The Bacillus anthracis Cell Envelope: Composition, Physiological Role, and Clinical Relevance


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
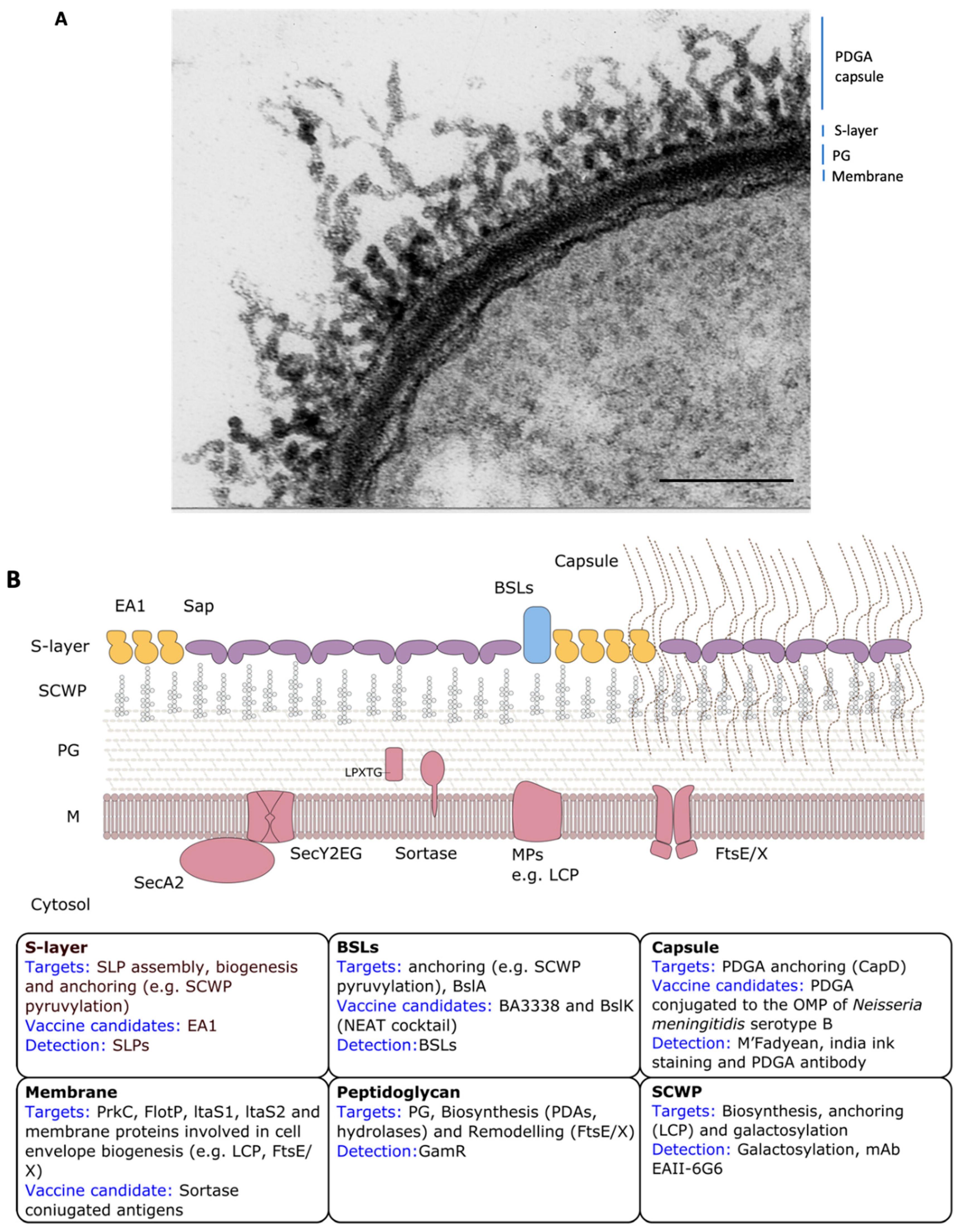
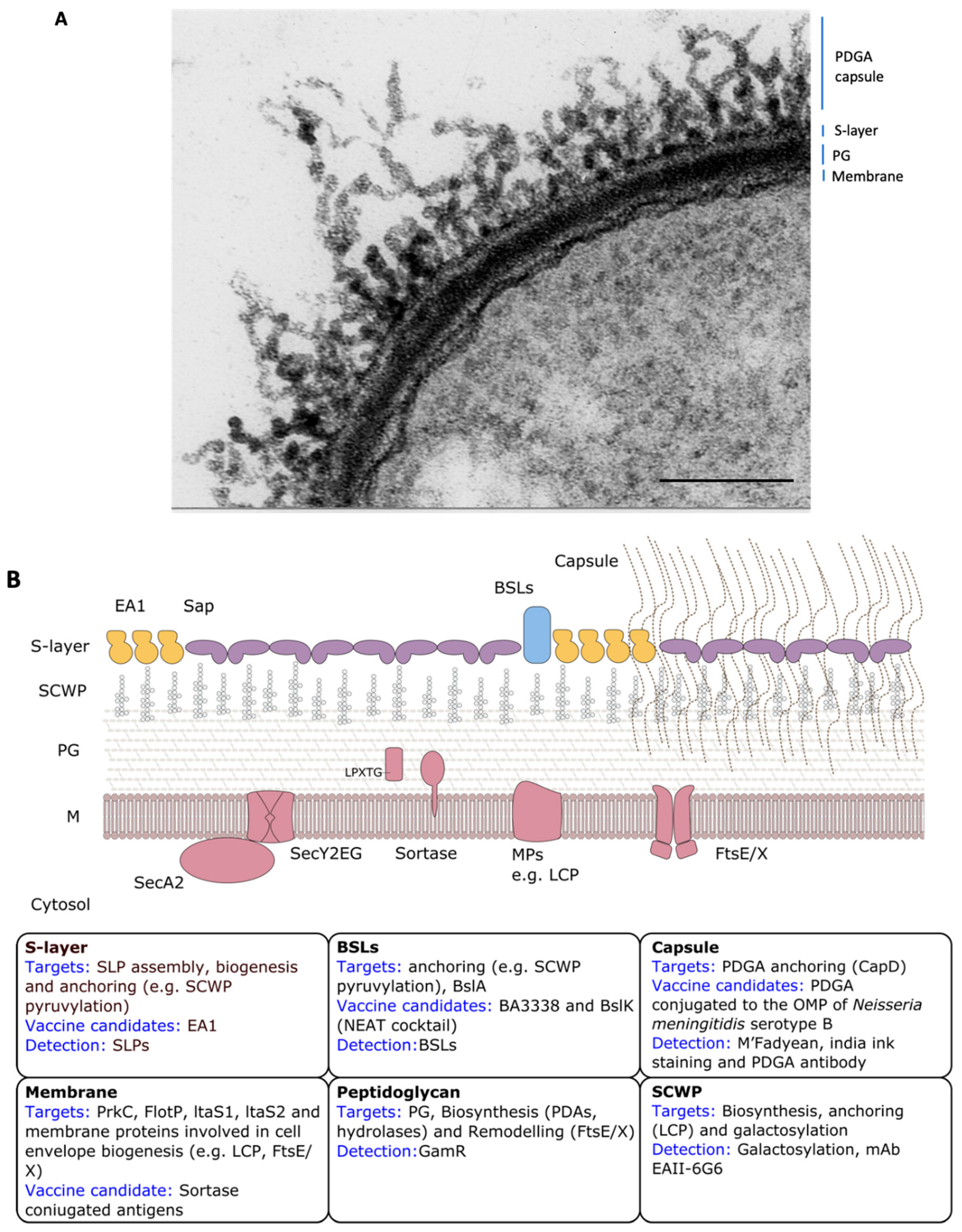
Bacillus anthracis Cell Envelope
2. The Boundary that Enables Life: The Cytoplasmic Membrane
Membrane Proteins
3. The Bacterial Great Wall: The Peptidoglycan Layer
3.1. Cell-Wall-Associated Glycopolymers
3.2. Secondary Cell-Wall Polysaccharide (SCWP)
3.3. Peptidoglycan Covalently Anchored Proteins
4. The Proteinaceous Armor: The S-layer
4.1. Surface-Localized Proteins
4.2. S-layer Anchoring
5. The Invisibility Cloak: The Capsule
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Hueffer, K.; Drown, D.M.; Romanovsky, V.; Hennessy, T. Factors Contributing to Anthrax Outbreaks in the Circumpolar North. EcoHealth 2020, 17, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Zimmermann, F.; Biek, R.; Kuehl, H.; Nowak, K.; Mundry, R.; Agbor, A.; Angedakin, S.; Arandjelovic, M.; Blankenburg, A.; et al. Persistent anthrax as a major driver of wildlife mortality in a tropical rainforest. Nat. Cell Biol. 2017, 548, 82–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukarati, N.L.; Matope, G.; De Garine-Wichatitsky, M.; Ndhlovu, D.N.; Caron, A.; Pfukenyi, D.M. The pattern of anthrax at the wildlife-livestock-human interface in Zimbabwe. PLoS Negl. Trop. Dis. 2020, 14, e0008800. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.J.; Kracalik, I.T.; Ross, N.; Alexander, K.A.; Hugh-Jones, M.E.; Fegan, M.; Elkin, B.T.; Epp, T.; Shury, T.; Zhang, W.; et al. The global distribution of Bacillus anthracis and associated anthrax risk to humans, livestock and wildlife. Nat. Microbiol. 2019, 4, 1337–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonation, K.S.; Grützmacher, K.; Dupke, S.; Mabon, P.; Zimmermann, F.; Lankester, F.; Peller, T.; Feistner, A.; Todd, A.; Herbinger, I.; et al. Bacillus cereus Biovar Anthracis Causing Anthrax in Sub-Saharan Africa—Chromosomal Monophyly and Broad Geographic Distribution. PLoS Negl. Trop. Dis. 2016, 10, e0004923. [Google Scholar] [CrossRef] [PubMed]
- Leendertz, F.H.; Ellerbrok, H.; Boesch, C.; Couacy-Hymann, E.; Mätz-Rensing, K.; Hakenbeck, R.; Bergmann, C.; Abaza, P.; Junglen, S.; Moebius, Y.; et al. Anthrax kills wild chimpanzees in a tropical rainforest. Nat. Cell Biol. 2004, 430, 451–452. [Google Scholar] [CrossRef] [PubMed]
- Hoffmaster, A.R.; Hill, K.K.; Gee, J.E.; Marston, C.K.; De, B.K.; Popovic, T.; Sue, D.; Wilkins, P.P.; Avashia, S.B.; Drumgoole, R.; et al. Characterization of Bacillus cereus Isolates Associated with Fatal Pneumonias: Strains Are Closely Related to Bacillus anthracis and Harbor B. anthracis Virulence Genes. J. Clin. Microbiol. 2006, 44, 3352–3360. [Google Scholar] [CrossRef] [Green Version]
- Kamal, S.M.; Rashid, A.K.M.M.; A Bakar, M.; A Ahad, M. Anthrax: An update. Asian Pac. J. Trop. Biomed. 2011, 1, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Inglesby, T.V.; O’Toole, T.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Friedlander, A.M.; Gerberding, J.; Hauer, J.; Hughes, J.; et al. Anthrax as a Biological Weapon, 2002. JAMA 2002, 287, 2236–2252. [Google Scholar] [CrossRef]
- Kummerfeldt, C.E. Raxibacumab: Potential role in the treatment of inhalational anthrax. Infect. Drug Resist. 2014, 7, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Huang, E.; Pillai, S.K.; Bower, W.A.; Hendricks, K.A.; Guarnizo, J.T.; Hoyle, J.D.; Gorman, S.E.; Boyer, A.E.; Quinn, C.P.; Meaney-Delman, D. Antitoxin Treatment of Inhalation Anthrax: A Systematic Review. Heal. Secur. 2015, 13, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goel, A.K. Anthrax: A disease of biowarfare and public health importance. World J. Clin. Cases 2015, 3, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, M. Dr. Jekyll and Mr. Hyde: A short history of anthrax. Mol. Asp. Med. 2009, 30, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Splino, M.; Patocka, J.; Prymula, R.; Chlibek, R. Anthrax Vaccines. Ann. Saudi Med. 2005, 25, 143–149. [Google Scholar] [CrossRef]
- Whiting, G.; Rijpkema, S.; Adams, T.; Corbel, M. Characterisation of adsorbed anthrax vaccine by two-dimensional gel electrophoresis. Vaccine 2004, 22, 4245–4251. [Google Scholar] [CrossRef]
- Collier, R.J.; Young, J.A. Anthrax Toxin. Annu. Rev. Cell Dev. Biol. 2003, 19, 45–70. [Google Scholar] [CrossRef]
- Hopkins, R.J.; Daczkowski, N.F.; Kaptur, P.E.; Muse, D.; Sheldon, E.; LaForce, C.; Sari, S.; Rudge, T.L.; Bernton, E. Randomized, double-blind, placebo-controlled, safety and immunogenicity study of 4 formulations of Anthrax Vaccine Adsorbed plus CPG 7909 (AV7909) in healthy adult volunteers. Vaccine 2013, 31, 3051–3058. [Google Scholar] [CrossRef] [Green Version]
- Uchida, M.; Harada, T.; Enkhtuya, J.; Kusumoto, A.; Kobayashi, Y.; Chiba, S.; Shyaka, A.; Kawamoto, K. Protective effect of Bacillus anthracis surface protein EA1 against anthrax in mice. Biochem. Biophys. Res. Commun. 2012, 421, 323–328. [Google Scholar] [CrossRef]
- Fioravanti, A.; Van Hauwermeiren, F.; Van Der Verren, S.E.; Jonckheere, W.; Goncalves, A.; Pardon, E.; Steyaert, J.; De Greve, H.; Lamkanfi, M.; Remaut, H. Structure of S-layer protein Sap reveals a mechanism for therapeutic intervention in anthrax. Nat. Microbiol. 2019, 4, 1805–1814. [Google Scholar] [CrossRef] [PubMed]
- Mesnage, S.; Tosi-Couture, E.; Gounon, P.; Mock, M.; Fouet, A. The Capsule and S-Layer: Two Independent and Yet Compatible Macromolecular Structures in Bacillus anthracis. J. Bacteriol. 1998, 180, 52–58. [Google Scholar] [CrossRef] [Green Version]
- Kaneda, T. Factors affecting the relative ratio of fatty acids in Bacillus cereus. Can. J. Microbiol. 1971, 17, 269–275. [Google Scholar] [CrossRef]
