Cording Mycobacterium tuberculosis Bacilli Have a Key Role in the Progression towards Active Tuberculosis, Which is Stopped by Previous Immune Response

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Batch Production and Experimental Design
2.2. Image Analysis of Bacillary Aggregates
2.3. Animals and Ethics
2.4. BL
2.5. Pathology
2.6. Ex Vivo SEM
2.7. Immune Response
2.8. Statistics
3. Results
3.1. Cording Mtb Induces a Significant Change in the Progression of the Infection
3.2. Cording Bacilli Can Be Detected in Exudative Lesions
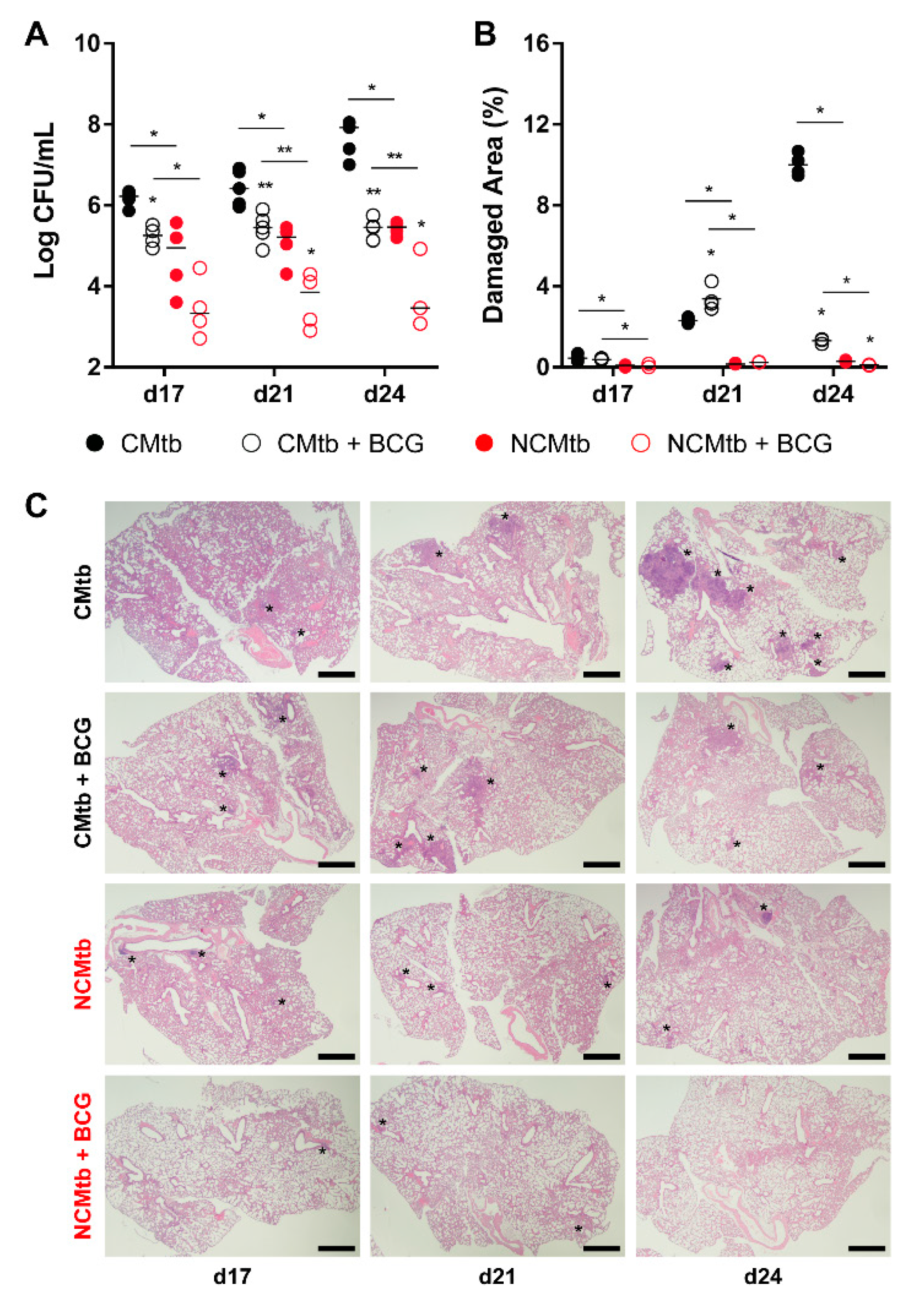
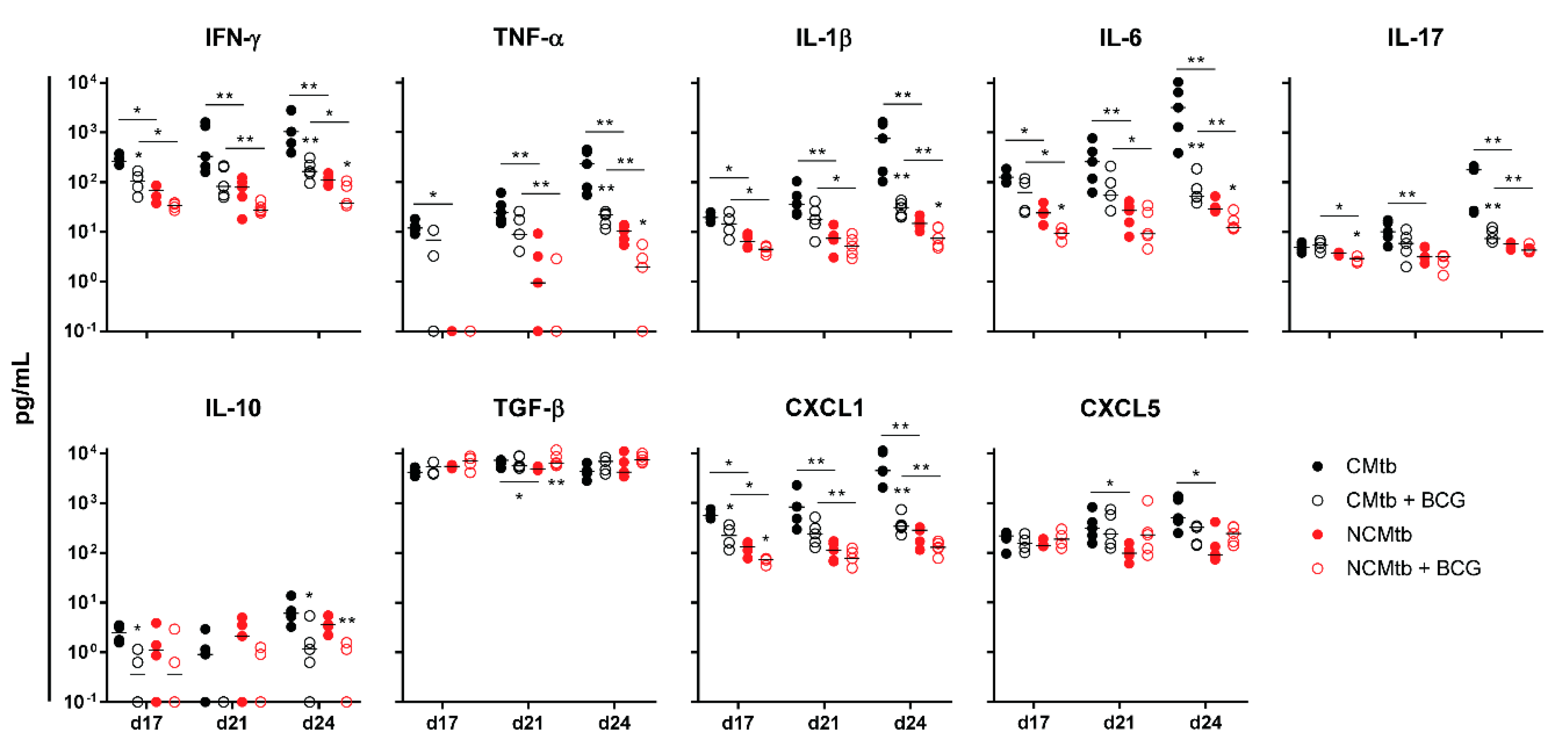
3.3. BCG Vaccination Protects Against TB Reducing the Inflammatory Response
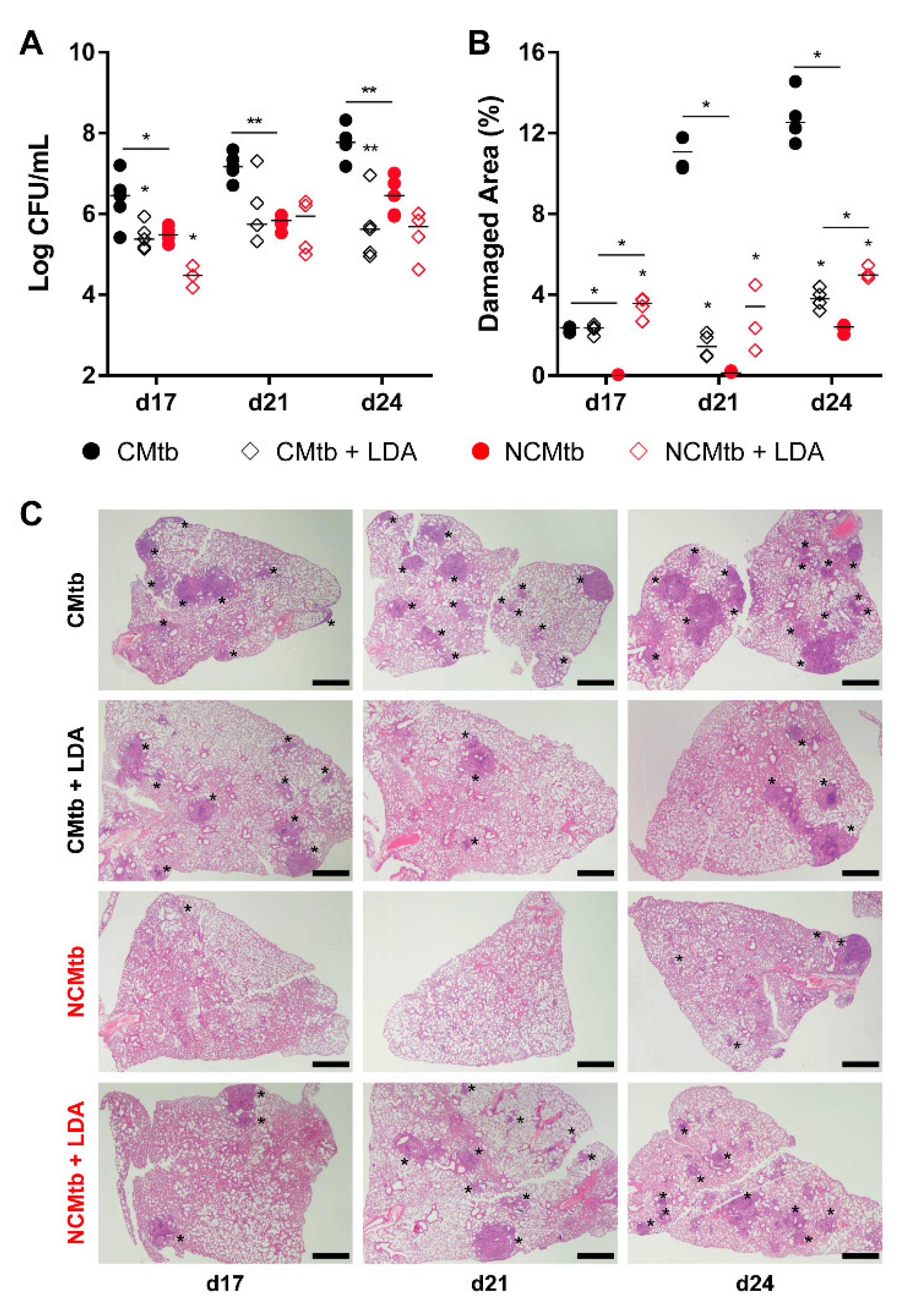
