Benzalkonium Chloride Induces a VBNC State in Listeria monocytogenes

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Listeria monocytogenes Isolate under Study and Biocide Susceptibility Testing
2.2. Adaptation to Benzalkonium Chloride
2.3. Determination of Culturable Cells after BC Exposure
2.4. Antibiotic Susceptibility Testing and Flow Cytometric Analysis
2.5. Statistics
3. Results
3.1. Adapted L. monocytogenes Cells Show Shifts in MICs of BC and Some Antibiotics
3.2. Adaptation Increased the Proportion of Cells with Intact Membranes at high BC Concentrations
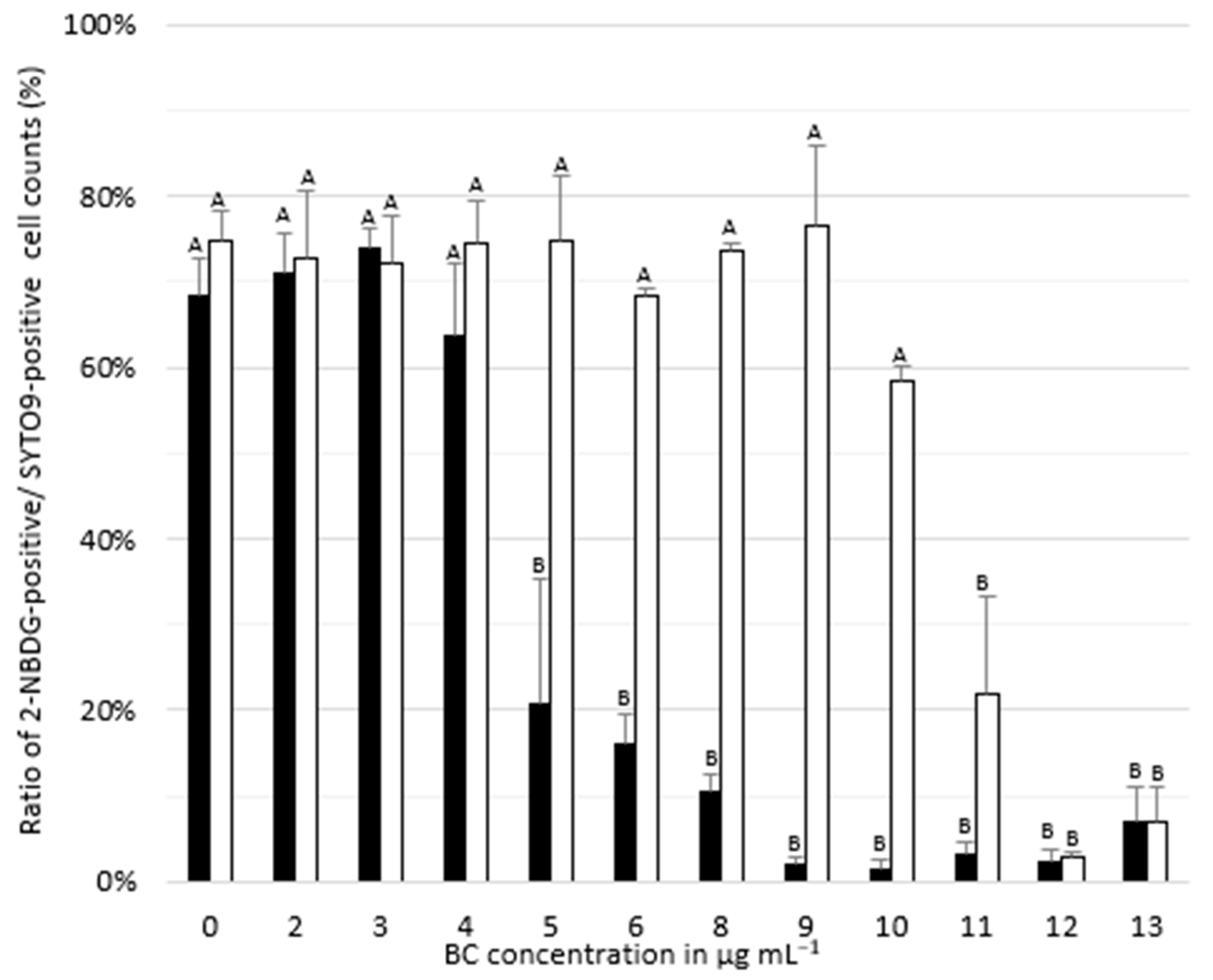
3.3. Adapted Cells Maintain Metabolic Activity at High BC Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swaminathan, B.; Gerner-Smidt, P. The epidemiology of human listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allerberger, F. Listeria: Growth, phenotypic differentiation and molecular microbiology. FEMS Immunol. Med. Microbiol. 2003, 3, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Weiler, C.; Ifland, A.; Naumann, A.; Kleta, S.; Noll, M. Incorporation of Listeria monocytogenes strains in raw milk biofilms. Int. J. Food Microbiol. 2013, 161, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- McLauchlin, J.; Mitchell, R.T.; Smerdon, W.J.; Jewell, K. Listeria monocytogenes and listeriosis: A review of hazard characterisation for use in microbiological risk assessment of foods. Int. J. Food Microbiol. 2004, 92, 15–33. [Google Scholar] [CrossRef]
- Szendy, M.; Westhaeuser, F.; Baude, B.; Reim, J.; Dähne, L.; Noll, M. Controlled release of nisin from Neusilin particles to enhance food safety of sour curd cheese. J. Food Sci. Technol. 2019, 56, 1613–1621. [Google Scholar] [CrossRef]
- Besnard, V.; Federighi, M.; Declerq, E.; Jugiau, F.; Cappelier, J.-M. Environmental and physico-chemical factors induce VBNC state in Listeria monocytogenes. Vet. Res. 2002, 33, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.D. Formation of viable but nonculturable cells. In Starvation in Bacteria; Kjelleberg, S., Ed.; Springer: Boston, MA, USA, 1993; pp. 239–272. ISBN 978-0-306-44430-2. [Google Scholar]
- Lin, H.; Ye, C.; Chen, S.; Zhang, S.; Yu, X. Viable but non-culturable E. coli induced by low level chlorination have higher persistence to antibiotics than their culturable counterparts. Environ. Pollut. 2017, 230, 242–249. [Google Scholar] [CrossRef]
- Lindbäck, T.; Rottenberg, M.E.; Roche, S.M.; Rørvik, L.M. The ability to enter into an avirulent viable but non-culturable (VBNC) form is widespread among Listeria monocytogenes isolates from salmon, patients and environment. Vet. Res. 2010, 41, 8. [Google Scholar] [CrossRef] [Green Version]