- Kaneda, T. Fatty Acids in the Genus Bacillus II. Similarity in the Fatty Acid Compositions of Bacillus thuringiensis, Bacillus anthracis, and Bacillus cereus1. J. Bacteriol. 1968, 95, 2210–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneda, T. Fatty Acids in the Genus Bacillus I. Iso- and Anteiso-Fatty Acids as Characteristic Constituents of Lipids in 10 Species1. J. Bacteriol. 1967, 93, 894–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnár, J.; Prágai, B. Attempts to detect the presence of teichoic acid in Bacillus anthracis. Acta Microbiol. Acad. Sci. Hung. 1971, 18, 105–108. [Google Scholar] [PubMed]
- Fisher, N.A.; Shetron-Rama, L.; Herring-Palmer, A.; Heffernan, B.; Bergman, N.; Hanna, P.C. The dltABCD Operon of Bacillus anthracis Sterne Is Required for Virulence and Resistance to Peptide, Enzymatic, and Cellular Mediators of Innate Immunity. J. Bacteriol. 2006, 188, 1301–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heaton, M.P.; Neuhaus, F.C. Biosynthesis of D-alanyl-lipoteichoic acid: Cloning, nucleotide sequence, and expression of the Lactobacillus casei gene for the D-alanine-activating enzyme. J. Bacteriol. 1992, 174, 4707–4717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garufi, G.; Hendrickx, A.P.; Beeri, K.; Kern, J.W.; Sharma, A.; Richter, S.G.; Schneewind, O.; Missiakas, D. Synthesis of Lipoteichoic Acids in Bacillus anthracis. J. Bacteriol. 2012, 194, 4312–4321. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-M.; Elmets, C.A.; Tang, D.-C.C.; Li, F.; Yusuf, N. Proteomics Reveals that Proteins Expressed During the Early Stage of Bacillus anthracis Infection Are Potential Targets for the Development of Vaccines and Drugs. Genom. Proteom. Bioinform. 2004, 2, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Barat, B.; Ray, W.K.; Helm, R.F.; Melville, S.B.; Popham, D.L. Membrane Proteomes and Ion Transporters inBacillus anthracisandBacillus subtilisDormant and Germinating Spores. J. Bacteriol. 2019, 201, e00662-18. [Google Scholar] [CrossRef] [Green Version]
- Aucher, W.; Davison, S.; Fouet, A. Characterization of the Sortase Repertoire in Bacillus anthracis. PLoS ONE 2011, 6, e27411. [Google Scholar] [CrossRef]
- Gaspar, A.H.; Marraffini, L.A.; Glass, E.M.; Debord, K.L.; Ton-That, H.; Schneewind, O. Bacillus anthracis Sortase A (SrtA) Anchors LPXTG Motif-Containing Surface Proteins to the Cell Wall Envelope. J. Bacteriol. 2005, 187, 4646–4655. [Google Scholar] [CrossRef] [Green Version]
- Maresso, A.W.; Garufi, G.; Schneewind, O. Bacillus anthracis Secretes Proteins That Mediate Heme Acquisition from Hemoglobin. PLoS Pathog. 2008, 4, e1000132. [Google Scholar] [CrossRef] [Green Version]
- Marraffini, L.A.; Schneewind, O. Sortase C-Mediated Anchoring of BasI to the Cell Wall Envelope of Bacillus anthracis. J. Bacteriol. 2007, 189, 6425–6436. [Google Scholar] [CrossRef] [Green Version]
- Zink, S.D.; Burns, D.L. Importance of srtA and srtB for Growth of Bacillus anthracis in Macrophages. Infect. Immun. 2005, 73, 5222–5228. [Google Scholar] [CrossRef] [Green Version]
- Maresso, A.W.; Chapa, T.J.; Schneewind, O. Surface Protein IsdC and Sortase B Are Required for Heme-Iron Scavenging of Bacillus anthracis. J. Bacteriol. 2006, 188, 8145–8152. [Google Scholar] [CrossRef] [Green Version]
- Weiner, E.M.; Robson, S.; Marohn, M.; Clubb, R.T. The Sortase a Enzyme That Attaches Proteins to the Cell Wall ofBacillus anthracisContains an Unusual Active Site Architecture. J. Biol. Chem. 2010, 285, 23433–23443. [Google Scholar] [CrossRef] [Green Version]
- Chan, A.H.; Yi, S.W.; Terwilliger, A.L.; Maresso, A.W.; Jung, M.E.; Clubb, R.T. Structure of the Bacillus anthracis Sortase A Enzyme Bound to Its Sorting Signal. J. Biol. Chem. 2015, 290, 25461–25474. [Google Scholar] [CrossRef] [Green Version]
- Fouet, A. The surface of Bacillus anthracis. Mol. Asp. Med. 2009, 30, 374–385. [Google Scholar] [CrossRef]
- Marraffini, L.A.; Schneewind, O. Targeting proteins to the cell wall of sporulating Bacillus anthracis. Mol. Microbiol. 2006, 62, 1402–1417. [Google Scholar] [CrossRef]
- Gat, O.; Zaide, G.; Inbar, I.; Grosfeld, H.; Chitlaru, T.; Levy, H.; Shafferman, A. Characterization ofBacillus anthracisiron-regulated surface determinant (Isd) proteins containing NEAT domains. Mol. Microbiol. 2008, 70, 983–999. [Google Scholar] [CrossRef]
- Garufi, G.; Wang, Y.-T.; Oh, S.-Y.; Maier, H.; Missiakas, D.M.; Schneewind, O. Sortase-conjugation generates a capsule vaccine that protects guinea pigs against Bacillus anthracis. Vaccine 2012, 30, 3435–3444. [Google Scholar] [CrossRef] [Green Version]
- Somani, V.K.; Aggarwal, S.; Singh, D.; Prasad, T.; Bhatnagar, R. Identification of Novel Raft Marker Protein, FlotP in Bacillus anthracis. Front. Microbiol. 2016, 7, 169. [Google Scholar] [CrossRef] [PubMed]
- Lingwood, D.; Simons, K. Lipid Rafts as a Membrane-Organizing Principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Hicks, D.A.; Nalivaeva, N.N.; Turner, A.J. Lipid Rafts and Alzheimer’s Disease: Protein-Lipid Interactions and Perturbation of Signaling. Front. Physiol. 2012, 3, 189. [Google Scholar] [CrossRef] [Green Version]
- Michel, V.; Bakovic, M. Lipid rafts in health and disease. Biol. Cell 2007, 99, 129–140. [Google Scholar] [CrossRef]
- Dhasmana, N.; Kumar, N.; Gangwal, A.; Keshavam, C.C.; Singh, L.K.; Sangwan, N.; Nashier, P.; Biswas, S.; Pomerantsev, A.P.; Leppla, S.H.; et al. Bacillus anthracis chain length, a virulence determinant, is regulated by a transmembrane Ser/Thr protein kinase PrkC. bioRxiv 2020. [Google Scholar] [CrossRef]
