Abstract
Salmonella infection can pose serious health issues, especially to children, elders or immunosuppressed humans. Wild populations of reptiles can reach Salmonella prevalence of up to 100% and the direct or indirect transmission from reptiles to humans have been extensively reported. Fernando de Noronha (FN) is an inhabited oceanic archipelago in the northeast coast of Brazil, with an economy based on tourism. The tegu (Salvator merianae) is the largest lizard native to South America and was introduced to the archipelago in the early 20th century. This study determines the prevalence, serotypes, antimicrobial resistance, and molecular epidemiology of Salmonella enterica in the tegu population from FN archipelago. Results show that S. enterica is widely distributed in the FN tegu population, with 43.8% prevalence. The bacteria were isolated from 70.5% of the sampled sites and a total of 15 serotypes were detected in 98 S. enterica isolates. Strains were further classified into 31 genotypes. Recaptured animals presented distinct genotypes in each season, demonstrating a seasonal strain turnover. Most S. enterica isolates from FN tegus presented low antimicrobial resistance. This is possibly due to geographical isolation of the island population, hampering contact with strains from livestock from the continent, where antimicrobial resistance is common.
1. Introduction
Salmonella infection is a major source of gastrointestinal disease in humans, especially children, elders or immunosuppressed individuals [1,2]. Reptiles are common asymptomatic reservoirs of Salmonella, while the bacteria retain the pathogenicity for warm blooded animals [1]. Food of animal origin are considered to be the main sources of Salmonella infection [3], but human infection can also occur when keeping reptiles as pets or through contaminated soil or water [3,4,5]. Wild populations of lizards can reach Salmonella prevalence of up to 100% [6,7], thus bearing a high potential of contamination to other species. Aside from the great importance to public health, there’s also the possibility of impact upon the native fauna such as Salmonella-related mortality in birds [7].
The Brazilian archipelago of Fernando de Noronha is located 340 km offshore from the northeast South American coast and consists of 21 islands and islets. Total land area of the archipelago is 18 km2 where the main island, also named Fernando de Noronha (FN) is about 16.7 km2 wide (Figure 1). The archipelago is a UNESCO world heritage site and has recently been named as a Ramsar site, being visited by over 90 thousand tourists every year [8]. Urbanized areas are restricted to the main island and inside the environmental protected area (APA), but the total number of inhabitants and tourists can reach up to eight thousand people in the peak season [9]. The remainder of the main island, including the other islands and islets from the archipelago, is uninhabited and constitutes the National Park (PARNAMAR), where only indirect use is permitted.
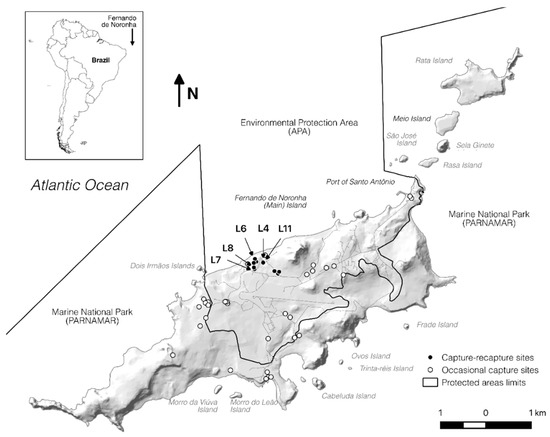
Figure 1.
Fernando de Noronha archipelago. Hollow dots = occasional captures sites, filled dots = capture-recapture sites. L4, L6, L7, L8, L11 correspond to the location sites with higher Salmonella sp. prevalence.
The black and white tegu lizard (Salvator merianae), hereby referred to as tegu, was deliberately introduced to the main island of FN at the beginning of the 20th century [10], where it is currently considered an invasive exotic species. This diurnal and omnivorous species is the largest endemic lizard of the mainland South America where is commonly seen living and feeding close to inhabited areas [11,12,13,14,15]. In most areas where the tegu occurs, they are hunted for their skin and meat [11,16], which has warranted the inclusion of the species on the CITES II appendix [17]. The tegu population in FN was recently estimated as being between seven thousand and twelve thousand individuals [18].
