The interaction between Carbohydrates and the Antimicrobial Peptide P-113Tri is Involved in the Killing of Candida albicans


Abstract
1. Introduction
2. Materials and Methods
2.1. Antifungal Peptides and Reagents
2.2. C. albicans Strains and Growth Media
2.3. C. albicans Killing Assay
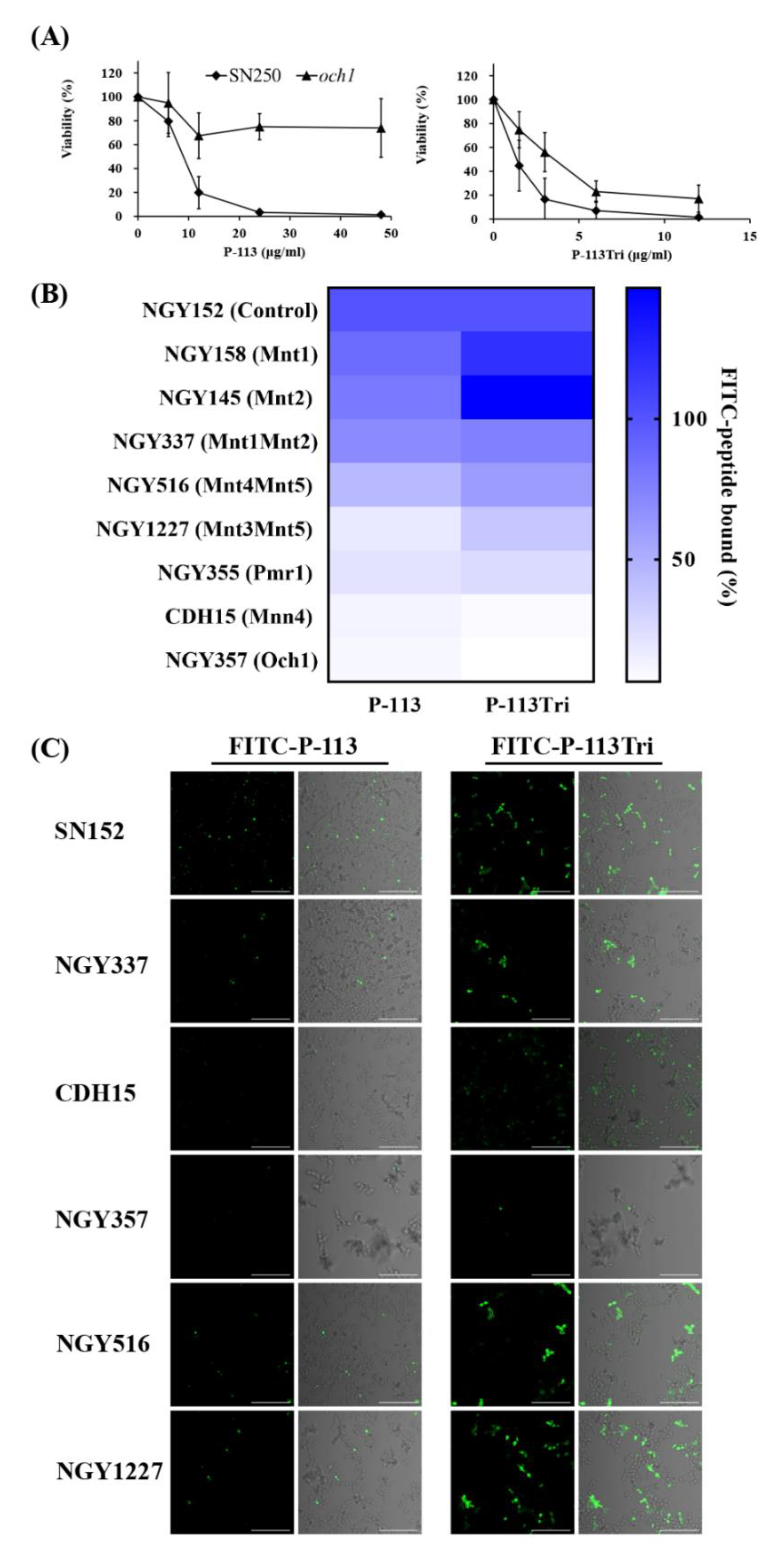
2.4. Measurement of Minimum Inhibitory Concentrations (MICs)
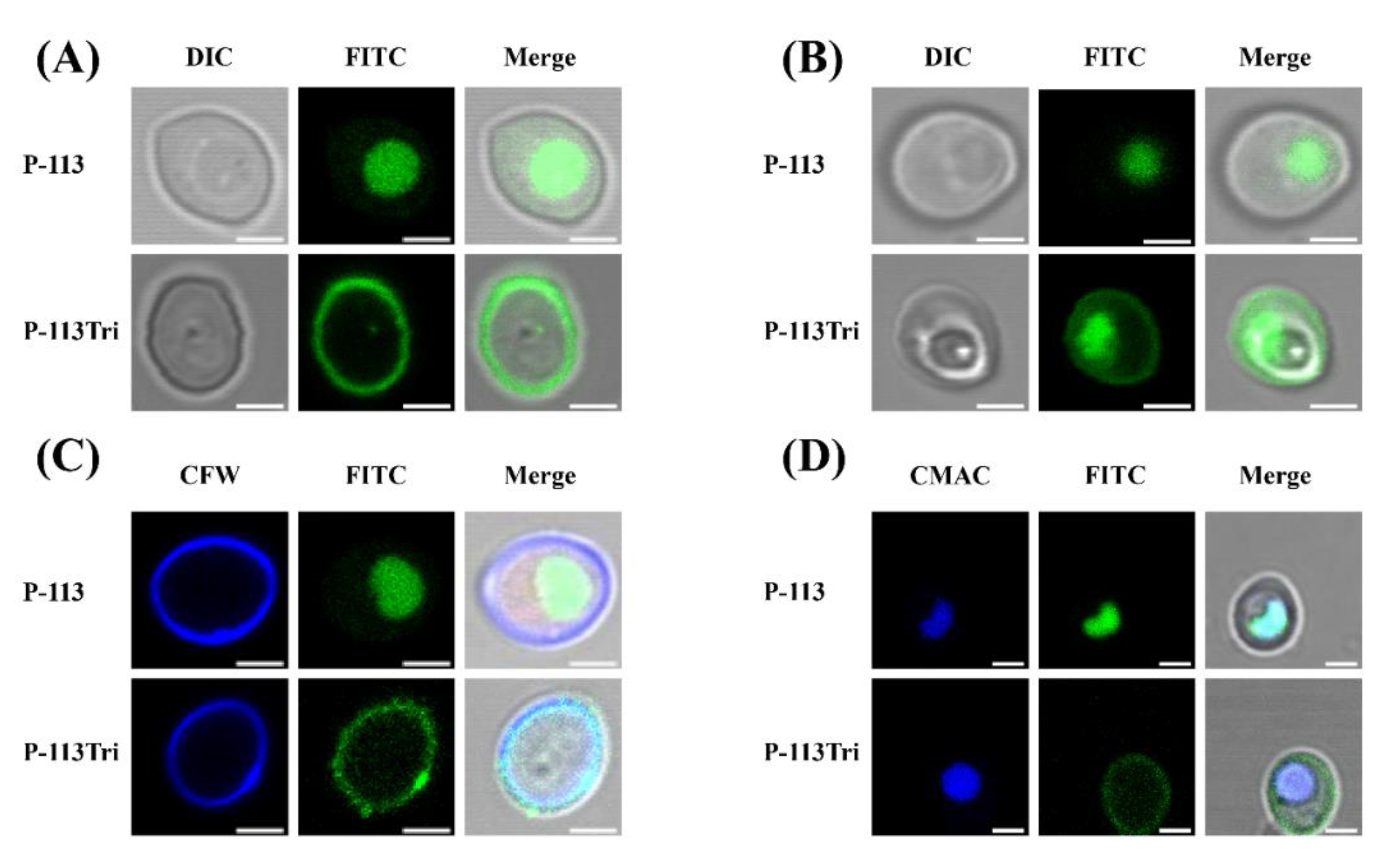
2.5. Confocal Scanning Laser Microscopy
2.6. Binding of P-113 and P-113Tri to C. albicans Cells
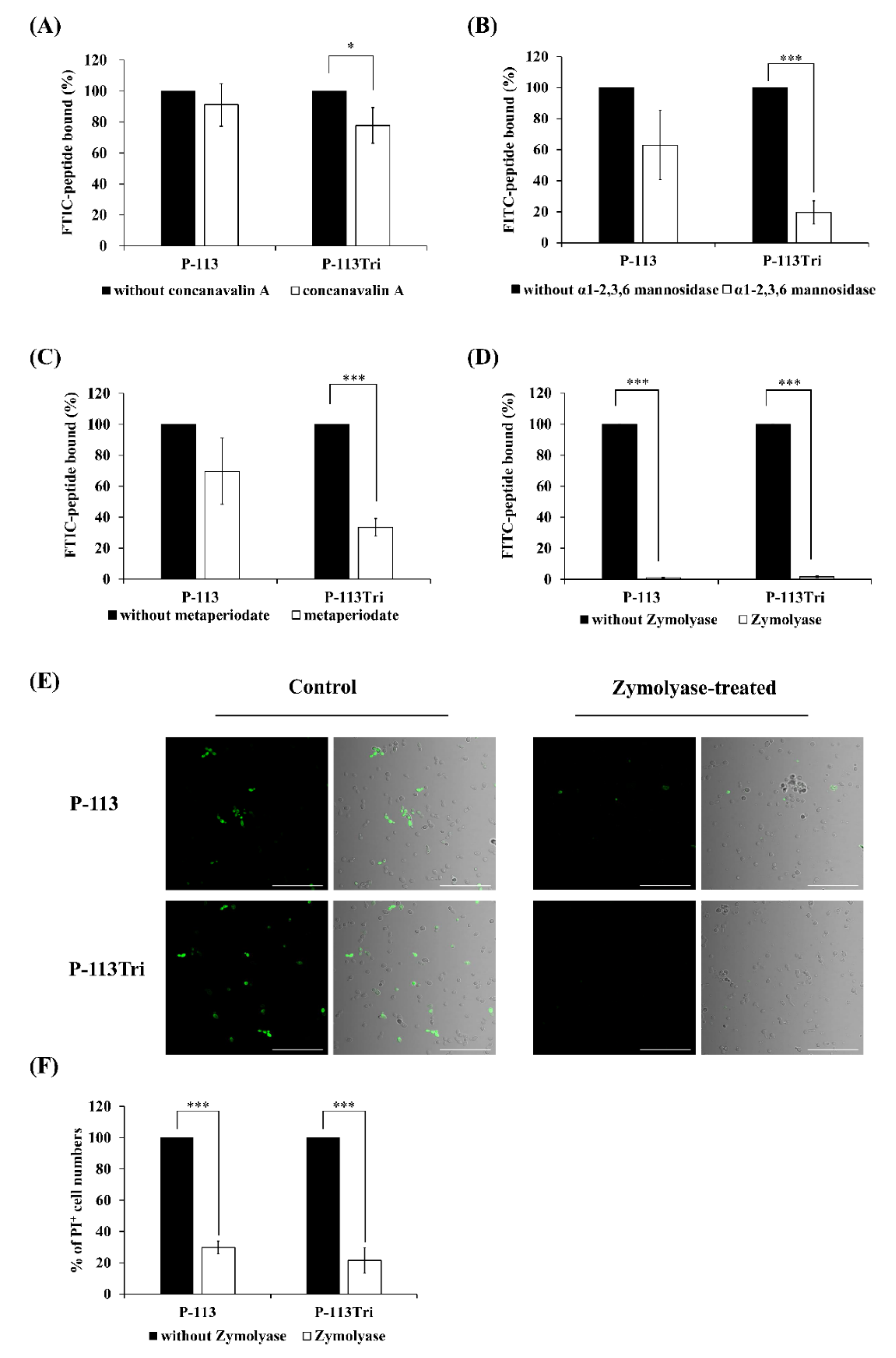
2.7. β-Glucan Staining
2.8. Competition Assays
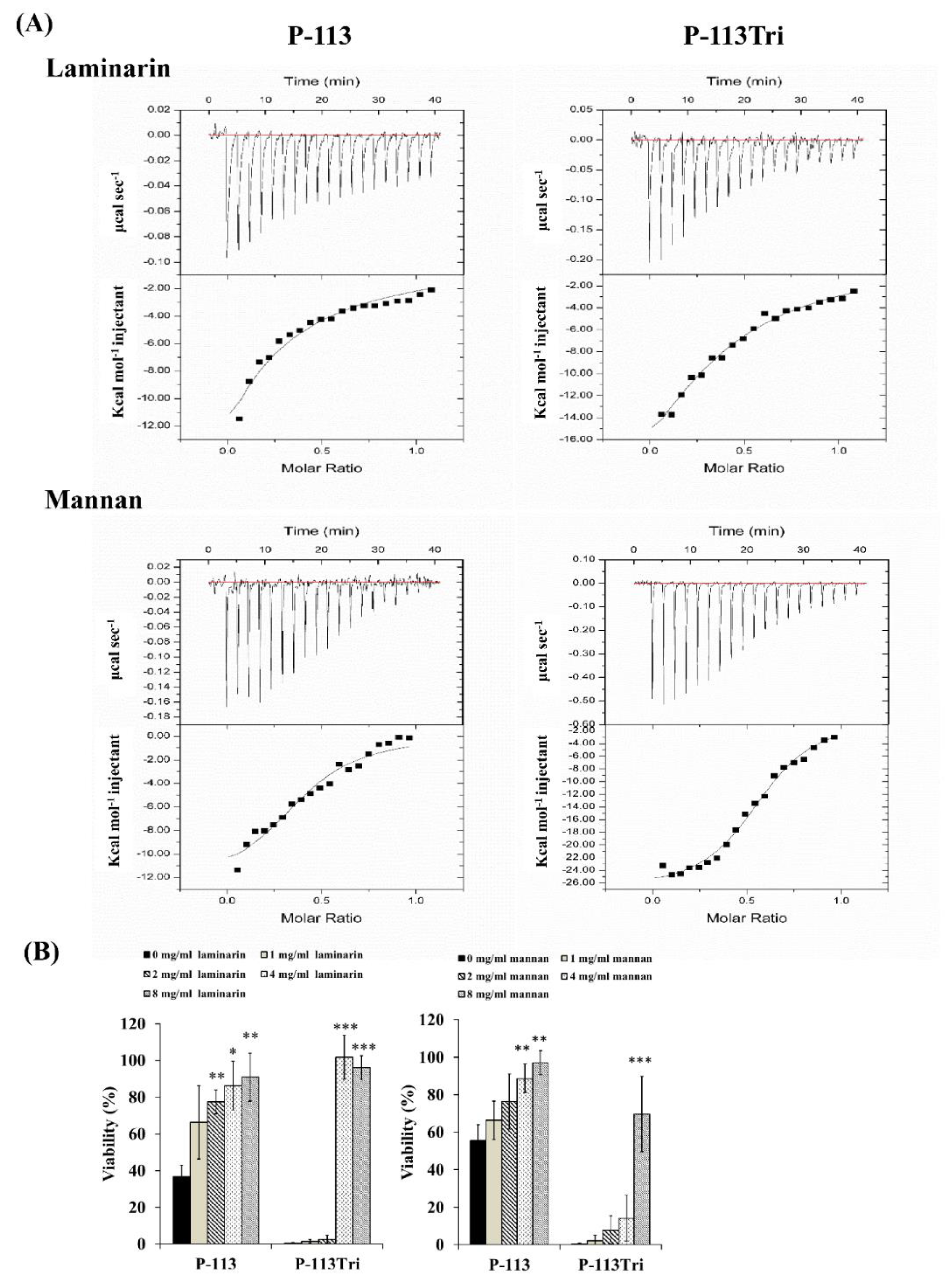
2.9. Measurement of the Dissociation Constants for the Peptide/Glycan Complexes
2.10. Glycan Microarray Analysis
2.11. Statistical Analysis
3. Results
3.1. P-113Tri Directly Interacts with the Cell Surface of C. albicans
3.2. P-113Tri Binding to C. albicans Cell Wall Carbohydrates Is Related to the Candidacidal Activity of the Peptide
3.3. Peptides Bind to N-Linked Mannan But Not O-Linked Mannan and Play a Partial Role in Candidacidal Activity