- Dreux, N.; Albagnac, C.; Federighi, M.; Carlin, F.; Morris, C.E.; Nguyen-the, C. Viable but non-culturable Listeria monocytogenes on parsley leaves and absence of recovery to a culturable state. J. Appl. Microbiol. 2007, 103, 1272–1281. [Google Scholar] [CrossRef]
- Xu, H.S.; Roberts, N.; Singleton, F.L.; Attwell, R.W.; Grimes, D.J.; Colwell, R.R. Survival and viability of nonculturable Escherichia coli and Vibrio cholerae in the estuarine and marine environment. Microb. Ecol. 1982, 8, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Robben, C.; Witte, A.K.; Schoder, D.; Stessl, B.; Rossmanith, P.; Mester, P. A fast and easy ATP-based approach enables MIC testing for non-resuscitating VBNC pathogens. Front. Microbiol. 2019, 10, 1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desneux, J.; Biscuit, A.; Picard, S.; Pourcher, A.-M. Fate of viable but non-culturable Listeria monocytogenes in pig manure microcosms. Front. Microbiol. 2016, 7, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afari, G.K.; Hung, Y.-C. Detection and verification of the viable but nonculturable (VBNC) state of Escherichia coli O157:H7 and Listeria monocytogenes using flow cytometry and standard plating. J. Food Sci. 2018, 83, 1913–1920. [Google Scholar] [CrossRef] [PubMed]
- Berney, M.; Weilenmann, H.-U.; Egli, T. Flow-cytometric study of vital cellular functions in Escherichia coli during solar disinfection (SODIS). Microbiology 2006, 152, 1719–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morishige, Y.; Fujimori, K.; Amano, F. Use of flow cytometry for quantitative analysis of metabolism of viable but non-culturable (VBNC) Salmonella. Biol. Pharm. Bull. 2015, 38, 1255–1264. [Google Scholar] [CrossRef]
- Merchel Piovesan Pereira, B.; Tagkopoulos, I. Benzalkonium chlorides: Uses, regulatory status, and microbial resistance. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [Green Version]
- To, M.S.; Favrin, S.; Romanova, N.; Griffiths, M.W. Postadaptational resistance to benzalkonium chloride and subsequent physicochemical modifications of Listeria monocytogenes. Appl. Env. Microbiol. 2002, 68, 5258–5264. [Google Scholar] [CrossRef] [Green Version]
- Wessels, S.; Ingmer, H. Modes of action of three disinfectant active substances: A review. Regul. Toxicol. Pharmacol. 2013, 67, 456–467. [Google Scholar] [CrossRef]