The aims of this study were to determine the prevalence, serotypes, antimicrobial resistance, and molecular epidemiology of Salmonella enterica in the tegu population of Fernando de Noronha archipelago, Brazil.
2. Materials and Methods
2.1. Sampling
The sampling locations were defined to enable the assessment of the tegu population throughout the island, considering low, medium, and high human usage. Sampling periods were determined based on literature data about the mainland tegu, known to hibernate during the autumn–winter seasons [19,20].
Samples were obtained between 2015 and 2016, during five sampling periods of 2–3 weeks of consecutive daily sampling in the main island of the archipelago. Sample periods were at the end (Jan–Feb) and beginning (Oct–Nov) of the dry season. Animals were captured using Tomahawk® and PVC funnel traps, as previously described elsewhere [18], covering areas with low, medium, and high human use. Snout vent length (SVL) was measured to the nearest 0.5 cm, with the use of a tape measure. The weight was taken using a Pesola® scale with 10 g precision. Captured animals were individually marked with a transponder implanted subcutaneously, prior to release. Recaptured individuals in the same sampling season were promptly released and no data or samples were collected. Recapture efforts were done in the same locations every season, so only part of the sampled population was prone to be recaptured.
Captured animals had the cloacal region cleaned with cotton soaked in chlorhexidine alcoholic solution at 0.5% (Riohex®) prior to sample collection. Swabs were introduced in the cloaca avoiding contact with external region. The samples were kept in Stuart’s medium and refrigerated at 4 °C until sent to the laboratory at the end of each campaign (up to three weeks after being obtained).
This study was taken under SISBIO permit no. 41,682 and USP ethics committee no. 2724150515 (approved 17 November 2015).
2.2. Salmonella Isolation
The cloacal swabs were directly plated in Xylose Lysine Tergitol 4 (XLT4) agar (Difco—BBL, Sparks, MD, USA), incubated in aerobiosis at 37 °C for 24–48 h. In parallel, samples were also inoculated in tetrathionate broth and incubated at 37 °C for 24 h, and then plated in XLT4 (Difco) and incubated at 37 °C for 24–48 h. One colony of Salmonella spp. from each sample, previously identified by biochemical tests, was retrieved for further phenotypic and genotypic characterization. The Salmonella isolates were maintained at –80 °C until further serological and molecular typing.
2.3. Serotyping
The antigenic characterization of Salmonella spp. was obtained using the fast agglutination technique based on the antigenic formulas for Salmonella [21] at the Enteric Pathogens Laboratory from Oswaldo Cruz Institute Foundation (FIOCRUZ-RJ).
2.4. Molecular Characterization
Purified DNA was recovered according to Boom et al. [22] protocol and stored at −20 °C. Salmonella suggestive colonies were confirmed through invA gene amplification as previously described [23].
The single-enzyme amplified fragments length polymorphism (SE-AFLP) was performed according to McLauchlin et al. [24] protocol. DNA fragments were detected through electrophoresis at 24 V for 26 h in 2% agarose gel stained with BlueGreen® (LGC Biotecnologia, São Paulo, Brazil) and images were captured under UV illumination by Gel Doc XR System (Bio-Rad Laboratories, Hercules, CA, USA). Molecular weight determinations were done using the 100 bp DNA Ladder (New England BioLabs Inc., Ipswich, MA, USA).
The colistin resistant strains were further assessed for mcr genes (mcr-1 to mcr-5) by PCR using Lescat et al. [25] protocol. The PCR reactions contained 200 μM of each primer, 10× PCR buffer, 1.5 μM MgCl2, 200 μM dNTPs, and 1.25 U of Taq polymerase. Amplified fragments were detected by agarose gel (1.5%) electrophoresis using BlueGreen ™ (LGC Biotecnologia, São Paulo, Brazil) and 100 bp DNA ladder (New England BioLabs Inc., Ipswich, MA, USA).