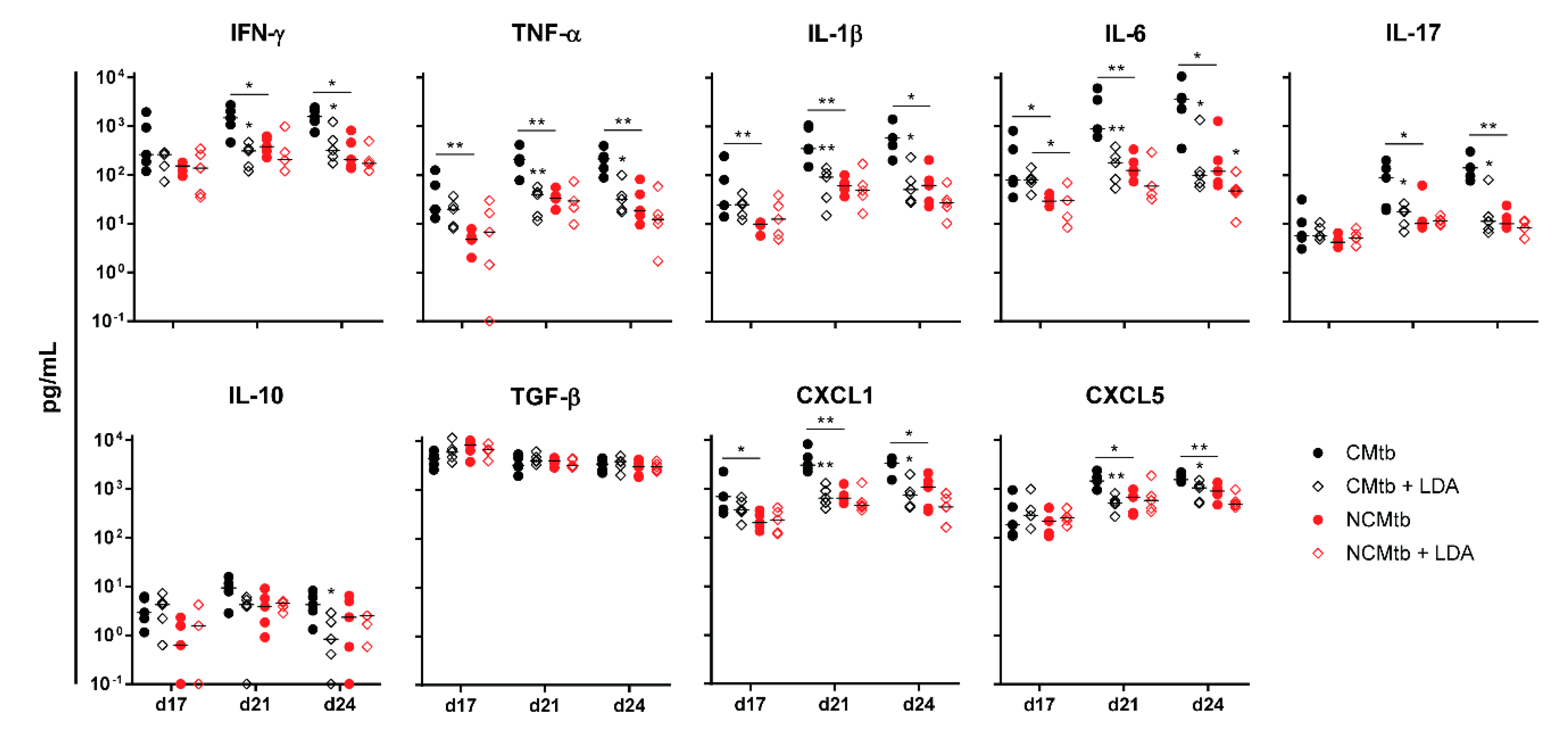
3.4. LDA Protects Against TB Progression after CMtb Challenge
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO Global Tuberculosis Report 2018; WHO: Geneva, Switzerland, 2018.