- Jouvion, G.; Corre, J.-P.; Khun, H.; Moya-Nilges, M.; Roux, P.; Latroche, C.; Tournier, J.-N.; Huerre, M.; Chrétien, F.; Goossens, P.L. Physical Sequestration ofBacillus anthracisin the Pulmonary Capillaries in Terminal Infection. J. Infect. Dis. 2016, 214, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Glomski, I.J.; Corre, J.-P.; Mock, M.; Goossens, P.L. Noncapsulated Toxinogenic Bacillus anthracis Presents a Specific Growth and Dissemination Pattern in Naive and Protective Antigen-Immune Mice. Infect. Immun. 2007, 75, 4754–4761. [Google Scholar] [CrossRef] [Green Version]
- Glomski, I.J.; Piris-Gimenez, A.; Huerre, M.; Mock, M.; Goossens, P.L. Primary Involvement of Pharynx and Peyer’s Patch in Inhalational and Intestinal Anthrax. PLoS Pathog. 2007, 3, e76. [Google Scholar] [CrossRef]
- Shakir, S.M.; Bryant, K.M.; Larabee, J.L.; Hamm, E.E.; Lovchik, J.; Lyons, C.R.; Ballard, J.D. Regulatory Interactions of a Virulence-Associated Serine/Threonine Phosphatase-Kinase Pair in Bacillus anthracis. J. Bacteriol. 2009, 192, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Kern, V.J.; Kern, J.W.; Theriot, J.A.; Schneewind, O.; Missiakas, D. Surface-Layer (S-Layer) Proteins Sap and EA1 Govern the Binding of the S-Layer-Associated Protein BslO at the Cell Septa of Bacillus anthracis. J. Bacteriol. 2012, 194, 3833–3840. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.W.; Errington, J. Bacterial cell division: Assembly, maintenance and disassembly of the Z ring. Nat. Rev. Genet. 2009, 7, 642–653. [Google Scholar] [CrossRef]
- Severin, A.; Tabei, K.; Tomasz, A. The Structure of the Cell Wall Peptidoglycan ofBacillus cereusRSVF1, a Strain Closely Related toBacillus anthracis. Microb. Drug Resist. 2004, 10, 77–82. [Google Scholar] [CrossRef]
- Davis, K.M.; Weiser, J.N. Modifications to the Peptidoglycan Backbone Help Bacteria to Establish Infection. Infect. Immun. 2010, 79, 562–570. [Google Scholar] [CrossRef] [Green Version]
- Vollmer, W. Structural variation in the glycan strands of bacterial peptidoglycan. FEMS Microbiol. Rev. 2008, 32, 287–306. [Google Scholar] [CrossRef] [PubMed]
- Sychantha, D.; Chapman, R.N.; Bamford, N.C.; Boons, G.-J.; Howell, P.L.; Clarke, A.J. Molecular Basis for the Attachment of S-Layer Proteins to the Cell Wall of Bacillus anthracis. Biochem. 2018, 57, 1949–1953. [Google Scholar] [CrossRef] [PubMed]
- Helgason, E.; Økstad, O.A.; Caugant, D.A.; Johansen, H.A.; Fouet, A.; Mock, M.; Hegna, I.; Kolstø, A.-B. Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis—One Species on the Basis of Genetic Evidence. Appl. Environ. Microbiol. 2000, 66, 2627–2630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balomenou, S.; Fouet, A.; Tzanodaskalaki, M.; Couture-Tosi, E.; Bouriotis, V.; Boneca, I.G. Distinct functions of polysaccharide deacetylases in cell shape, neutral polysaccharide synthesis and virulence ofBacillus anthracis. Mol. Microbiol. 2013, 87, 867–883. [Google Scholar] [CrossRef]
- Balomenou, S.; Koutsioulis, D.; Tomatsidou, A.; Tzanodaskalaki, M.; Petratos, K.; Bouriotis, V. Polysaccharide deacetylases serve as new targets for the design of inhibitors against Bacillus anthracis and Bacillus cereus. Bioorg. Med. Chem. 2018, 26, 3845–3851. [Google Scholar] [CrossRef]
- Turnbull, P.C.B.; Sirianni, N.M.; Lebron, C.I.; Samaan, M.N.; Sutton, F.N.; Reyes, A.E.; Peruski, L.F.J. MICs of Selected Antibiotics for Bacillus anthracis, Bacillus cereus, Bacillus thuringiensis, and Bacillus mycoides from a Range of Clinical and Environmental Sources as Determined by the Etest. J. Clin. Microbiol. 2004, 42, 3626–3634. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.J.; Rutala, W.A. Bacillus Species. Infect. Control. Hosp. Epidemiol. 1988, 9, 368–373. [Google Scholar] [CrossRef]
- Chen, Y.; Succi, J.; Tenover, F.C.; Koehler, T.M. β-Lactamase Genes of the Penicillin-Susceptible Bacillus anthracis Sterne Strain. J. Bacteriol. 2003, 185, 823–830. [Google Scholar] [CrossRef] [Green Version]
- Crawford, M.A.; Zhu, Y.; Green, C.S.; Burdick, M.D.; Sanz, P.; Alem, F.; O’Brien, A.D.; Mehrad, B.; Strieter, R.M.; Hughes, M.A. Antimicrobial Effects of Interferon-Inducible CXC Chemokines against Bacillus anthracis Spores and Bacilli. Infect. Immun. 2009, 77, 1664–1678. [Google Scholar] [CrossRef] [Green Version]
- Margulieux, K.R.; Fox, J.W.; Nakamoto, R.K.; Hughes, M.A. CXCL10 Acts as a Bifunctional Antimicrobial Molecule against Bacillus anthracis. mBio 2016, 7, e00334-16. [Google Scholar] [CrossRef] [Green Version]
- Margulieux, K.R.; Liebov, B.K.; Tirumala, V.S.K.K.S.; Singh, A.; Bushweller, J.H.; Nakamoto, R.K.; Hughes, M.A. Bacillus anthracis Peptidoglycan Integrity Is Disrupted by the Chemokine CXCL10 through the FtsE/X Complex. Front. Microbiol. 2017, 8, 740. [Google Scholar] [CrossRef]
- Chan, Y.G.Y.; Frankel, M.B.; Dengler, V.; Schneewind, O.; Missiakas, D. Staphylococcus aureus Mutants Lacking the LytR-CpsA-Psr Family of Enzymes Release Cell Wall Teichoic Acids into the Extracellular Medium. J. Bacteriol. 2013, 195, 4650–4659. [Google Scholar] [CrossRef] [Green Version]
- Kawai, Y.; Marles-Wright, J.; Cleverley, R.M.; Emmins, R.; Ishikawa, S.; Kuwano, M.; Heinz, N.; Bui, N.K.; Hoyland, C.N.; Ogasawara, N.; et al. A widespread family of bacterial cell wall assembly proteins. EMBO J. 2011, 30, 4931–4941. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.G.-Y.; Kim, H.K.; Schneewind, O.; Missiakas, D. The Capsular Polysaccharide ofStaphylococcus aureusIs Attached to Peptidoglycan by the LytR-CpsA-Psr (LCP) Family of Enzymes. J. Biol. Chem. 2014, 289, 15680–15690. [Google Scholar] [CrossRef] [Green Version]
- Rausch, M.; Deisinger, J.P.; Ulm, H.; Müller, A.; Li, W.; Hardt, P.; Wang, X.; Li, X.; Sylvester, M.; Engeser, M.; et al. Coordination of capsule assembly and cell wall biosynthesis in Staphylococcus aureus. Nat. Commun. 2019, 10, 1404. [Google Scholar] [CrossRef]