2.5. Antimicrobial Susceptibility Profiling
The minimal inhibitory concentration (MIC) was determined by broth microdilution technique, as recommended by the Clinical and Laboratory Standards Institute supplement VET08 [26], using a panel of 18 selected antimicrobials: Ceftiofur, Amoxicillin/Clavulanate, Ampicillin, Meropenem, Fosfomycin, Oxytetracycline, Chloramphenicol, Florfenicol, Nalidixic Acid, Ciprofloxacin, Marbofloxacin, Gentamicin, Neomycin, Azithromycin, Colistin, Sulfamethoxazole, and Trimethoprim/Sulfamethoxazole (MilliporeSigma, St. Louis, MO, USA). Staphylococcus aureus ATCC 29,213 was used as internal quality control.
2.6. Statistical Analysis
The distribution of strains frequencies according to origin, serotype, resistance, and SE-AFLP profile was performed with SPSS 16.0 (SPSS Inc., Chicago, IL, USA). The multidrug resistance, MIC50 and MIC90 values for the respective antimicrobials were determined according to Schwarz et al. [27].
SE-AFLP results were analyzed with Bionumerics 7.6 software (Applied Maths NV, Saint-Martens-Latem, Belgium). Fingerprint patterns were analyzed by a comprehensive pairwise comparison of restriction fragment sizes, using the Dice coefficient. The mean values obtained from Dice coefficients were employed in UPGMA (unweighted pair group method with arithmetic mean) to generate a dendrogram. A cut-off value of 90% of genetic similarity was applied to analyze the resulting clusters [28], and the discriminatory index was calculated as previously described by Hunter and Gaston [29].
3. Results
A total of 153 animals were captured, between 2015 and 2016, on 44 different sites in the main island of FN (Figure 1). Although tegu is present in the Rata Island—as verified through indirect signs—our efforts returned no captures there. The capture locations were classified according to the human use to facilitate understanding the exposure risk.
From the 153 captured animals, 62 were female, 88 were male and in three individuals the gender could not be determined. One hundred and fifteen individuals from the 14 fix sites (filled dots—Figure 1) were captured, marked, and released. Thirty-eight animals from 30 occasional sites (hollow dots—Figure 1) were collected. From the released animals, 26 were recaptured in two different periods, seven were recaptured in three different periods, and only one was recaptured in four out of five collects; a total of 196 cloacal samples taken. From 153 studied animals, 67 (43.8%) were positive for Salmonella isolation in at least one sample.
From 196 samples taken, 98 (50.0%) were positive to Salmonella sp. Although 70.5% (31/44) of sampled locations were positive for Salmonella isolation, half of the isolates originated from only four sites (L6, L4, L11, L8). Most isolates were obtained from collects 1 and 4 (C1 and C4) (30.6% and 23.5%, respectively), followed by C5 with 20.4%, C3 with 17.3%, and the remaining 8.2% originated from C2 collect.
All isolates were identified as Salmonella enterica subsp. enterica by PCR. Only 12.2% of isolates were not able to be classified by serotyping (Table 1). Rubislaw and Javiana were the most frequent serotypes (13.3% and 12.2%, respectively), followed by Mbandaka, Panama, and Muenchen with 9.2% each, and Minnesota with 7.1%, comprising 58.2% of studied isolates. It is highlighted that only serotype Rubislaw was detected in all five collects (Table 1).

Table 1.
Distribution of detected serotypes among sampling periods—N (%).
Genotyping by SE-AFLP resulted in 31 profiles (A1–A31) (Figure 2). Cluster analysis of fingerprint patterns enabled differentiation of one main group composed by 80 isolates, from 18 genotypes (A1–A18), with over 75% genetic similarity. The remaining 18 isolates presented higher genetic heterogeneity and were distributed among 13 SE-AFLP profiles, and most of them were isolated in the first collect (C1). Three genotypes (A1, A2, and A16) comprised 42.9% of the 98 studied isolates.

Figure 2.
Dendrogram showing the relationship among single-enzyme amplified fragments length polymorphism (SE-AFLP) genotypes and antimicrobial resistance profiles of S. enterica strains from tegus of Fernando de Noronha.
