN. FEMS Microbiol. Lett. 2006, 255, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, D.; Schneper, L.; Merighi, M.; Smith, R.; Narasimhan, G.; Lory, S.; Mathee, K. The regulatory repertoire of Pseudomonas aeruginosa AmpC ß-lactamase regulator AmpR includes virulence genes. PLoS ONE 2012, 7, e34067. [Google Scholar] [CrossRef] [Green Version]
- Richardot, C.; Juarez, P.; Jeannot, K.; Patry, I.; Plésiat, P.; Llanes, C. Amino acid substitutions account for most MexS alterations in clinical nfxC mutants of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2016, 60, 2302–2310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaoui, C.; Overhage, J.; Löns, D.; Zimmermann, A.; Müsken, M.; Bielecki, P.; Pustelny, C.; Becker, T.; Nimtz, M.; Häussler, S. An orphan sensor kinase controls quinolone signal production via MexT in Pseudomonas aeruginosa. Mol. Microbiol. 2012, 83, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Funk, A.; Divekar, P.V. Caerulomycin, a new antibiotic from Streptomyces caeruleus Baldacci. I. Production, isolation, assay, and biological properties. Can. J. Microbiol. 1959, 5, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Henríquez, T.; Stein, N.V.; Jung, H. Resistance to Bipyridyls Mediated by the TtgABC Efflux System in Pseudomonas putida KT2440. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Bagdasarian, M.; Lurz, R.; Rückert, B.; Franklin, F.C.H.; Bagdasarian, M.M.; Frey, J.; Timmis, K.N. Specific-purpose plasmid cloning vectors II. Broad host range, high copy number, RSF 1010-derived vectors, and a host-vector system for gene cloning in Pseudomonas. Gene 1981, 16, 237–247. [Google Scholar] [CrossRef]
- Cronin, C.N.; McIntire, W.S. pUCP-Nco and pUCP-Nde: Escherichia-Pseudomonas shuttle vectors for recombinant protein expression in Pseudomonas. Anal. Biochem. 1999, 272, 112–115. [Google Scholar] [CrossRef]
- Silva-Rocha, R.; Martinez-Garcia, E.; Calles, B.; Chavarria, M.; Arce-Rodriguez, A.; de Las Heras, A.; Paez-Espino, A.D.; Durante-Rodriguez, G.; Kim, J.; Nikel, P.I.; et al. The Standard European Vector Architecture (SEVA): A coherent platform for the analysis and deployment of complex prokaryotic phenotypes. Nucleic Acids Res. 2013, 41, D666–D675. [Google Scholar] [CrossRef]
- Gödeke, J.; Heun, M.; Bubendorfer, S.; Paul, K.; Thormann, K.M. Roles of two Shewanella oneidensis MR-1 extracellular endonucleases. Appl. Environ. Microbiol. 2011, 77, 5342–5351. [Google Scholar] [CrossRef] [Green Version]
- Sakhtah, H.; Koyama, L.; Zhang, Y.; Morales, D.K.; Fields, B.L.; Price-Whelan, A.; Hogan, D.A.; Shepard, K.; Dietrich, L.E.P. The Pseudomonas aeruginosa efflux pump MexGHI-OpmD transports a natural phenazine that controls gene expression and biofilm development. Proc. Natl. Acad. Sci. USA 2016, 113, E3538–E3547. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassak, J.; Henche, A.-L.; Binnenkade, L.; Thormann, K.M. ArcS, the cognate sensor kinase in an atypical Arc system of Shewanella oneidensis MR-1. Appl. Environ. Microbiol. 2010, 76, 3263–3274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winsor, G.L.; Griffiths, E.J.; Lo, R.; Dhillon, B.K.; Shay, J.A.; Brinkman, F.S. Enhanced annotations and features for comparing thousands of Pseudomonas genomes in the Pseudomonas genome database. Nucleic Acids Res. 2016, 44, D646–D653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, K.E.; Weinel, C.; Paulsen, I.T.; Dodson, R.J.; Hilbert, H.; Martins dos Santos, V.A.; Fouts, D.E.; Gill, S.R.; Pop, M.; Holmes, M.; et al. Complete genome sequence and comparative analysis of the metabolically versatile Pseudomonas putida KT2440. Environ. Microbiol. 2002, 4, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Monchy, S.; Taghavi, S.; Zhu, W.; Ramos, J.; van der Lelie, D. Comparative genomics and functional analysis of niche-specific adaptation in Pseudomonas Putida. FEMS Microbiol. Rev. 2011, 35, 299–323. [Google Scholar] [CrossRef] [Green Version]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef]
- Jin, Y.; Yang, H.; Qiao, M.; Jin, S. MexT regulates the type III secretion system through MexS and PtrC in Pseudomonas aeruginosa. J. Bacteriol. 2011, 193, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Sobel, M.L.; Neshat, S.; Poole, K. Mutations in PA2491 (mexS) promote MexT-dependent mexEF-oprN expression and multidrug resistance in a clinical strain of Pseudomonas aeruginosa. J. Bacteriol. 2005, 187, 1246–1253. [Google Scholar] [CrossRef] [Green Version]