- Richter, S.; Anderson, V.J.; Garufi, G.; Lu, L.; Budzik, J.M.; Joachimiak, A.; He, C.; Schneewind, O.; Missiakas, D. Capsule anchoring inBacillus anthracisoccurs by a transpeptidation reaction that is inhibited by capsidin. Mol. Microbiol. 2009, 71, 404–420. [Google Scholar] [CrossRef] [Green Version]
- Zilla, M.L.; Lunderberg, J.M.; Schneewind, O.; Missiakas, D. Bacillus anthracis lcpGenes Support Vegetative Growth, Envelope Assembly, and Spore Formation. J. Bacteriol. 2015, 197, 3731–3741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilla, M.L.; Chan, Y.G.Y.; Lunderberg, J.M.; Schneewind, O.; Missiakas, D. LytR-CpsA-Psr Enzymes as Determinants of Bacillus anthracis Secondary Cell Wall Polysaccharide Assembly. J. Bacteriol. 2014, 197, 343–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hübscher, J.; Lüthy, L.; Berger-Bächi, B.; Meier, P.S. Phylogenetic distribution and membrane topology of the LytR-CpsA-Psr protein family. BMC Genom. 2008, 9, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, B.; Leoff, C.; Saile, E.; Wilkins, P.; Quinn, C.P.; Kannenberg, E.L.; Carlson, R.W. The Structure of the Major Cell Wall Polysaccharide ofBacillus anthracisIs Species-specific. J. Biol. Chem. 2006, 281, 27932–27941. [Google Scholar] [CrossRef] [Green Version]
- Forsberg, L.S.; Choudhury, B.; Leoff, C.; Marston, C.K.; Hoffmaster, A.R.; Saile, E.; Quinn, C.P.; Kannenberg, E.L.; Carlson, R.W. Secondary cell wall polysaccharides from Bacillus cereus strains G9241, 03BB87 and 03BB102 causing fatal pneumonia share similar glycosyl structures with the polysaccharides from Bacillus anthracis. Glycobiology 2011, 21, 934–948. [Google Scholar] [CrossRef]
- Leoff, C.; Choudhury, B.; Saile, E.; Quinn, C.P.; Carlson, R.W.; Kannenberg, E.L. Structural Elucidation of the Nonclassical Secondary Cell Wall Polysaccharide fromBacillus cereusATCC 10987. J. Biol. Chem. 2008, 283, 29812–29821. [Google Scholar] [CrossRef] [Green Version]
- Candela, T.; Maes, E.; Garénaux, E.; Rombouts, Y.; Krzewinski, F.; Gohar, M.; Guerardel, Y. Environmental and Biofilm-dependent Changes in aBacillus cereusSecondary Cell Wall Polysaccharide. J. Biol. Chem. 2011, 286, 31250–31262. [Google Scholar] [CrossRef] [Green Version]
- Forsberg, L.S.; Abshire, T.G.; Friedlander, A.; Quinn, C.P.; Kannenberg, E.L.; Carlson, R.W. Localization and structural analysis of a conserved pyruvylated epitope in Bacillus anthracis secondary cell wall polysaccharides and characterization of the galactose-deficient wall polysaccharide from avirulent B. anthracis CDC 684. Glycobiology 2012, 22, 1103–1117. [Google Scholar] [CrossRef]
- Okinaka, R.T.; Price, E.P.; Wolken, S.R.; Gruendike, J.; Chung, W.K.; Pearson, T.; Xie, G.; Emunk, C.; Hill, K.K.; Challacombe, J.; et al. An attenuated strain of Bacillus anthracis (CDC 684) has a large chromosomal inversion and altered growth kinetics. BMC Genom. 2011, 12, 477. [Google Scholar] [CrossRef]
- Chateau, A.; Lunderberg, J.M.; Oh, S.Y.; Abshire, T.; Friedlander, A.; Quinn, C.P.; Missiakas, D.M.; Schneewind, O. Galactosylation of the Secondary Cell Wall Polysaccharide of Bacillus anthracis and Its Contribution to Anthrax Pathogenesis. J. Bacteriol. 2018, 200, e00562-17. [Google Scholar] [CrossRef] [Green Version]
- Chateau, A.; Oh, S.Y.; Tomatsidou, A.; Brockhausen, I.; Schneewind, O.; Missiakas, D. Distinct pathways carry out alpha and beta galactosylation of secondary cell wall polysaccharide in Bacillus anthracis. J. Bacteriol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ezzell, J.W.; Abshire, T.G.; Little, S.F.; Lidgerding, B.C.; Brown, C. Identification of Bacillus anthracis by using monoclonal antibody to cell wall galactose-N-acetylglucosamine polysaccharide. J. Clin. Microbiol. 1990, 28, 223–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De, B.K.; Bragg, S.L.; Sanden, G.N.; Wilson, K.E.; Diem, L.A.; Marston, C.K.; Hoffmaster, A.R.; Barnet, G.A.; Weyant, R.S.; Abshire, T.G.; et al. Two-Component Direct Fluorescent-Antibody Assay for Rapid Identification ofBacillus anthracis. Emerg. Infect. Dis. 2002, 8, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Kamal, N.; Ganguly, J.; Saile, E.; Klee, S.R.; Hoffmaster, A.; Carlson, R.W.; Forsberg, L.S.; Kannenberg, E.L.; Quinn, C.P. Structural and immunochemical relatedness suggests a conserved pathogenicity motif for secondary cell wall polysaccharides in Bacillus anthracis and infection-associated Bacillus cereus. PLoS ONE 2017, 12, e0183115. [Google Scholar] [CrossRef] [Green Version]
- Schuch, R.; Nelson, D.; Fischetti, V.A. A bacteriolytic agent that detects and kills Bacillus anthracis. Nat. Cell Biol. 2002, 418, 884–889. [Google Scholar] [CrossRef]
- Low, L.Y.; Yang, C.; Perego, M.; Osterman, A.; Liddington, R.C. Structure and Lytic Activity of aBacillus anthracisProphage Endolysin. J. Biol. Chem. 2005, 280, 35433–35439. [Google Scholar] [CrossRef] [Green Version]
- Ganguly, J.; Low, L.Y.; Kamal, N.; Saile, E.; Forsberg, L.S.; Gutierrez-Sanchez, G.; Hoffmaster, A.R.; Liddington, R.; Quinn, C.P.; Carlson, R.W.; et al. The secondary cell wall polysaccharide of Bacillus anthracis provides the specific binding ligand for the C-terminal cell wall-binding domain of two phage endolysins, PlyL and PlyG. Glycobiology 2013, 23, 820–832. [Google Scholar] [CrossRef]
- Mo, K.-F.; Li, X.; Li, H.; Low, L.Y.; Quinn, C.P.; Boons, G.-J. Endolysins of Bacillus anthracis Bacteriophages Recognize Unique Carbohydrate Epitopes of Vegetative Cell Wall Polysaccharides with High Affinity and Selectivity. J. Am. Chem. Soc. 2012, 134, 15556–15562. [Google Scholar] [CrossRef] [Green Version]