There was no clear tendency of clustering according to isolates origin or serotype. Nevertheless, there is a slight tendency to cluster according to the collects, a portion from A1 and A2 genotypes that present higher variety of origin. The discriminatory index obtained for SE-AFLP technique was 0.92. Only two sets of isolates from recaptured animals presented persistence of SE-AFLP profiles (T67 and T68), while most isolates from recaptured animals presented distinct genotypes at each collect (Table 2).
Table 2.
Assessment of recaptured animals among sampling periods according to isolation sites and AFLP genotypes (cells are colored according to isolation sites).
All isolates were susceptible to ceftiofur, meropenem, fosfomycin, oxytetracycline, chloramphenicol, marbofloxacin, gentamycin, neomycin, azithromycin, and trimethoprim/sulfamethoxazole (Table 3).
Table 3.
Minimal inhibitory concentration (MIC) range, MIC50, MIC90, and resistance rates of Salmonella isolates against tested antimicrobials. S = susceptible; I = intermediate; R = resistant.
Interestingly, 13.3% of isolates were resistant to colistin, 10.2% to sulfamethoxazole, and 21.4% presented intermediate susceptibility to florfenicol. Only one isolate also presented resistance to ampicillin and amoxicillin/clavulanate, while other was resistant solely to nalidixic acid and ciprofloxacin; a total of 55 isolates (56.1%) were susceptible to the 18 tested antimicrobials. The observed MIC values for colistin, sulfamethoxazole, and florfenicol were further compared to the EUCAST (European Committee on Antimicrobial Susceptibility Testing) MIC distributions and epidemiological cut-off values (ECOFFs) (https://mic.eucast.org/Eucast2/) for Salmonella enterica (Figure 3).
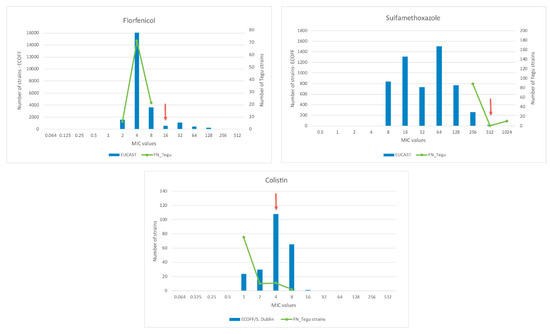
Figure 3.
Distribution of S. enterica MICs for florfenicol, sulfamethoxazole, and colistin in Fernando de Noronha. Bars correspond to European Committee on Antimicrobial Susceptibility Testing (EUCAST) data of the respective antimicrobials for Salmonella enterica; lines correspond to Fernando de Noronha tegu’s S. enterica MICs; arrows correspond to antimicrobials resistance breakpoints.
4. Discussion
The high Salmonella prevalence (43.8%) observed among wild tegu corroborates the literature for Salmonella occurrence in captive reptiles worldwide [30,31,32,33,34] and the few Brazilian studies that report up to 100% positivity for captive tegus in two Brazilian States [6,35].
At the time of this study, only one isolate of each sample was selected for further analyses; this could represent a drawback considering the possibility of Salmonella heterogeneity with the studied animals. Nevertheless, it is interesting that all studied isolates were identified as Salmonella enterica subsp. enterica. Although reptiles have been described as hosts of a wide variety of Salmonella enterica subspecies and exotic serotypes [5,33,36], the predominance of subspecies enterica demands attention for the zoonotic potential as its related serotypes have been commonly isolated from human, alimentary, and environmental sources.
SE-AFLP analysis resulted in high genetic variability among S. enterica from wild tegu, with genetic profiles comprising isolates from different serotypes. Even though genetic diversity of reptile Salmonella has been poorly assessed, pulsed field gel electrophoresis (PFGE) typing has already demonstrated inter- and intra-serotype heterogeneity. Franco et al. [37] reported genetic similarity between serotypes (Newport and Bardo) and variation of S. Pomona within nine pulsotypes among Salmonella isolated from land iguanas in an Ecuador island. Similarly, Bertelloni et al. [38] also reported different pulsotypes among Salmonella strains from the same serotype isolated from healthy pet reptiles in pet shops in Italy.