- Cardona, P.-J.; Català, M.; Arch, M.; Arias, L.; Alonso, S.; Cardona, P.; López, D.; Vilaplana, C.; Prats, C. Can systems immunology lead tuberculosis eradication? Curr. Opin. Syst. Biol. 2018, 12, 53–60. [Google Scholar] [CrossRef][Green Version]
- Houben, R.M.G.J.; Dodd, P.J. The Global Burden of Latent Tuberculosis Infection: A Re-estimation Using Mathematical Modelling. PLoS Med. 2016, 13, e1002152. [Google Scholar] [CrossRef] [PubMed]
- Menzies, N.A.; Wolf, E.; Connors, D.; Bellerose, M.; Sbarra, A.N.; Cohen, T.; Hill, A.N.; Yaesoubi, R.; Galer, K.; White, P.J.; et al. Progression from latent infection to active disease in dynamic tuberculosis transmission models: A systematic review of the validity of modelling assumptions. Lancet Infect. Dis. 2018, 18, e228–e238. [Google Scholar] [CrossRef]
- Cardona, P.-J. Patogénesis de la tuberculosis y otras micobacteriosis. Enferm. Infecc. Microbiol. Clin. 2018, 36, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Cardona, P.-J. The key role of exudative lesions and their encapsulation: Lessons learned from the pathology of human pulmonary tuberculosis. Front. Microbiol. 2015, 6, 612. [Google Scholar] [CrossRef]
- Glickman, M.S. Cording, Cord Factors, and Trehalose Dimycolate. In The Mycobacterial Cell Envelope; Daffé, M., Reyrat, J.-M., Eds.; ASM Press: Washington, DC, USA, 2008; pp. 63–73. [Google Scholar]
- Middlebrook, G.; Dubos, R.J.; Pierce, C. Differential characteristics of virulent and avirulent variants of mammalian tubercle bacilli. J. Bacteriol. 1947, 54, 66. [Google Scholar]
- Brambilla, C.; Llorens-Fons, M.; Julián, E.; Noguera-Ortega, E.; Tomàs-Martínez, C.; Pérez-Trujillo, M.; Byrd, T.F.; Alcaide, F.; Luquin, M. Mycobacteria clumping increase their capacity to damage macrophages. Front. Microbiol. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Julián, E.; Roldán, M.; Sánchez-Chardi, A.; Astola, O.; Agustí, G.; Luquin, M. Microscopic cords, a virulence-related characteristic of Mycobacterium tuberculosis, are also present in nonpathogenic mycobacteria. J. Bacteriol. 2010, 192, 1751–1760. [Google Scholar] [CrossRef]
- Kalsum, S.; Braian, C.; Koeken, V.A.C.M.; Raffetseder, J.; Lindroth, M.; van Crevel, R.; Lerm, M. The Cording Phenotype of Mycobacterium tuberculosis Induces the Formation of Extracellular Traps in Human Macrophages. Front. Cell. Infect. Microbiol. 2017, 7, 278. [Google Scholar] [CrossRef]
- Mahamed, D.; Boulle, M.; Ganga, Y.; Mc Arthur, C.; Skroch, S.; Oom, L.; Catinas, O.; Pillay, K.; Naicker, M.; Rampersad, S.; et al. Intracellular growth of Mycobacterium tuberculosis after macrophage cell death leads to serial killing of host cells. Elife 2017, 6, e22028. [Google Scholar] [CrossRef]
- Guirado, E.; Gordillo, S.; Gil, O.; Díaz, J.; Tapia, G.; Vilaplana, C.; Ausina, V.; Cardona, P.-J. Intragranulomatous necrosis in pulmonary granulomas is not related to resistance against Mycobacterium tuberculosis infection in experimental murine models induced by aerosol. Int. J. Exp. Pathol. 2006, 87, 139–149. [Google Scholar] [CrossRef]