- Heselpoth, R.D.; Owens, J.M.; Nelson, D.C. Quantitative analysis of the thermal stability of the gamma phage endolysin PlyG: A biophysical and kinetic approach to assaying therapeutic potential. Virology 2015, 477, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Boekhorst, J.; De Been, M.W.H.J.; Kleerebezem, M.; Siezen, R.J. Genome-Wide Detection and Analysis of Cell Wall-Bound Proteins with LPxTG-Like Sorting Motifs. J. Bacteriol. 2005, 187, 4928–4934. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Liang, X.; Chen, Y.; Koehler, T.M.; Höök, M. Identification and Biochemical Characterization of Two Novel Collagen Binding MSCRAMMs ofBacillus anthracis. J. Biol. Chem. 2004, 279, 51760–51768. [Google Scholar] [CrossRef] [Green Version]
- Davison, S.; Couture-Tosi, E.; Candela, T.; Mock, M.; Fouet, A. Identification of the Bacillus anthracis γ Phage Receptor. J. Bacteriol. 2005, 187, 6742–6749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gat, O.; Grosfeld, H.; Ariel, N.; Inbar, I.; Zaide, G.; Broder, Y.; Zvi, A.; Chitlaru, T.; Altboum, Z.; Stein, D.; et al. Search for Bacillus anthracis Potential Vaccine Candidates by a Functional Genomic-Serologic Screen. Infect. Immun. 2006, 74, 3987–4001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albers, S.-V.; Meyer, B.H. The archaeal cell envelope. Nat. Rev. Genet. 2011, 9, 414–426. [Google Scholar] [CrossRef]
- Sleytr, U.B.; Schuster, B.; Egelseer, E.-M.; Pum, D. S-layers: Principles and applications. FEMS Microbiol. Rev. 2014, 38, 823–864. [Google Scholar] [CrossRef] [PubMed]
- Gerbino, E.; Carasi, P.; Mobili, P.; A Serradell, M.; Gomezzavaglia, A. Role of S-layer proteins in bacteria. World J. Microbiol. Biotechnol. 2015, 31, 1877–1887. [Google Scholar] [CrossRef] [PubMed]
- Mignot, T.; Mesnage, S.; Couture-Tosi, E.; Mock, M.; Fouet, A. Developmental switch of S-layer protein synthesis in Bacillus anthracis. Mol. Microbiol. 2002, 43, 1615–1627. [Google Scholar] [CrossRef] [Green Version]
- Nguyen-Mau, S.-M.; Oh, S.-Y.; Kern, V.J.; Missiakas, D.M.; Schneewind, O. Secretion Genes as Determinants of Bacillus anthracis Chain Length. J. Bacteriol. 2012, 194, 3841–3850. [Google Scholar] [CrossRef] [Green Version]
- Lupas, A.; Engelhardt, H.; Peters, J.; Santarius, U.; Volker, S.; Baumeister, W. Domain structure of the Acetogenium kivui surface layer revealed by electron crystallography and sequence analysis. J. Bacteriol. 1994, 176, 1224–1233. [Google Scholar] [CrossRef] [Green Version]
- Kern, J.W.; Wilton, R.; Zhang, R.; Binkowski, T.A.; Joachimiak, A.; Schneewind, O. Structure of Surface Layer Homology (SLH) Domains fromBacillus anthracisSurface Array Protein. J. Biol. Chem. 2011, 286, 26042–26049. [Google Scholar] [CrossRef] [Green Version]
- Blackler, R.J.; López-Guzmán, A.; Hager, F.F.; Janesch, B.; Martinz, G.; Gagnon, S.M.L.; Haji-Ghassemi, O.; Kosma, P.; Messner, P.; Schäffer, C.; et al. Structural basis of cell wall anchoring by SLH domains in Paenibacillus alvei. Nat. Commun. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Baranova, E.; Fronzes, R.; Garcia-Pino, A.; Van Gerven, N.; Papapostolou, D.; Péhau-Arnaudet, G.; Pardon, E.; Steyaert, J.; Howorka, S.; Remaut, H. SbsB structure and lattice reconstruction unveil Ca2+ triggered S-layer assembly. Nat. Cell Biol. 2012, 487, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Couture-Tosi, E.; Delacroix, H.; Mignot, T.; Mesnage, S.; Chami, M.; Fouet, A.; Mosser, G. Structural Analysis and Evidence for Dynamic Emergence of Bacillus anthracis S-Layer Networks. J. Bacteriol. 2002, 184, 6448–6456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, D.; Zhang, Z.; Bi, L.; Zhang, J.; Ding, W.; Zhang, X. A S-Layer Protein of Bacillus anthracis as a Building Block for Functional Protein Arrays by In Vitro Self-Assembly. Small 2015, 11, 5826–5832. [Google Scholar] [CrossRef] [Green Version]
- Rad, B.; Haxton, T.K.; Shon, A.; Shin, S.-H.; Whitelam, S.; Ajo-Franklin, C.M. Ion-Specific Control of the Self-Assembly Dynamics of a Nanostructured Protein Lattice. ACS Nano 2014, 9, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Anderson, V.J.; Kern, J.W.; McCool, J.W.; Schneewind, O.; Missiakas, D. The SLH-domain protein BslO is a determinant of Bacillus anthracis chain length. Mol. Microbiol. 2011, 81, 192–205. [Google Scholar] [CrossRef] [Green Version]
- Mesnage, S.; Tosi-Couture, E.; Mock, M.; Gounon, P.; Fouet, A. Molecular characterization of the Bacillus anthracis main S-layer component: Evidence that it is the major cell-associated antigen. Mol. Microbiol. 1997, 23, 1147–1155. [Google Scholar] [CrossRef]
- Plaut, R.D.; Beaber, J.W.; Zemansky, J.; Kaur, A.P.; George, M.; Biswas, B.; Henry, M.; Bishop-Lilly, K.A.; Mokashi, V.; Hannah, R.M.; et al. Genetic Evidence for the Involvement of the S-Layer Protein Gene sap and the Sporulation Genes spo0A, spo0B, and spo0F in Phage AP50c Infection of Bacillus anthracis. J. Bacteriol. 2014, 196, 1143–1154. [Google Scholar] [CrossRef] [Green Version]
- Baillie, L.; Hebdon, R.; Flick-Smith, H.; Williamson, D. Characterisation of the immune response to the UK human anthrax vaccine. FEMS Immunol. Med. Microbiol. 2003, 36, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Lai, E.-M.; Phadke, N.D.; Kachman, M.T.; Giorno, R.; Vazquez, S.; Vazquez, J.A.; Maddock, J.R.; Driks, A. Proteomic Analysis of the Spore Coats of Bacillus subtilis and Bacillus anthracis. J. Bacteriol. 2003, 185, 1443–1454. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Akmal, A.; Stewart, A.C.; Hsia, R.-C.; Read, T.D. Identification of Bacillus anthracis Spore Component Antigens Conserved across Diverse Bacillus cereus sensu lato Strains. Mol. Cell. Proteom. 2009, 8, 1174–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, D.D.; Turnbough, C.L. Surface Layer Protein EA1 Is Not a Component of Bacillus anthracis Spores but Is a Persistent Contaminant in Spore Preparations. J. Bacteriol. 