3.4. Binding to Phosphomannan Also Plays a Profound Role in The Activity of The Peptides
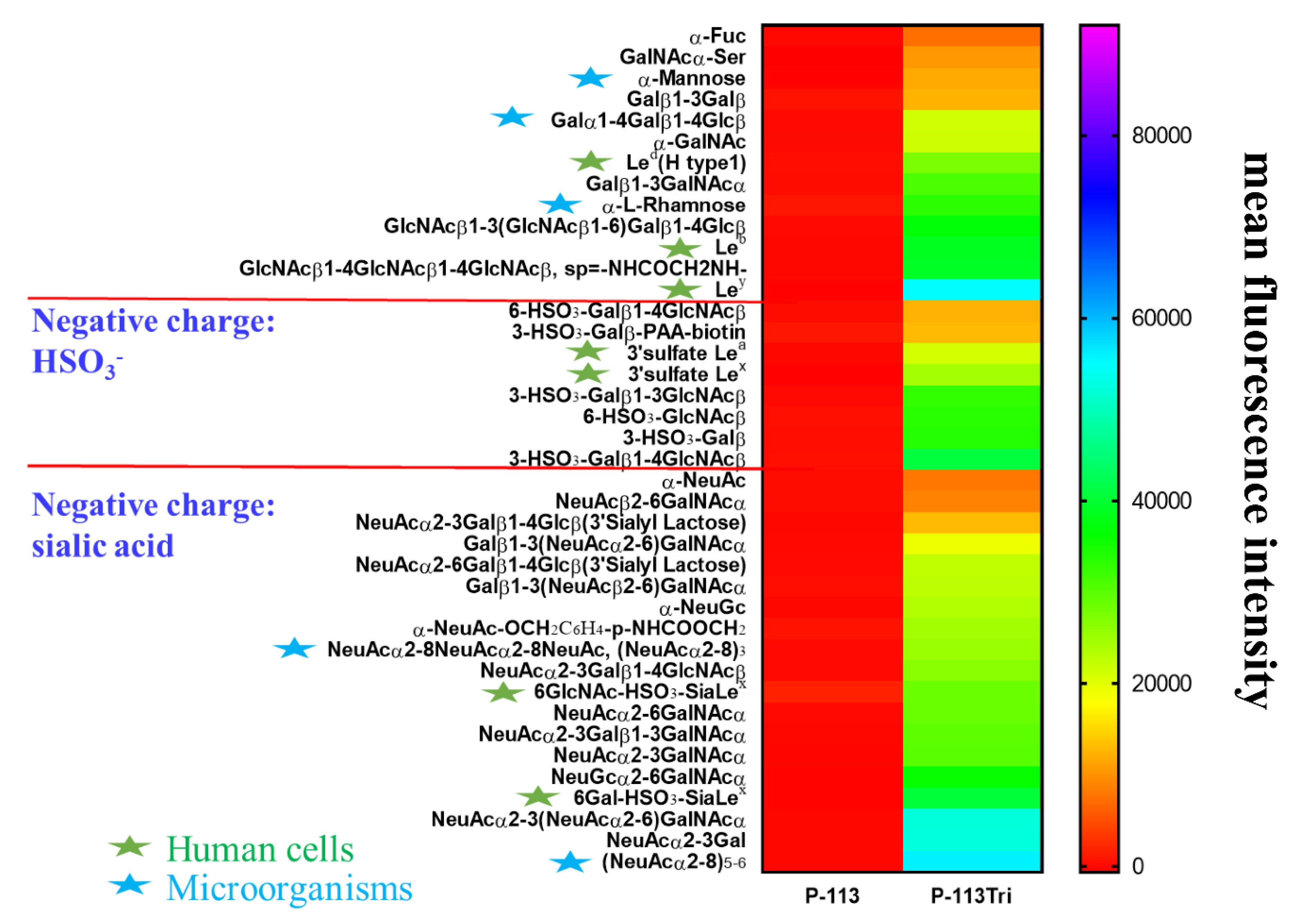
3.5. Screening the Potential Glycan Targets of P-113Tri by Glycan Array
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kullberg, B.J.; Arendrup, M.C. Invasive candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.; Wilson, D.; Drew, R.; Perfect, J. Azole antifungals: 35 years of invasive fungal infection management. Expert Rev. Anti. Infect. Ther. 2015, 13, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole antifungal resistance in Candida albicans and emerging non-albicans Candida species. Front. Microbiol. 2017, 7, 2173. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G.; Hultmark, D. Cell-free immunity in insects. Annu. Rev. Microbiol. 1987, 41, 103–126. [Google Scholar] [CrossRef]
- Fitton, J.E.; Dell, A.; Shaw, W.V. The amino acid sequence of the delta haemolysin of Staphylococcus aureus. FEBS Lett. 1980, 115, 209–212. [Google Scholar] [CrossRef]