- Irwin, S.M.; Driver, E.; Lyon, E.; Schrupp, C.; Ryan, G.; Gonzalez-Juarrero, M.; Basaraba, R.J.; Nuermberger, E.L.; Lenaerts, A.J. Presence of multiple lesion types with vastly different microenvironments in C3HeB/FeJ mice following aerosol infection with Mycobacterium tuberculosis. Dis. Model. Mech. 2015, 8, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Kramnik, I.; Beamer, G. Mouse models of human TB pathology: Roles in the analysis of necrosis and the development of host-directed therapies. Semin. Immunopathol. 2016, 38, 221–237. [Google Scholar] [CrossRef]
- Marzo, E.; Vilaplana, C.; Tapia, G.; Diaz, J.; Garcia, V.; Cardona, P.-J. Damaging role of neutrophilic infiltration in a mouse model of progressive tuberculosis. Tuberculosis 2014, 94, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Caceres, N.; Vilaplana, C.; Prats, C.; Marzo, E.; Llopis, I.; Valls, J.; Lopez, D.; Cardona, P.J. Evolution and role of corded cell aggregation in Mycobacterium tuberculosis cultures. Tuberculosis 2013, 93, 690–698. [Google Scholar] [CrossRef]
- Arias, L.; Goig, G.A.; Cardona, P.; Torres-Puente, M.; Díaz, J.; Rosales, Y.; Garcia, E.; Tapia, G.; Comas, I.; Vilaplana, C.; et al. Influence of Gut Microbiota on Progression to Tuberculosis Generated by High Fat Diet-Induced Obesity in C3HeB/FeJ Mice. Front. Immunol. 2019, 10, 2464. [Google Scholar] [CrossRef]
- Cardona, P.-J. What we have learned and what we have missed in tuberculosis pathophysiology for a new vaccine design: Searching for the “pink swan. ” Front. Immunol. 2017, 8, 556. [Google Scholar] [CrossRef] [PubMed]
- Gil, O.; Díaz, I.; Vilaplana, C.; Tapia, G.; Díaz, J.; Fort, M.; Cáceres, N.; Pinto, S.; Caylà, J.; Corner, L.; et al. Granuloma encapsulation is a key factor for containing tuberculosis infection in minipigs. PLoS ONE 2010, 5, e10030. [Google Scholar] [CrossRef]
- Prats, C.; Vilaplana, C.; Valls, J.; Marzo, E.; Cardona, P.-J.; López, D. Local inflammation, dissemination and coalescence of lesions are key for the progression toward active tuberculosis: The bubble model. Front. Microbiol. 2016, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.; Lee, J.; Ren, F.; Chen, M.; Kornfeld, H.; Remold, H.G. Mycobacterium tuberculosis blocks crosslinking of annexin-1 and apoptotic envelope formation on infected macrophages to maintain virulence. Nat. Immunol. 2008, 9, 1189–1197. [Google Scholar] [CrossRef]
- Lee, J.; Remold, H.G.; Ieong, M.H.; Kornfeld, H. Macrophage Apoptosis in Response to High Intracellular Burden of Mycobacterium tuberculosis Is Mediated by a Novel Caspase-Independent Pathway. J. Immunol. 2006, 176, 4267–4274. [Google Scholar] [CrossRef] [PubMed]
- Wood, R.; Morrow, C.; Barry, C.E.; Bryden, W.A.; Call, C.J.; Hickey, A.J.; Rodes, C.E.; Scriba, T.J.; Blackburn, J.; Issarow, C.; et al. Real-Time Investigation of Tuberculosis Transmission: Developing the Respiratory Aerosol Sampling Chamber (RASC). PLoS ONE 2016, 11, e0146658. [Google Scholar] [CrossRef] [PubMed]