2004, 186, 566–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, T.E.; Redmond, C.; Mayers, C.N. Real time detection of anthrax spores using highly specific anti-EA1 recombinant antibodies produced by competitive panning. J. Immunol. Methods 2008, 334, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kern, J.W.; Schneewind, O. BslA, a pXO1-encoded adhesin of Bacillus anthracis. Mol. Microbiol. 2008, 68, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Chitlaru, T.; Zaide, G.; Ehrlich, S.; Inbar, I.; Cohen, O.; Shafferman, A. HtrA is a major virulence determinant of Bacillus anthracis. Mol. Microbiol. 2011, 81, 1542–1559. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, D.; Ren, J.; Tong, C.; Feng, E.; Wang, X.; Wang, H.; Wang, H. Identification of the Immunogenic Spore and Vegetative Proteins of Bacillus anthracis Vaccine Strain A16R. PLoS ONE 2013, 8, e57959. [Google Scholar] [CrossRef]
- Tarlovsky, Y.; Fabian, M.; Solomaha, E.; Honsa, E.; Olson, J.S.; Maresso, A.W. A Bacillus anthracis S-Layer Homology Protein That Binds Heme and Mediates Heme Delivery to IsdC. J. Bacteriol. 2010, 192, 3503–3511. [Google Scholar] [CrossRef] [Green Version]
- Balderas, M.A.; Nguyen, C.T.Q.; Terwilliger, A.; Keitel, W.A.; Iniguez, A.; Torres, R.; Palacios, F.; Goulding, C.W.; Maresso, A.W. Progress toward the Development of a NEAT Protein Vaccine for Anthrax Disease. Infect. Immun. 2016, 84, 3408–3422. [Google Scholar] [CrossRef] [Green Version]
- Jelinski, J.; Terwilliger, A.; Green, S.; Maresso, A.W. Progress towards the Development of a NEAT Vaccine for Anthrax II: Immunogen Specificity and Alum Effectiveness in an Inhalational Model. Infect. Immun. 2020, 88. [Google Scholar] [CrossRef]
- Fedhila, S.; Daou, N.; Lereclus, D.; Nielsen-Leroux, C. Identification ofBacillus cereusinternalin and other candidate virulence genes specifically induced during oral infection in insects. Mol. Microbiol. 2006, 62, 339–355. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Puranik, N.; Varshney, A.; Tripathi, N.; Pal, V.; Goel, A.K. BA3338, a surface layer homology domain possessing protein augments immune response and protection efficacy of protective antigen against Bacillus anthracis in mouse model. J. Appl. Microbiol. 2020, 129, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, C.M.; Kern, J.W.; Sheen, T.R.; Ebrahimi-Fardooee, M.A.; Van Sorge, N.M.; Schneewind, O.; Doran, K.S. Penetration of the Blood-Brain Barrier by Bacillus anthracis Requires the pXO1-Encoded BslA Protein. J. Bacteriol. 2009, 191, 7165–7173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glinert, I.; Weiss, S.; Sittner, A.; Bar-David, E.; Ben-Shmuel, A.; Schlomovitz, J.; Kobiler, D.; Levy, H. Infection with a Nonencapsulated Bacillus anthracis Strain in Rabbits—The Role of Bacterial Adhesion and the Potential for a Safe Live Attenuated Vaccine. Toxins 2018, 10, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ries, W.; Hotzy, C.; Schocher, I.; Sleytr, U.B.; Sára, M. Evidence that the N-terminal part of the S-layer protein from Bacillus stearothermophilus PV72/p2 recognizes a secondary cell wall polymer. J. Bacteriol. 1997, 179, 3892–3898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesnage, S.; Fontaine, T.; Mignot, T.; Delepierre, M.; Mock, M.; Fouet, A. Bacterial SLH domain proteins are non-covalently anchored to the cell surface via a conserved mechanism involving wall polysaccharide pyruvylation. EMBO J. 2000, 19, 4473–4484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, R.N.; Liu, L.; Boons, G.-J. 4,6-O-Pyruvyl Ketal Modified N-Acetylmannosamine of the Secondary Cell Wall Polysaccharide of Bacillus anthracis Is the Anchoring Residue for Its Surface Layer Proteins. J. Am. Chem. Soc. 2018, 140, 17079–17085. [Google Scholar] [CrossRef]
- Sychantha, D.; Little, D.J.; Chapman, R.N.; Boons, G.; Robinson, H.; Howell, P.L.; Clarke, A.J. PatB1 is an O-acetyltransferase that decorates secondary cell wall polysaccharides. Nat. Chem. Biol. 2017, 14, 79–85. [Google Scholar] [CrossRef]
- Lunderberg, J.M.; Nguyen-Mau, S.-M.; Richter, G.S.; Wang, Y.-T.; Dworkin, J.; Missiakas, D.M.; Schneewind, O. Bacillus anthracis Acetyltransferases PatA1 and PatA2 Modify the Secondary Cell Wall Polysaccharide and Affect the Assembly of S-Layer Proteins. J. Bacteriol. 2012, 195, 977–989. [Google Scholar] [CrossRef]
- World Health. Guidelines for the Surveillance and Control of Anthrax in Humans and Animals, 3rd ed.; Turnbull, P.C.B., Ed.; World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Bell, C.A.; Uhl, J.R.; Hadfield, T.L.; David, J.C.; Meyer, R.F.; Smith, T.F.; Iii, F.R.C. Detection of Bacillus anthracis DNA by LightCycler PCR. J. Clin. Microbiol. 2002, 40, 2897–2902. [Google Scholar] [CrossRef] [Green Version]
- Jończyk-Matysiak, E.; Kłak, M.; Weber-Dąbrowska, B.; Borysowski, J.; Górski, A. Possible Use of Bacteriophages Active againstBacillus anthracisand OtherB. cereusGroup Members in the Face of a Bioterrorism Threat. BioMed Res. Int. 2014, 2014, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Koper, O.B.; Klabunde, J.S.; Marchin, G.L.; Klabunde, K.J.; Stoimenov, P.; Bohra, L. Nanoscale Powders and Formulations with Biocidal Activity Toward Spores and Vegetative Cells of Bacillus Species, Viruses, and Toxins. Curr. Microbiol. 2002, 44, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Walter, M.H. Efficacy and durability of Bacillus anthracis bacteriophages used against spores. J. Environ. Health 2003, 66, 9. [Google Scholar]