- Park, S.; Park, S.H.; Ahn, H.C.; Kim, S.; Kim, S.S.; Lee, B.J.; Lee, B.J. Structural study of novel antimicrobial peptides, nigrocins, isolated from Rana nigromaculata. FEBS Lett. 2001, 507, 95–100. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial peptides from plants. Pharmaceuticals (Basel) 2015, 8, 711–757. [Google Scholar] [CrossRef]
- Seo, M.D.; Won, H.S.; Kim, J.H.; Mishig-Ochir, T.; Lee, B.J. Antimicrobial peptides for therapeutic applications: A review. Molecules (Basel) 2012, 17, 12276–12286. [Google Scholar] [CrossRef]
- Bondaryk, M.; Staniszewska, M.; Zielinska, P.; Urbanczyk-Lipkowska, Z. Natural antimicrobial peptides as inspiration for design of a new generation antifungal compounds. J. Fungi (Basel) 2017, 3, 46. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. Antimicrobial peptides: An emerging category of therapeutic agents. Front. Cell Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed]
- Nicola, A.M.; Albuquerque, P.; Paes, H.C.; Fernandes, L.; Costa, F.F.; Kioshima, E.S.; Abadio, A.K.R.; Bocca, A.L.; Felipe, M.S. Antifungal drugs: New insights in research & development. Pharmacol. Ther. 2019, 195, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Swidergall, M.; Ernst, J.F. Interplay between Candida albicans and the antimicrobial peptide armory. Eukaryot. Cell 2014, 13, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, D.M.; Spacciapoli, P.; Tran, L.T.; Xu, T.; Roberts, F.D.; Dalla Serra, M.; Buxton, D.K.; Oppenheim, F.G.; Friden, P. Anticandida activity is retained in P-113, a 12-amino-acid fragment of histatin 5. Antimicrob. Agents Chemother. 2001, 45, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Mickels, N.; McManus, C.; Massaro, J.; Friden, P.; Braman, V.; D’Agostino, R.; Oppenheim, F.; Warbington, M.; Dibart, S.; Van Dyke, T. Clinical and microbial evaluation of a histatin-containing mouthrinse in humans with experimental gingivitis. J. Clin. Periodontol. 2001, 28, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Helmerhorst, E.J.; Van’t Hof, W.; Veerman, E.C.; Simoons-Smit, I.; Nieuw Amerongen, A.V. Synthetic histatin analogues with broad-spectrum antimicrobial activity. Biochem. J. 1997, 326 (Pt 1), 39–45. [Google Scholar] [CrossRef]
- Jang, W.S.; Li, X.S.; Sun, J.N.; Edgerton, M. The P-113 fragment of histatin 5 requires a specific peptide sequence for intracellular translocation in Candida albicans, which is independent of cell wall binding. Antimicrob. Agents Chemother. 2008, 52, 497–504. [Google Scholar] [CrossRef]
- Yu, H.Y.; Tu, C.H.; Yip, B.S.; Chen, H.L.; Cheng, H.T.; Huang, K.C.; Lo, H.J.; Cheng, J.W. Easy strategy to increase salt resistance of antimicrobial peptides. Antimicrob. Agents Chemother. 2011, 55, 4918–4921. [Google Scholar] [CrossRef]
- Lin, G.Y.; Chen, H.F.; Xue, Y.P.; Yeh, Y.C.; Chen, C.L.; Liu, M.S.; Cheng, W.C.; Lan, C.Y. The antimicrobial peptides P-113Du and P-113Tri function against Candida albicans. Antimicrob. Agents Chemother. 2016, 60, 6369–6373. [Google Scholar] [CrossRef]
- Clinical Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts Approved Standard—Third Edition. CLSI Doc. M27-A3 Suppl. S. 2008, Volume 3, 6–12.