- Sloot, R.; Schim van der Loeff, M.F.; Kouw, P.M.; Borgdorff, M.W. Risk of Tuberculosis after Recent Exposure. A 10-Year Follow-up Study of Contacts in Amsterdam. Am. J. Respir. Crit. Care Med. 2014, 190, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Trauer, J.M.; Moyo, N.; Tay, E.-L.; Dale, K.; Ragonnet, R.; McBryde, E.S.; Denholm, J.T. Risk of Active Tuberculosis in the Five Years Following Infection... 15%? Chest 2016, 149, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Cardona, P.-J.; Prats, C. The Small Breathing Amplitude at the Upper Lobes Favors the Attraction of Polymorphonuclear Neutrophils to Mycobacterium tuberculosis Lesions and Helps to Understand the Evolution toward Active Disease in An Individual-Based Model. Front. Microbiol. 2016, 7, 354. [Google Scholar] [CrossRef] [PubMed]
- Dockrell, H.M.; Smith, S.G. What have we learnt about BCG vaccination in the last 20 years? Front. Immunol. 2017, 8, 1134. [Google Scholar] [CrossRef]
- Heimbeck, J. Incidence of tuberculosis in young adult women with special reference to employment. Br. J. Tuberc. 1938, 32, 154–166. [Google Scholar] [CrossRef]
- Bloom, B.R. New Promise for Vaccines against Tuberculosis. N. Engl. J. Med. 2018, 379, 1672–1674. [Google Scholar] [CrossRef]
- Vilaplana, C.; Marzo, E.; Tapia, G.; Diaz, J.; Garcia, V.; Cardona, P.J. Ibuprofen therapy resulted in significantly decreased tissue bacillary loads and increased survival in a new murine experimental model of active tuberculosis. J. Infect. Dis. 2013, 208, 199–202. [Google Scholar] [CrossRef]
- Zumla, A.; Maeurer, M.; Chakaya, J.; Hoelscher, M.; Ntoumi, F.; Rustomjee, R.; Vilaplana, C.; Yeboah-Manu, D.; Rasolofo, V.; Munderi, P.; et al. Towards host-directed therapies for tuberculosis. Nat. Rev. Drug Discov. 2015, 14, 511–512. [Google Scholar] [CrossRef]
- Boro, M.; Balaji, K.N. CXCL1 and CXCL2 Regulate NLRP3 Inflammasome Activation via G-Protein–Coupled Receptor CXCR2. J. Immunol. 2017, 199, 1660–1671. [Google Scholar] [CrossRef] [PubMed]
- Lombard, R.; Doz, E.; Carreras, F.; Epardaud, M.; Le Vern, Y.; Buzoni-Gatel, D.; Winter, N. IL-17RA in non-hematopoietic cells controls CXCL-1 and 5 critical to recruit neutrophils to the lung of mycobacteria-infected mice during the adaptive immune response. PLoS ONE 2016, 11, e0149455. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cruz, A.; Ludovico, P.; Torrado, E.; Gama, J.B.; Sousa, J.; Gaifem, J.; Appelberg, R.; Rodrigues, F.; Cooper, A.M.; Pedrosa, J.; et al. IL-17A promotes intracellular growth of Mycobacterium by inhibiting apoptosis of infected macrophages. Front. Immunol. 2015, 6, 498. [Google Scholar] [CrossRef]
- Basile, J.I.; Geffner, L.J.; Romero, M.M.; Balboa, L.; Sabio Y García, C.; Ritacco, V.; García, A.; Cuffré, M.; Abbate, E.; López, B.; et al. Outbreaks of Mycobacterium Tuberculosis MDR strains induce high IL-17 T-cell response in patients with MDR tuberculosis that is closely associated with high antigen load. J. Infect. Dis. 2011, 204, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.M.; Ramakrishnan, L. The Role of the Granuloma in Expansion and Dissemination of Early Tuberculous Infection. Cell 2009, 136, 37–49. [Google Scholar] [CrossRef]
- Canetti, G. Exogenous reinfection and pulmonary tuberculosis a study of the pathology. Tubercle 1950, 31, 224–233. [Google Scholar] [CrossRef]