- Brigati, J.; Williams, D.D.; Sorokulova, I.B.; Nanduri, V.; Chen, I.-H.; Turnbough, C.L.; A Petrenko, V. Diagnostic Probes for Bacillus anthracis Spores Selected from a Landscape Phage Library. Clin. Chem. 2004, 50, 1899–1906. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Johnson, M.L.; Guntupalli, R.; Petrenko, V.A.; Chin, B.A. Detection of Bacillus anthracis spores in liquid using phage-based magnetoelastic micro-resonators. Sens. Actuators B Chem. 2007, 127, 559–566. [Google Scholar] [CrossRef]
- Shen, W.; Lakshmanan, R.S.; Mathison, L.C.; Petrenko, V.A.; Chin, B.A. Phage coated magnetoelastic micro-biosensors for real-time detection of Bacillus anthracis spores☆. Sens. Actuators B Chem. 2009, 137, 501–506. [Google Scholar] [CrossRef]
- Fu, L.; Li, S.; Zhang, K.; Chen, I.-H.; Barbaree, J.M.; Zhang, A.; Cheng, Z.-Y. Detection of Bacillus anthracis Spores Using Phage-Immobilized Magnetostrictive Milli/Micro Cantilevers. IEEE Sens. J. 2010, 11, 1684–1691. [Google Scholar] [CrossRef]
- Sutherland, I.W.; Hughes, K.A.; Skillman, L.C.; Tait, K. The interaction of phage and biofilms. FEMS Microbiol. Lett. 2004, 232, 1–6. [Google Scholar] [CrossRef]
- Schofield, D.A.; Sharp, N.J.; Westwater, C. Phage-based platforms for the clinical detection of human bacterial pathogens. Bacteriophage 2012, 2, 105–121. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.R.; Moody, M.D.; Treece, E.L.; Smith, C.W. Differential diagnosis of bacillus cereus, bacillus anthracis, and BACILLUS Cereus var. mycoides. J. Bacteriol. 1958, 75, 499–509. [Google Scholar] [CrossRef] [Green Version]
- Negus, D.; Burton, J.; Sweed, A.; Gryko, R.; Taylor, P.W. Poly-γ-d-Glutamic Acid Capsule Interferes with Lytic Infection of Bacillus anthracis by B. anthracis-Specific Bacteriophages. Appl. Environ. Microbiol. 2012, 79, 714–717. [Google Scholar] [CrossRef] [Green Version]
- Kan, S.; Fornelos, N.; Schuch, R.; Fischetti, V.A. Identification of a Ligand on the Wip1 Bacteriophage Highly Specific for a Receptor on Bacillus anthracis. J. Bacteriol. 2013, 195, 4355–4364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, E.; Ivánovics, G. Association of Probable Defective Phage Particles with Lysis by Bacteriophage AP50 in Bacillus anthracis. J. Gen. Microbiol. 1977, 102, 215–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sozhamannan, S.; McKinstry, M.; Lentz, S.M.; Jalasvuori, M.; McAfee, F.; Smith, A.; Dabbs, J.; Ackermann, H.-W.; Bamford, J.K.H.; Mateczun, A.; et al. Molecular Characterization of a Variant of Bacillus anthracis-Specific Phage AP50 with Improved Bacteriolytic Activity. Appl. Environ. Microbiol. 2008, 74, 6792–6796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop-Lilly, K.A.; Plaut, R.D.; E Chen, P.; Akmal, A.; Willner, K.M.; Butani, A.; Dorsey, S.; Mokashi, V.P.; Mateczun, A.J.; E Chapman, C.; et al. Whole genome sequencing of phage resistant Bacillus anthracis mutants reveals an essential role for cell surface anchoring protein CsaB in phage AP50c adsorption. Virol. J. 2012, 9, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavallo, J.-D.; Ramisse, F.; Girardet, M.; Vaissaire, J.; Mock, M.; Hernandez, E. Antibiotic Susceptibilities of 96 Isolates of Bacillus anthracis Isolated in France between 1994 and 2000. Antimicrob. Agents Chemother. 2002, 46, 2307–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inal, J.M.; Karunakaran, K.V. φ20, A Temperate Bacteriophage Isolated from Bacillus anthracis Exists as a Plasmidial Prophage. Curr. Microbiol. 1996, 32, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Inal, J.M. Phage therapy: A reappraisal of bacteriophages as antibiotics. Arch. Immunol. Ther. Exp. 2003, 51, 237–244. [Google Scholar]
- Brabban, A.D.; Hite, E.; Callaway, T. Evolution of Foodborne Pathogens via Temperate Bacteriophage-Mediated Gene Transfer. Foodborne Pathog. Dis. 2005, 2, 287–303. [Google Scholar] [CrossRef]
- Love, M.J.; Bhandari, D.; Dobson, R.C.J.; Billington, C. Potential for Bacteriophage Endolysins to Supplement or Replace Antibiotics in Food Production and Clinical Care. Antibiotics 2018, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Peng, Q.; Yuan, Y. Characterization of a novel phage infecting the pathogenic multidrug-resistant Bacillus cereus and functional analysis of its endolysin. Appl. Microbiol. Biotechnol. 2018, 102, 7901–7912. [Google Scholar] [CrossRef]
- Schuch, R.; Pelzek, A.J.; Nelson, D.C.; Fischetti, V.A. The PlyB Endolysin of Bacteriophage vB_BanS_Bcp1 Exhibits Broad-Spectrum Bactericidal Activity against Bacillus cereus Sensu Lato Isolates. Appl. Environ. Microbiol. 2019, 85, e00003-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swift, S.M.; Etobayeva, I.V.; Reid, K.P.; Waters, J.J.; Oakley, B.B.; Donovan, D.M.; Nelson, D.C. Characterization of LysBC17, a Lytic Endopeptidase from Bacillus cereus. Antibiotics 2019, 8, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoong, P.; Schuch, R.; Nelson, D.; Fischetti, V.A.; Tremblay, D.M.; Tegoni, M.; Spinelli, S.; Campanacci, V.; Blangy, S.; Huyghe, C.; et al. PlyPH, a Bacteriolytic Enzyme with a Broad pH Range of Activity and Lytic Action against Bacillus anthracis. J. Bacteriol. 2006, 188, 2400–2410. [Google Scholar] [CrossRef] [Green Version]
- Mehta, K.K.; E Paskaleva, E.; Azizi-Ghannad, S.; Ley, D.J.; Page, M.A.; Dordick, J.; Kane, R.S. Characterization of AmiBA2446, a Novel Bacteriolytic Enzyme Active against Bacillus Species. Appl. Environ. Microbiol. 2013, 79, 5899–5906. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kwon, S.-J.; Wu, X.; Sauve, J.; Lee, I.; Nam, J.; Kim, J.; Dordick, J. Selective Killing of Pathogenic Bacteria by Antimicrobial Silver Nanoparticle—Cell Wall Binding Domain Conjugates. ACS Appl. Mater. Interfaces 2018, 10, 13317–13324. [Google Scholar] [CrossRef]
- Lowe, D.E.; Glomski, I.J. Cellular and Physiological Effects of Anthrax Exotoxin and Its Relevance to Disease. Front. Cell. Infect. Microbiol. 2012, 2, 76. [Google Scholar] [CrossRef] [Green Version]