In this study, genetic heterogeneity was observed not only among Salmonella serotypes but also between the studied areas of FN island. Considering the landscape and that tegus in FN have a home range of up to 15 hectares [18], the detection of the same genotype in distant capture areas at the same sampling season suggests other species could be involved in Salmonella dissemination throughout the island. Humans, birds [39,40] or domestic animals could be involved on this transmission chain, but further studies are required to stablish the complete Salmonella dynamics in FN.
The large number of different serotypes detected with only a few representatives of each serotype corroborates previous reports of high diversity of Salmonella serotypes among reptiles [5,32,33,41]. Most of the serotypes detected in wild tegus had already been reported in reptiles worldwide, of which Agona, Braenderup, Panama, Rubislaw, Saintpaul, and Worthington serotypes had been previously detected in Brazilian captive tegus [6,35]. As previously stated, most of these serotypes have also been identified in humans, birds (including poultry), pigs, and cattle [40].
In contrast to the literature, the S. enterica isolates from FN wild tegus presented low prevalence of antimicrobial resistance. Chen et al. [42] and Hossain et al. [43] have reported high drug resistance rates in S. enterica isolated from reptiles and eggs in Asia, including over 30% of cephalosporin resistance. In opposition, our study finds 56.1% of the isolates were susceptible to all tested antimicrobials. This found suggests a geographically isolated population that is not in contact with strains common to domestic animals in the continent, where antimicrobial resistance is common in S. enterica. Isolation has also been genetically demonstrated for rat populations in Fernando de Noronha and Rata islands in FN archipelago [44].
Interestingly, resistance to colistin and sulfamethoxazole, and intermediate susceptibility to florfenicol were detected among studied isolates. However, Figure 3 demonstrates that when our MIC values are compared to the EUCAST data the MICs distributions are very similar to the values described in the wild type Salmonella strains (not exposed to antimicrobials). In addition, the identified colistin resistant strains were further assessed for mcr genes by PCR, and all were negative for mcr-1 to mcr-5 genes presence.
Considering that the archipelago receives tourists of all age groups year-round, the identification of wild tegus as spreaders of Salmonella enterica subsp. enterica serotypes is of great concern to public health. Moreover, it should be noted that tegu lizards are not used as a food source in FN. From a conservation perspective, adding commercial value to tegu in FN (e.g., using it as a protein source) will likely create dependence on this resource by the local community, creating further obstacles to any control or eradication program in the future and perpetuating the establishment of tegu in FN archipelago.
Author Contributions
Conceptualization, C.R.A., L.Z.M., A.M.M., and R.A.D.; methodology, C.R.A., L.Z.M., C.E.C.M., N.R.B., and R.A.D.; formal analysis, C.R.A., L.Z.M., and R.A.D.; investigation, C.R.A. and L.Z.M.; resources, F.F., J.C.R.S., and R.A.D.; writing—original draft preparation, C.R.A., L.Z.M., and R.A.D.; writing—review and editing, C.R.A., L.Z.M., J.C.R.S., F.F., A.M.M., and R.A.D.; supervision, A.M.M. and R.A.D.; project administration, C.R.A. and R.A.D.; funding acquisition, C.R.A., F.F., A.M.M., and R.A.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by ICMBio (PVE Project Number 88881.065000/2014-1) and CAPES (Finance code 001). LZM and CECM are recipients of São Paulo Research Foundation (FAPESP) fellowships (grants 2016/25745-7 and 2015/26159-1).