- Köhler, T.; Michéa-Hamzehpour, M.; Henze, U.; Gotoh, N.; Curty, L.K.; Pechère, J.C. Characterization of MexE-MexF-OprN, a positively regulated multidrug efflux system of Pseudomonas aeruginosa. Mol. Microbiol. 1997, 23, 345–354. [Google Scholar] [CrossRef]
- Fetar, H.; Gilmour, C.; Klinoski, R.; Daigle, D.M.; Dean, C.R.; Poole, K. mexEF-oprN multidrug efflux operon of Pseudomonas aeruginosa: Regulation by the MexT activator in response to nitrosative stress and chloramphenicol. Antimicrob. Agents Chemother. 2011, 55, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Köhler, T.; Epp, S.F.; Curty, L.K.; Pechère, J.C. Characterization of MexT, the regulator of the MexE-MexF-OprN multidrug efflux system of Pseudomonas aeruginosa. J. Bacteriol. 1999, 181, 6300–6305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrangsu, P.; Rensing, C.; Helmann, J.D. Metal homeostasis and resistance in bacteria. Nat. Rev. Microbiol. 2017, 15, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Herrera, M.C.; Duque, E.; Rodríguez-Herva, J.J.; Fernández-Escamilla, A.M.; Ramos, J.L. Identification and characterization of the PhhR regulon in Pseudomonas putida. Environ. Microbiol. 2010, 12, 1427–1438. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Tomida, J.; Kawamura, Y. Efflux-mediated fluoroquinolone resistance in the multidrug-resistant Pseudomonas aeruginosa clinical isolate PA7: Identification of a novel MexS variant involved in upregulation of the mexEF-oprN multidrug efflux operon. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Uwate, M.; Ichise, Y.K.; Shirai, A.; Omasa, T.; Nakae, T.; Maseda, H. Two routes of MexS-MexT-mediated regulation of MexEF-OprN and MexAB-OprM efflux pump expression in Pseudomonas aeruginosa. Microbiol. Immunol. 2013, 57, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Mooij, M.J.; O’Connor, H.F.; Tian, Z.X.; Wang, Y.P.; Adams, C.; Morrissey, J.P.; O’Gara, F. Antibiotic selection leads to inadvertent selection of nfxC-type phenotypic mutants in Pseudomonas aeruginosa. Environ. Microbiol. Rep. 2010, 2, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Gomi, S.; Amano, S.; Sato, E.; Miyadoh, S.; Kodama, Y. Novel antibiotics SF2738A, B and C, and their analogs produced by Streptomyces sp. J. Antibiot. 1994, 47, 1385–1394. [Google Scholar] [CrossRef]
- Kivistik, P.A.; Kivisaar, M.; Hõrak, R. Target Site Selection of Pseudomonas putida Transposon Tn4652. J. Bacteriol. 2007, 189, 3918. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name (Strains) | Description | Source |
|---|---|---|
| Wild type (WT) | Pseudomonas putida KT2440 | [17] |
| ΔttgB | Derived from strain KT2440 by deletion of pp_1385 | [15] |
| Revertant A, B and C | Spontaneous second site revertants of ΔttgB strain | This work |
| ΔttgB Δpp_2827 | Derived from the ΔttgB strain by deletion of pp_2827 | This work |
| Δpp_2827 | Derived from strain KT2440 by deletion of pp_2827 | This work |
| Revertant A ΔmexF | Derived from Revertant A strain by deletion of pp_3426 | This work |
| ΔttgB Δpp_2827 ΔmexF | Derived from the ΔttgB Δpp_2827 strain by deletion of pp_3426 | This work |
| Name (plasmid) | Description | Source |
| pUCP (pUCP-Nde) | pUCP-NdeI (AmpR) shuttle vector | [18] |
| pUCP-ttgB | Derived from pUCP by cloning the ttgB gene from wild type strain into the multicloning site | [15] |
| pUCP-pp_2827 | Derived from pUCP by cloning pp_2827 from wild type strain into the multicloning site | This work |
| pSEVA224 | KmR; pSEVA221 derivative with lacIq/Ptrc expression system | [19] |
| pSEVA224-mexF | pSEVA224 derivative with pp_3426 cloned into the multicloning site | This work |
| pBBR1-MCS5-lux | pBBR1-based plasmid containing promoter-less luxCDABE, and the aacC1 gene (GenR) | [20] |
| pBBR1-MCS5-Pmex::lux | pBBR1-MCS5-lux derivative containing PmexE::luxCDABE | This work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henriquez, T.; Baldow, T.; Lo, Y.K.; Weydert, D.; Brachmann, A.; Jung, H. Involvement of MexS and MexEF-OprN in Resistance to Toxic Ion Chelators in Pseudomonas putida KT2440. Microorganisms 2020, 8, 1782. https://doi.org/10.3390/microorganisms8111782
Henriquez T, Baldow T, Lo YK, Weydert D, Brachmann A, Jung H. Involvement of MexS and MexEF-OprN in Resistance to Toxic Ion Chelators in Pseudomonas putida KT2440. Microorganisms. 2020; 8(11):1782. https://doi.org/10.3390/microorganisms8111782
Chicago/Turabian StyleHenriquez, Tania, Tom Baldow, Yat Kei Lo, Dina Weydert, Andreas Brachmann, and Heinrich Jung. 2020. "Involvement of MexS and MexEF-OprN in Resistance to Toxic Ion Chelators in Pseudomonas putida KT2440" Microorganisms 8, no. 11: 1782. https://doi.org/10.3390/microorganisms8111782