- Makrantoni, V.; Dennison, P.; Stark, M.J.; Coote, P.J. A novel role for the yeast protein kinase Dbf2p in vacuolar H+-ATPase function and sorbic acid stress tolerance. Microbiology (Reading, England) 2007, 153, 4016–4026. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.W.; Yang, C.Y.; Chang, H.T.; Lan, C.Y. Human antimicrobial peptide LL-37 inhibits adhesion of Candida albicans by interacting with yeast cell-wall carbohydrates. PLoS ONE 2011, 6, e17755. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Jyoti, M.A.; Song, H.Y.; Jang, W.S. Antifungal activity and action mechanism of Histatin 5-Halocidin hybrid peptides against Candida. ssp. PLoS ONE 2016, 11, e0150196. [Google Scholar] [CrossRef] [PubMed]
- Baranauskiene, L.; Petrikaite, V.; Matuliene, J.; Matulis, D. Titration calorimetry standards and the precision of isothermal titration calorimetry data. Int. J. Mol. Sci. 2009, 10, 2752–2762. [Google Scholar] [CrossRef]
- Chang, C.F.; Pan, J.F.; Lin, C.N.; Wu, I.L.; Wong, C.H.; Lin, C.H. Rapid characterization of sugar-binding specificity by in-solution proximity binding with photosensitizers. Glycobiology 2011, 21, 895–902. [Google Scholar] [CrossRef][Green Version]
- Hall, R.A.; Gow, N.A. Mannosylation in Candida albicans: Role in cell wall function and immune recognition. Mol Microbiol. 2013, 90, 1147–1161. [Google Scholar] [CrossRef]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. BioEssays 2006, 28, 799–808. [Google Scholar] [CrossRef]
- Sherrington, S.L.; Sorsby, E.; Mahtey, N.; Kumwenda, P.; Lenardon, M.D.; Brown, I.; Ballou, E.R.; MacCallum, D.M.; Hall, R.A. Adaptation of Candida albicans to environmental pH induces cell wall remodelling and enhances innate immune recognition. PLoS Pathog. 2017, 13, e1006403. [Google Scholar] [CrossRef]
- Noble, S.M.; Johnson, A.D. Strains and strategies for large-scale gene deletion studies of the diploid human fungal pathogen Candida. albicans. Eukaryot. Cell 2005, 4, 298–309. [Google Scholar] [CrossRef]
- Noble, S.M.; French, S.; Kohn, L.A.; Chen, V.; Johnson, A.D. Systematic screens of a Candida albicans homozygous deletion library decouple morphogenetic switching and pathogenicity. Nat. Genet. 2010, 42, 590–598. [Google Scholar] [CrossRef]
- Mora-Montes, H.M.; Bates, S.; Netea, M.G.; Castillo, L.; Brand, A.; Buurman, E.T.; Diaz-Jimenez, D.F.; Jan Kullberg, B.; Brown, A.J.; Odds, F.C.; et al. A multifunctional mannosyltransferase family in Candida albicans determines cell wall mannan structure and host-fungus interactions. J Biol Chem. 2010, 285, 12087–12095. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.; Hughes, H.B.; Munro, C.A.; Thomas, W.P.; MacCallum, D.M.; Bertram, G.; Atrih, A.; Ferguson, M.A.; Brown, A.J.; Odds, F.C.; et al. Outer chain N-glycans are required for cell wall integrity and virulence of Candida albicans. J. Biol. Chem. 2006, 281, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Munro, C.A.; Bates, S.; Buurman, E.T.; Hughes, H.B.; Maccallum, D.M.; Bertram, G.; Atrih, A.; Ferguson, M.A.; Bain, J.M.; Brand, A.; et al. Mnt1p and Mnt2p of Candida albicans are partially redundant alpha-1,2-mannosyltransferases that participate in O-linked mannosylation and are required for adhesion and virulence. J. Biol Chem. 2005, 280, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Hobson, R.P.; Munro, C.A.; Bates, S.; MacCallum, D.M.; Cutler, J.E.; Heinsbroek, S.E.; Brown, G.D.; Odds, F.C.; Gow, N.A. Loss of cell wall mannosylphosphate in Candida albicans does not influence macrophage recognition. J. Biol. Chem. 2004, 279, 39628–39635. [Google Scholar] [CrossRef]
- Bates, S.; MacCallum, D.M.; Bertram, G.; Munro, C.A.; Hughes, H.B.; Buurman, E.T.; Brown, A.J.; Odds, F.C.; Gow, N.A. Candida albicans Pmr1p, a secretory pathway P-type Ca2+/Mn2+-ATPase, is required for glycosylation and virulence. J. Biol. Chem. 2005, 280, 23408–23415. [Google Scholar] [CrossRef]