- Abrahams, J.P.; Leslie, A.G.W. Methods used in the structure determination of bovine mitochondrial F1 ATPase. Acta Crystallogr. Sect. D Biol. Crystallogr. 1996, 52, 30–42. [Google Scholar] [CrossRef]
- Drysdale, M.; Heninger, S.; Hutt, J.; Chen, Y.; Lyons, C.R.; Koehler, T.M. Capsule synthesis by Bacillus anthracis is required for dissemination in murine inhalation anthrax. EMBO J. 2004, 24, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Mikesell, P.; E Ivins, B.; Ristroph, J.D.; Dreier, T.M. Evidence for plasmid-mediated toxin production in Bacillus anthracis. Infect. Immun. 1983, 39, 371–376. [Google Scholar] [CrossRef] [Green Version]
- Green, B.D.; Battisti, L.; Koehler, T.M.; Thorne, C.B.; E Ivins, B. Demonstration of a capsule plasmid in Bacillus anthracis. Infect. Immun. 1985, 49, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Makino, S.; Uchida, I.; Terakado, N.; Sasakawa, C.; Yoshikawa, M. Molecular characterization and protein analysis of the cap region, which is essential for encapsulation in Bacillus anthracis. J. Bacteriol. 1989, 171, 722–730. [Google Scholar] [CrossRef] [Green Version]
- Vietri, N.; Marrero, R.; Hoover, T.; Welkos, S.L. Identification and characterization of a trans-activator involved in the regulation of encapsulation by Bacillus anthracis. Gene 1995, 152, 1–9. [Google Scholar] [CrossRef]
- Drysdale, M.; Bourgogne, A.; Koehler, T.M. Transcriptional Analysis of the Bacillus anthracis Capsule Regulators. J. Bacteriol. 2005, 187, 5108–5114. [Google Scholar] [CrossRef] [Green Version]
- Drysdale, M.; Bourgogne, A.; Hilsenbeck, S.G.; Koehler, T.M. atxA Controls Bacillus anthracis Capsule Synthesis via acpA and a Newly Discovered Regulator, acpB. J. Bacteriol. 2004, 186, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Château, A.; Van Schaik, W.; Six, A.; Aucher, W.; Fouet, A. CodY regulation is required for full virulence and heme iron acquisition inBacillus anthracis. FASEB J. 2011, 25, 4445–4456. [Google Scholar] [CrossRef]
- Candela, T.; Fouet, A. Bacillus anthracis CapD, belonging to the γ-glutamyltranspeptidase family, is required for the covalent anchoring of capsule to peptidoglycan. Mol. Microbiol. 2005, 57, 717–726. [Google Scholar] [CrossRef]
- Makino, S.-I.; Watarai, M.; Cheun, H.; Shirahata, T.; Uchida, I. Effect of the Lower Molecular Capsule Released from the Cell Surface ofBacillus anthracison the Pathogenesis of Anthrax. J. Infect. Dis. 2002, 186, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Candela, T.; Balomenou, S.; Aucher, W.; Bouriotis, V.; Simore, J.-P.; Fouet, A.; Boneca, I.G. N-Acetylglucosamine Deacetylases Modulate the Anchoring of the Gamma-Glutamyl Capsule to the Cell Wall ofBacillus anthracis. Microb. Drug Resist. 2014, 20, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Uchida, I.; Makino, S.; Sasakawa, C.; Yoshikawa, M.; Sugimoto, C.; Terakado, N. Identification of a novel gene, dep, associated with depolymerization of the capsular polymer in Bacillus anthracis. Mol. Microbiol. 1993, 9, 487–496. [Google Scholar] [CrossRef]
- Scorpio, A.; Tobery, S.A.; Ribot, W.J.; Friedlander, A.M. Treatment of Experimental Anthrax with Recombinant Capsule Depolymerase. Antimicrob. Agents Chemother. 2007, 52, 1014–1020. [Google Scholar] [CrossRef] [Green Version]
- Negus, D.; Taylor, P.W. A poly-γ-d-glutamic acid depolymerase that degrades the protective capsule ofBacillus anthracis. Mol. Microbiol. 2014, 91, 1136–1147. [Google Scholar] [CrossRef]
- Wu, S.J.; Eiben, C.B.; Carra, J.H.; Huang, I.; Zong, D.; Liu, P.; Wu, C.T.; Nivala, J.; Dunbar, J.; Huber, T.; et al. Improvement of a Potential Anthrax Therapeutic by Computational Protein Design. J. Biol. Chem. 2011, 286, 32586–32592. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Bhatnagar, R.; Gaur, D. Bacillus anthracis Poly-γ-D-Glutamate Capsule Inhibits Opsonic Phagocytosis by Impeding Complement Activation. Front. Immunol. 2020, 11, 11. [Google Scholar] [CrossRef]
- Wang, T.T.; Lucas, A.H. The Capsule of Bacillus anthracis Behaves as a Thymus-Independent Type 2 Antigen. Infect. Immun. 2004, 72, 5460–5463. [Google Scholar] [CrossRef] [Green Version]
- Chabot, D.J.; Ribot, W.J.; Joyce, J.G.; Cook, J.; Hepler, R.; Nahas, D.; Chua, J.; Friedlander, A.M. Protection of rhesus macaques against inhalational anthrax with a Bacillus anthracis capsule conjugate vaccine. Vaccine 2016, 34, 4012–4016. [Google Scholar] [CrossRef]
- Mechaly, A.; Vitner, E.; Levy, H.; Weiss, S.; Bar-David, E.; Gur, D.; Koren, M.; Cohen, H.; Cohen, O.; Mamroud, E.; et al. Simultaneous Immunodetection of Anthrax, Plague, and Tularemia from Blood Cultures by Use of Multiplexed Suspension Arrays. J. Clin. Microbiol. 2018, 56, e01479-17. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chateau, A.; Van der Verren, S.E.; Remaut, H.; Fioravanti, A. The Bacillus anthracis Cell Envelope: Composition, Physiological Role, and Clinical Relevance. Microorganisms 2020, 8, 1864. https://doi.org/10.3390/microorganisms8121864
Chateau A, Van der Verren SE, Remaut H, Fioravanti A. The Bacillus anthracis Cell Envelope: Composition, Physiological Role, and Clinical Relevance. Microorganisms. 2020; 8(12):1864. https://doi.org/10.3390/microorganisms8121864
Chicago/Turabian StyleChateau, Alice, Sander E. Van der Verren, Han Remaut, and Antonella Fioravanti. 2020. "The Bacillus anthracis Cell Envelope: Composition, Physiological Role, and Clinical Relevance" Microorganisms 8, no. 12: 1864. https://doi.org/10.3390/microorganisms8121864
APA StyleChateau, A., Van der Verren, S. E., Remaut, H., & Fioravanti, A. (2020). The Bacillus anthracis Cell Envelope: Composition, Physiological Role, and Clinical Relevance. Microorganisms, 8(12), 1864. https://doi.org/10.3390/microorganisms8121864