- Romanova, N.A.; Wolffs, P.F.G.; Brovko, L.Y.; Griffiths, M.W. Role of efflux pumps in adaptation and resistance of Listeria monocytogenes to benzalkonium chloride. Appl. Env. Microbiol. 2006, 72, 3498–3503. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Jiang, X.; Zhang, Y.; Ji, S.; Gao, W.; Shi, L. Effect of benzalkonium chloride adaptation on sensitivity to antimicrobial agents and tolerance to environmental stresses in Listeria monocytogenes. Front. Microbiol. 2018, 9, 2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roedel, A.; Dieckmann, R.; Brendebach, H.; Hammerl, J.A.; Kleta, S.; Noll, M.; Al Dahouk, S.; Vincze, S. Biocide tolerant Listeria monocytogenes isolates from German food production plants do not show cross-resistance to clinically relevant antibiotics. Appl. Environ. Microbiol. 2019. [CrossRef] [Green Version]
- DIN EN ISO 11290. Mikrobiologie der Lebensmittelkette- Horizontales Verfahren für den Nachweis und die Zählung von Listeria monocytogenes und von Listeria spp.- Teil 1: Nachweisverfahren (ISO 11290-1:2017); Deutsche Fassung EN ISO 11290-1:2017; Beuth Verlag GmbH: Berlin, Germany.
- Noll, M.; Kleta, S.; Al Dahouk, S. Antibiotic susceptibility of 259 Listeria monocytogenes strains isolated from food, food-processing plants and human samples in Germany. J. Infect. Public Health 2018, 11, 572–577. [Google Scholar] [CrossRef] [PubMed]
- R Core Team v. 3.3.3. R: A Language and Environment for Statistical Computing; R Core Team v. 3.3.3: Vienna, Austria, 2017. [Google Scholar]
- Lundén, J.; Autio, T.; Markkula, A.; Hellström, S.; Korkeala, H. Adaptive and cross-adaptive responses of persistent and non-persistent Listeria monocytogenes strains to disinfectants. Int. J. Food Microbiol. 2003, 82, 265–272. [Google Scholar] [CrossRef]
- Aase, B.; Sundheim, G.; Langsrud, S.; Rørvik, L.M. Occurrence of and a possible mechanism for resistance to a quaternary ammonium compound in Listeria monocytogenes. Int. J. Food Microbiol. 2000, 62, 57–63. [Google Scholar] [CrossRef]
- Rakic-Martinez, M.; Drevets, D.A.; Dutta, V.; Katic, V.; Kathariou, S. Listeria monocytogenes strains selected on ciprofloxacin or the disinfectant benzalkonium chloride exhibit reduced susceptibility to ciprofloxacin, gentamicin, benzalkonium chloride, and other toxic compounds. Appl. Env. Microbiol. 2011, 77, 8714–8721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Léonard, L.; Bouarab Chibane, L.; Ouled Bouhedda, B.; Degraeve, P.; Oulahal, N. Recent advances on multi-parameter flow cytometry to characterize antimicrobial treatments. Front. Microbiol. 2016, 7, 1225. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Faucher, S.P. The importance of the viable but non-culturable state in human bacterial pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [CrossRef] [Green Version]