- Harris, M.; Mora-Montes, H.M.; Gow, N.A.; Coote, P.J. Loss of mannosylphosphate from Candida albicans cell wall proteins results in enhanced resistance to the inhibitory effect of a cationic antimicrobial peptide via reduced peptide binding to the cell surface. Microbiology (Reading, England) 2009, 155, 1058–1070. [Google Scholar] [CrossRef]
- Mora-Montes, H.M.; Bates, S.; Netea, M.G.; Diaz-Jimenez, D.F.; Lopez-Romero, E.; Zinker, S.; Ponce-Noyola, P.; Kullberg, B.J.; Brown, A.J.; Odds, F.C.; et al. Endoplasmic reticulum alpha-glycosidases of Candida albicans are required for N glycosylation, cell wall integrity, and normal host-fungus interaction. Eukaryot. Cell 2007, 6, 2184–2193. [Google Scholar] [CrossRef]
- Ibeas, J.I.; Lee, H.; Damsz, B.; Prasad, D.T.; Pardo, J.M.; Hasegawa, P.M.; Bressan, R.A.; Narasimhan, M.L. Fungal cell wall phosphomannans facilitate the toxic activity of a plant PR-5 protein. Plant J. 2000, 23, 375–383. [Google Scholar] [CrossRef]
- Koo, J.C.; Lee, B.; Young, M.E.; Koo, S.C.; Cooper, J.A.; Baek, D.; Lim, C.O.; Lee, S.Y.; Yun, D.J.; Cho, M.J. Pn-AMP1, a plant defense protein, induces actin depolarization in yeasts. Plant Cell Physiol. 2004, 45, 1669–1680. [Google Scholar] [CrossRef]
- Dielbandhoesing, S.K.; Zhang, H.; Caro, L.H.; van der Vaart, J.M.; Klis, F.M.; Verrips, C.T.; Brul, S. Specific cell wall proteins confer resistance to nisin upon yeast cells. Appl. Environ. Microbiol. 1998, 64, 4047–4052. [Google Scholar] [CrossRef]
- Edgerton, M.; Koshlukova, S.E.; Lo, T.E.; Chrzan, B.G.; Straubinger, R.M.; Raj, P.A. Candidacidal activity of salivary histatins. Identification of a histatin 5-binding protein on Candida albicans. J. Biol. Chem. 1998, 273, 20438–20447. [Google Scholar] [CrossRef] [PubMed]
- Peschel, A.; Otto, M.; Jack, R.W.; Kalbacher, H.; Jung, G.; Gotz, F. Inactivation of the dlt operon in Staphylococcus aureus confers sensitivity to defensins, protegrins, and other antimicrobial peptides. J. Biol. Chem. 1999, 274, 8405–8410. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Munch, D.; and Sahl, H.G. Structural variations of the cell wall precursor lipid II in Gram-positive bacteria-Impact on binding and efficacy of antimicrobial peptides. Biochim. Biophys. Acta. 2015, 1848, 3062–3071. [Google Scholar] [CrossRef] [PubMed]
- de Leeuw, E.; Li, C.; Zeng, P.; Li, C.; Diepeveen-de Buin, M.; Lu, W.Y.; Breukink, E.; Lu, W. Functional interaction of human neutrophil peptide-1 with the cell wall precursor lipid II. FEBS Lett. 2010, 584, 1543–1548. [Google Scholar] [CrossRef] [PubMed]
- De Lucca, A.J.; Walsh, T.J. Antifungal peptides: Novel therapeutic compounds against emerging pathogens. Antimicrob. Agents Chemother 1999, 43, 1–11. [Google Scholar] [CrossRef]
- Kim, M.K.; Park, H.S.; Kim, C.H.; Park, H.M.; Choi, W. Inhibitory effect of nikkomycin Z on chitin synthases in Candida albicans. Yeast 2002, 19, 341–349. [Google Scholar] [CrossRef]
- Jang, W.S.; Bajwa, J.S.; Sun, J.N.; Edgerton, M. Salivary histatin 5 internalization by translocation, but not endocytosis, is required for fungicidal activity in Candida albicans. Mol. Microbiol. 2010, 77, 354–370. [Google Scholar] [CrossRef]
- Xue, Y.P.; Kao, M.C.; Lan, C.Y. Novel mitochondrial complex I-inhibiting peptides restrain NADH dehydrogenase activity. Sci. Rep. 2019, 9, 13694. [Google Scholar] [CrossRef]
- Lowman, D.W.; Ensley, H.E.; Greene, R.R.; Knagge, K.J.; Williams, D.L.; Kruppa, M.D. Mannan structural complexity is decreased when Candida albicans is cultivated in blood or serum at physiological temperature. Carbohydr. Res. 2011, 346, 2752–2759. [Google Scholar] [CrossRef]
- Graus, M.S.; Wester, M.J.; Lowman, D.W.; Williams, D.L.; Kruppa, M.D.; Martinez, C.M.; Young, J.M.; Pappas, H.C.; Lidke, K.A.; Neumann, A.K. Mannan molecular substructures control nanoscale glucan exposure in Candida. Cell Rep. 2018, 24, 2432–2442. [Google Scholar] [CrossRef] [PubMed]
- Le Pendu, J.; Marionneau, S.; Cailleau-Thomas, A.; Rocher, J.; Le Moullac-Vaidye, B.; Clement, M. ABH and Lewis histo-blood group antigens in cancer. APMIS 2001, 109, 9–31. [Google Scholar] [CrossRef] [PubMed]