- Pottenger, F.M. Tuberculosis in the Child and the Adult. Arch. Intern. Med. 1935, 55, 169. [Google Scholar]
- Driver, E.R.; Ryan, G.J.; Hoff, D.R.; Irwin, S.M.; Basaraba, R.J.; Kramnik, I.; Lenaerts, A.J. Evaluation of a mouse model of necrotic granuloma formation using C3HeB/FeJ mice for testing of drugs against Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2012, 56, 3181–3195. [Google Scholar] [CrossRef]
- Rosenthal, I.M.; Tasneen, R.; Peloquin, C.A.; Zhang, M.; Almeida, D.; Mdluli, K.E.; Karakousis, P.C.; Grosset, J.H.; Nuermberger, E.L. Dose-ranging comparison of rifampin and rifapentine in two pathologically distinct murine models of tuberculosis. Antimicrob. Agents Chemother. 2012, 56, 4331–4340. [Google Scholar] [CrossRef]
- Lanoix, J.P.; Lenaerts, A.J.; Nuermberger, E.L. Heterogeneous disease progression and treatment response in a C3HeB/FeJ mouse model of tuberculosis. DMM Dis. Model. Mech. 2015, 8, 603–610. [Google Scholar] [CrossRef]
- North, R.J. Mycobacterium tuberculosis is strikingly more virulent for mice when given via the respiratory than via the intravenous route. J. Infect. Dis. 1995, 172, 1550–1553. [Google Scholar] [CrossRef] [PubMed]
- Cardona, P.J.; Cooper, A.; Luquín, M.; Ariza, A.; Filipo, F.; Orme, I.M.; Ausina, V. The intravenous model of murine tuberculosis is less pathogenic than the aerogenic model owing to a more rapid induction of systemic immunity. Scand. J. Immunol. 1999, 49, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.J.; LaCourse, R.; Ryan, L.; North, R.J. “Immunization” against airborne tuberculosis by an earlier primary response to a concurrent intravenous infection. Immunology 2008, 124, 514–521. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arias, L.; Cardona, P.; Català, M.; Campo-Pérez, V.; Prats, C.; Vilaplana, C.; Julián, E.; Cardona, P.-J. Cording Mycobacterium tuberculosis Bacilli Have a Key Role in the Progression towards Active Tuberculosis, Which is Stopped by Previous Immune Response. Microorganisms 2020, 8, 228. https://doi.org/10.3390/microorganisms8020228
Arias L, Cardona P, Català M, Campo-Pérez V, Prats C, Vilaplana C, Julián E, Cardona P-J. Cording Mycobacterium tuberculosis Bacilli Have a Key Role in the Progression towards Active Tuberculosis, Which is Stopped by Previous Immune Response. Microorganisms. 2020; 8(2):228. https://doi.org/10.3390/microorganisms8020228
Chicago/Turabian StyleArias, Lilibeth, Paula Cardona, Martí Català, Víctor Campo-Pérez, Clara Prats, Cristina Vilaplana, Esther Julián, and Pere-Joan Cardona. 2020. "Cording Mycobacterium tuberculosis Bacilli Have a Key Role in the Progression towards Active Tuberculosis, Which is Stopped by Previous Immune Response" Microorganisms 8, no. 2: 228. https://doi.org/10.3390/microorganisms8020228
APA StyleArias, L., Cardona, P., Català, M., Campo-Pérez, V., Prats, C., Vilaplana, C., Julián, E., & Cardona, P.-J. (2020). Cording Mycobacterium tuberculosis Bacilli Have a Key Role in the Progression towards Active Tuberculosis, Which is Stopped by Previous Immune Response. Microorganisms, 8(2), 228. https://doi.org/10.3390/microorganisms8020228