Acknowledgments
The authors would like to thank the NGI of Fernando de Noronha protected areas and its volunteers; the fire department of Fernando de Noronha for logistical support, Fernando de Noronha Administration; Priscilla Anne Melville for technical laboratory support; Brazilian Institute for Conservation Medicine (Tríade) for technical and logistical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Corrente, M.; Totaro, M.; Martella, V.; Campolo, M.; Lorusso, A.; Ricci, M.; Buonavoglia, C. Reptile-associated Salmonellosis in Man, Italy. Emerg. Infect. Dis. 2006, 12, 358–359. [Google Scholar] [CrossRef] [PubMed]
- Fornazari, F.; Teixeira, C.R. Salmonelose em répteis: Aspectos epidemiológicos, clínicos e zoonóticos. Veterinária Zootec. 2009, 16, 19–25. [Google Scholar]
- Tomastikova, Z.; Romero, S.B.; Knotek, Z.; Karpiskova, R. Prevalence and characteristics of Salmonella species isolated from captive reptiles in the Czech Republic. Veterinární Med. 2017, 62, 456–469. [Google Scholar] [CrossRef]
- Warwick, C.; Lambiris, A.J.L.; Westwood, D.; Steedman, C. Reptile-Related Salmonellosis in Europe; SAGE Publications Sage UK: London, UK, 2001; Volume 94, pp. 124–126. [Google Scholar]
- Whiley, H.; Gardner, M.G.; Ross, K. A Review of Salmonella and Squamates (Lizards, Snakes and Amphisbians): Implications for Public Health. Pathogens 2017, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Maciel, B.M.; Filho, R.C.A.; Nogueira, S.S.D.C.; Dias, J.C.T.; Rezende, R.P. High Prevalence of Salmonella in Tegu Lizards (Tupinambis merianae), and Susceptibility of the Serotypes to Antibiotics. Zoonoses Public Health 2009, 57, e26–e32. [Google Scholar] [CrossRef] [PubMed]
- Kikillus, K.H.; Gartrell, B.D.; Motion, E. Prevalence of Salmonella spp., and serovars isolated from captive exotic reptiles in New Zealand. N. Z. Veter. J. 2011, 59, 174–178. [Google Scholar] [CrossRef]
- Ribeiro, C. Fernando de Noronha Registra alta no Fluxo Turístico em 2016. Available online: http://www.turismo.gov.br/ultimas-noticias/7397-fernando-de-noronha-registra-alta-no-fluxo-turístico-em-2016.html (accessed on 15 April 2020).
- Marinho, A.C. Qual a População de Fernando de Noronha? 2016. Available online: http://g1.globo.com/pernambuco/blog/viver-noronha/post/qual-populacao-de-fernando-de-noronha.html (accessed on 15 April 2020).
- Santos, A.T. Fernando de Noronha, Ilha de Cenários Múltiplos, 1st ed.; RIO ed: Rio de Janeiro, Brazil, 1950; 143p. [Google Scholar]
- Oren, D.C. Resultados de uma nova expedição zoológica a Fernando de Noronha. Bol. Mus. Para. Emílio. Goeldi. Zool. 1984, 1, 19–44. [Google Scholar]
- Sazima, I.; Haddad, C.F.B. Repteis da Serra do Japi: Notas Sobre Historia Natural. In História Nat da Serra do Japi Ecol e Preserv uma área Florest no Sudeste do Bras; Morellato, L.P.C., Ed.; Editora da Unicamp/FAPESP: Campinas, Brazil, 1992; p. 321. [Google Scholar]
- Bovendorp, R.S.; Alvarez, A.D.; Galetti, M. Density of the tegu lizard (Tupinambis merianae) and its role as nest predator at Anchieta island, Brazil. Neotrop. Biol. Conserv. 2008, 3, 9–12. [Google Scholar]
- Winck, G.R.; Blanco, C.C.; Cechin, S.Z. Population Ecology of Tupinambis merianae (Squamata, Teiidae): Home-Range, Activity and Space Use. Animal Biology. 2011, 61, 493–510. [Google Scholar] [CrossRef]
- Klug, P.E.; Reed, R.N.; Mazzotti, F.J.; McEachern, M.A.; Vinci, J.J.; Craven, K.K.; Adams, A.A.Y. The influence of disturbed habitat on the spatial ecology of Argentine black and white tegu (Tupinambis merianae), a recent invader in the Everglades ecosystem (Florida, USA). Biol. Invasions. 2015, 17, 1785–1797. [Google Scholar] [CrossRef]
- Nóbrega Alves, R.R.; Filho, G.A.P.; Vieira, K.S.; Souto, W.M.S.; Mendonça, L.E.T.; Montenegro, P.; Almeida, W.D.O.; Vieira, W.L.S. A zoological catalogue of hunted reptiles in the semiarid region of Brazil. J. Ethnobiol. Ethnomedicine 2012, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- CITES. Appendices I, II and III of Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES). 2017. Available online: https://www.cites.org/eng/app/appendices.php (accessed on 20 April 2020).