- Highmore, C.J.; Warner, J.C.; Rothwell, S.D.; Wilks, S.A.; Keevil, C.W. Viable-but-nonculturable Listeria monocytogenes and Salmonella enterica serovar Thompson induced by chlorine stress remain infectious. MBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Minimum Inhibitory Concentrations in µg mL−1 | A | |||||||||||||
| VAN | T/S | TGC | TET | RIF | PEN | MER | LIZ | GEN | ERY | DPT | CRO | CIP | AMP | BC [µg mL−1]1 |
| 1 | 0.0625/1.1875 | 0.0625 | 1 | 0.125 | 0.25 | 0.5 | 2 | 2 | <0.25 | 16 | 128 | 4 | 0.25 | 0 |
| <1 | 0.0625/1.1875 | 0.0625 | 0.5 | 0.125 | 0.125 | 0.125 | 1 | 2 | <0.25 | 16 | 128 | 4 | 0.25 | 1 |
| <1 | 0.0625/1.1875 | 0.0625 | 0.25 | 0.0625 | 0.0625 | 0.0625 | 0.5 | 0.5 | <0.25 | 16 | 64 | 1 | 0.125 | 2 |
| <1 | 0.03125/0.59375 | <0.03125 | <0.25 | <0.0625 | <0.0625 | <0.0625 | <0.5 | <0.5 | <0.25 | <0.5 | <1 | <0.25 | <0.0625 | 3 |
| Minimum Inhibitory Concentrations in µg mL−1 | B | |||||||||||||
| VAN | T/S | TGC | TET | RIF | PEN | MER | LIZ | GEN | ERY | DPT | CRO | CIP | AMP | BC [µg mL−1] 1 |
| 1 | <0.03125/0.59375 | 0.0625 | 0.5 | 0.25 | 0.25 | 0.5 | 1 | 1 | <0.25 | 16 | 64 | 4 | 0.25 | 0 |
| <1 | <0.03125/0.59375 | <0.03125 | <0.25 | 0.0625 | 0.0625 | 0.125 | 0.5 | <0.5 | <0.25 | 4 | 16 | 0.5 | 0.25 | 1 |
| <1 | <0.03125/0.59375 | <0.03125 | <0.25 | <0.0625 | 0.0625 | <0.0625 | <0.5 | <0.5 | <0.25 | <0.5 | 1 | <0.25 | <0.0625 | 2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noll, M.; Trunzer, K.; Vondran, A.; Vincze, S.; Dieckmann, R.; Al Dahouk, S.; Gold, C. Benzalkonium Chloride Induces a VBNC State in Listeria monocytogenes. Microorganisms 2020, 8, 184. https://doi.org/10.3390/microorganisms8020184
Noll M, Trunzer K, Vondran A, Vincze S, Dieckmann R, Al Dahouk S, Gold C. Benzalkonium Chloride Induces a VBNC State in Listeria monocytogenes. Microorganisms. 2020; 8(2):184. https://doi.org/10.3390/microorganisms8020184
Chicago/Turabian StyleNoll, Matthias, Katharina Trunzer, Antje Vondran, Szilvia Vincze, Ralf Dieckmann, Sascha Al Dahouk, and Carolin Gold. 2020. "Benzalkonium Chloride Induces a VBNC State in Listeria monocytogenes" Microorganisms 8, no. 2: 184. https://doi.org/10.3390/microorganisms8020184
APA StyleNoll, M., Trunzer, K., Vondran, A., Vincze, S., Dieckmann, R., Al Dahouk, S., & Gold, C. (2020). Benzalkonium Chloride Induces a VBNC State in Listeria monocytogenes. Microorganisms, 8(2), 184. https://doi.org/10.3390/microorganisms8020184