- Abrahão, C.R.; Russell, J.C.; Silva, J.C.R.; Ferreira, F.; Dias, R.A. Population Assessment of A Novel Island Invasive: Tegu (Salvator merianae) of Fernando de Noronha. In Proceedings of the International Conference on Island Invasives 2017, Dundee, UK, 14 July 2017; pp. 313–321. [Google Scholar]
- Andrade, D.V.; Sanders, C.; Milsom, W.K.; Abe, A.S. Overwintering in Tegu Lizards. In Life in the Cold: Evolution, Mechanisms, Adaptation, and Application, Twelfth International Hibernation Symposium; Barnes, B.M., Carey, H.V., Eds.; Biological Papers of the University of Alaska, number 27; University of Alaska: Fairbanks, AK, USA, 2004; pp. 339–348. [Google Scholar]
- De Souza, S.C.R.; De Carvalho, J.E.; Abe, A.S.; Bicudo, J.E.P.W.; Bianconcini, M.S.C. Seasonal metabolic depression, substrate utilization and changes in scaling patterns during the first year cycle of tegu lizards (Tupinambis merianae). J. Exp. Biol. 2004, 207, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Popoff, M.Y.; Le Minor, L. Antigenic Formulas of the Salmonella Serovars, WHO Collaborating Centre for Reference and Research on Salmonella; World Heal Organ: Geneva, Switzerland, 2001. [Google Scholar]
- Boom, R.; Sol, C.J.; Salimans, M.M.; Jansen, C.L.; Dillen, P.M.W.-V.; Van Der Noordaa, J. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Stone, G.G.; Oberst, R.D.; Hays, M.P.; McVey, S.; Chengappa, M.M. Detection of Salmonella serovars from clinical samples by enrichment broth cultivation-PCR procedure. J. Clin. Microbiol. 1994, 32, 1742–1749. [Google Scholar] [CrossRef] [PubMed]
- McLauchlin, J.; Ripabelli, G.; Brett, M.; Threlfall, E. Amplified fragment length polymorphism (AFLP) analysis of Clostridium perfringens for epidemiological typing. Int. J. Food Microbiol. 2000, 56, 21–28. [Google Scholar] [CrossRef]
- Lescat, M.; Poirel, L.; Nordmann, P. Rapid multiplex polymerase chain reaction for detection of mcr-1 to mcr-5 genes. Diagn. Microbiol. Infect. Dis. 2018, 92, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.L. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; CLSI Suppl. VET08; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; Van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Editorial: Assessing the antimicrobial susceptibility of bacteria obtained from animals. J. Antimicrob. Chemother. 2010, 65, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Van Belkum, A.; Tassios, P.; Dijkshoorn, L.; Haeggman, S.; Cookson, B.; Fry, N.; Fussing, V.; Green, J.; Feil, E.; Gerner-Smidt, P.; et al. Guidelines for the validation and application of typing methods for use in bacterial epidemiology. Clin. Microbiol. Infect. 2007, 13, 1–46. [Google Scholar] [CrossRef]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef]
- Geue, L.; Löschner, U. Salmonella enterica in reptiles of German and Austrian origin. Veter Microbiol. 2002, 84, 79–91. [Google Scholar] [CrossRef]
- Corrente, M.; Madio, A.; Friedrich, K.; Greco, G.; Desario, C.; Tagliabue, S.; D’Incau, M.; Campolo, M.; Buonavoglia, C. Isolation of Salmonella strains from reptile faeces and comparison of different culture media. J. Appl. Microbiol. 2004, 96, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Bauwens, L.; Vercammen, F.; Bertrand, S.; Collard, J.-M.; De Ceuster, S. Isolation of Salmonella from environmental samples collected in the reptile department of Antwerp Zoo using different selective methods. J. Appl. Microbiol. 2006, 101, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.; Nordentoft, S.; Hammer, A.S.; Lassen-Nielsen, A.-M. Serovars of Salmonella from captive reptiles. Zoonoses Public Health 2009, 56, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.Y.; Wong, S.P.; Dykes, G.A. Salmonella associated with captive and wild lizards in Malaysia. Herpetol. Notes 2014, 7, 145–147. [Google Scholar]
- Carvalho, A.D.M.; Júnior, A.K.P.; Andrade, M.A.; Jayme, V.D.S. Prevalência de Enterobacteriaceae em Tupinambis merianae (Squamata: Teiidae) de uma instalação em cativeiro no Brasil Central, com um perfil de resistência aos medicamentos antimicrobianos em Salmonella enterica. Phyllomedusa: J. Herpetol. 2012, 12, 57. [Google Scholar] [CrossRef][Green Version]
- Mitchell, M.A.; Shane, S.M. Salmonella in reptiles. Semin. Avian Exot. Pet Med. 2001, 10, 25–35. [Google Scholar] [CrossRef]
- Franco, A.; Hendriksen, R.; Lorenzetti, S.; Onorati, R.; Gentile, G.; Dell’Omo, G.; Aarestrup, F.M.; Battisti, A. Characterization of Salmonella Occurring at high prevalence in a population of the land iguana Conolophus subcristatus in Galápagos Islands, Ecuador. PLoS ONE 2011, 6, e23147. [Google Scholar] [CrossRef]
- Bertelloni, F.; Chemaly, M.; Cerri, D.; Le Gall, F.; Ebani, V.V. Salmonella infection in healthy pet reptiles: Bacteriological isolation and study of some pathogenic characters. Acta Microbiol. Immunol. Hung. 2016, 63, 203–216. [Google Scholar] [CrossRef]
- Silva, M.A.; Marvulo, M.F.; Mota, R.A.; Silva, J.C. A importância da ordem Ciconiiformes na cadeia epidemiológica de Salmonella spp. para a saúde pública e a conservação da diversidade biológica. Pesqui. Veterinária Bras. 2010, 30, 573–580. [Google Scholar] [CrossRef]
- Silva, M.A.; Érika, F.; Santana, S.C.; Marvulo, M.F.V.; Barros, M.R.; Vilela, S.M.; Reis, E.M.; Mota, R.A.; Silva, J.C. Isolation of Salmonella spp. in cattle egrets (Bubulcus ibis) from Fernando de Noronha Archipelago, Brazil. Braz. J. Microbiol. 2018, 49, 559–563. [Google Scholar] [CrossRef]
- Pasmans, F.; Martel, A.; Boyen, F.; Vandekerchove, D.; Wybo, I.; Van Immerseel, F.; Heyndrickx, M.; Collard, J.M.; Ducatelle, R.; Haesebrouck, F. Characterization of Salmonella isolates from captive lizards. Vet Microbiol. 2005, 110, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Chen, W.-C.; Chin, S.-C.; Lai, Y.-H.; Tung, K.-C.; Chiou, C.-S.; Hsu, Y.-M.; Chang, C.-C. Prevalence and antimicrobial susceptibility of Salmonellae isolates from reptiles in Taiwan. J. Veter Diagn. Investig. 2010, 22, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S.; Hossain, K.M.M.; Alam Sarker, M.; Hamid, S.A. Prevalence and antibiotic susceptibility of Salmonella from chicken eggs in Naogaon district of Bangladesh. J. Adv. Microbiol. 2019, 19, 1–6. [Google Scholar] [CrossRef]
- Gatto-Almeida, F.; Pichlmueller, F.; Micheletti, T.; Abrahão, C.R.; Mangini, P.R.; Russell, J.C. Using genetics to plan black rat (Rattus rattus) management in Fernando de Noronha archipelago, Brazil. Perspect. Ecol. Conserv. 2020, 18, 44–50. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).