- Chaffin, W.L.; Lopez-Ribot, J.L.; Casanova, M.; Gozalbo, D.; Martinez, J.P. Cell wall and secreted proteins of Candida albicans: Identification, function, and expression. MMBR 1998, 62, 130–180. [Google Scholar] [CrossRef] [PubMed]
- Sandini, S.; La Valle, R.; De Bernardis, F.; Macri, C.; Cassone, A. The 65 kDa mannoprotein gene of Candida albicans encodes a putative beta-glucanase adhesin required for hyphal morphogenesis and experimental pathogenicity. Cell Microbiol. 2007, 9, 1223–1238. [Google Scholar] [CrossRef] [PubMed]
- Li, X.S.; Reddy, M.S.; Baev, D.; Edgerton, M. Candida albicans Ssa1/2p is the cell envelope binding protein for human salivary histatin 5. J. Biol Chem. 2003, 278, 28553–28561. [Google Scholar] [CrossRef]
- Navarro-Arias, M.J.; Hernandez-Chavez, M.J.; Garcia-Carnero, L.C.; Amezcua-Hernandez, D.G.; Lozoya-Perez, N.E.; Estrada-Mata, E.; Martinez-Duncker, I.; Franco, B.; Mora-Montes, H.M. Differential recognition of Candida tropicalis, Candida guilliermondii, Candida krusei, and Candida auris by human innate immune cells. Infect. Drug Resist. 2019, 12, 783–794. [Google Scholar] [CrossRef]
- Ma, Q.Q.; Lv, Y.F.; Gu, Y.; Dong, N.; Li, D.S.; Shan, A.S. Rational design of cationic antimicrobial peptides by the tandem of leucine-rich repeat. Amino Acids 2013, 44, 1215–1224. [Google Scholar] [CrossRef]
- Netea, M.G.; Brown, G.D.; Kullberg, B.J.; Gow, N.A. An integrated model of the recognition of Candida albicans by the innate immune system. Nat. Rev. Microbiol. 2008, 6, 67–78. [Google Scholar] [CrossRef]
- Mariano, F.S.; Campanelli, A.P.; Nociti, F.H., Jr.; Mattos-Graner, R.O.; Goncalves, R.B. Antimicrobial peptides and nitric oxide production by neutrophils from periodontitis subjects. Braz. J. Med. Biol. Res. 2012, 45, 1017–1024. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Systematic Name | Gene Name | Description |
|---|---|---|---|
| 1 | orf19.7590 | Putative NADH-ubiquinone oxidoreductase | |
| 2 | orf19.7247 | RIM101 | Transcription factor |
| 3 | orf19.7391 | OCH1 | α-1,6-mannosyltransferase |
| 4 | orf19.13191 | SNF4 | Putative subunit of the AMP-activated Snf1p kinase |
| 5 | orf19.287 | NUO2 | NADH-ubiquinone oxidoreductase subunit |
| 6 | orf19.1625 | Putative ubiquinone oxidoreductase | |
| 7 | orf19.1710 | ALI1 | Putative NADH-ubiquinone oxidoreductase |
| 8 | orf19.2570 | MCI4 | Putative NADH-ubiquinone dehydrogenase |
| 9 | orf19.2821 | Protein of unknown function | |
| 10 | orf19.4758 | Putative reductase or dehydrogenase | |
| 11 | orf19.5547 | Protein of unknown function | |
| 12 | orf19.3995 | RIM13 | Protease of the pH-response pathway |
| 13 | orf19.4755 | KEX2 | Subtilisin-like protease |
| 14 | orf19.5068 | IRE1 | Putative protein kinase |
| 15 | orf19.6293 | EMP24 | COPII-coated vesicle component |
| MIC90 (μg/mL) a in LYM broth | ||
|---|---|---|
| P-113 | P-113Tri | |
| O-linked Glycosylation Mutants | ||
| CAF2-1 (Control) | 6 | 1.5 |
| SPCa2 (Pmt1) | ND b | ND b |
| SPCa4 (Pmt2) | 6 | 1.5 |
| SPCa6 (Pmt4) | 6 | 3 |
| SPCa8 (Pmt6) | 6 | 3 |
| SPCa10 (Pmt5) | 12 | 3 |
| NGY152 (Control) | 12 | 3 |
| NGY145 (Mnt2) | 24 | 3 |
| NGY158 (Mnt1) | 6 | 1.5 |
| NGY337 (Mnt1Mnt2) | 12 | 6 |
| N-linked glycosylation mutants | ||
| NGY152 (Control) | 12 | 3 |
| NGY516 (Mnt4Mnt5) | 24 | 1.5 |
| NGY1227 (Mnt3Mnt5) | >24 | 1.5 |
| NGY357 (Och1) | >24 | 12 |
| CDH15 (Mnn4) | 24 | >12 |
| O and N-linked glycosylation mutants | ||
| NGY355 (Pmr1) | 3 | 3 |
| MIC90 (μg/mL) a in LYM broth | ||
|---|---|---|
| P-113 | P-113Tri | |
| Control | 3 | 0.75 |
| 5 mM Glucosamine 6-phosphate | 6 | 1.5 |
| 10 mM Glucosamine 6-phosphate | 12 | 1.5 |
| 5 mg/mL monosaccharide mannose 6-phosphate-BSA | >24 | 12 |
| 5 mg/mL trisaccharide mannose 6-phosphate-BSA | 24 | 6 |
| 5 mg/mL pentasaccharide mannose 6-phosphate-BSA | >24 | 6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, G.-Y.; Chang, C.-F.; Lan, C.-Y. The interaction between Carbohydrates and the Antimicrobial Peptide P-113Tri is Involved in the Killing of Candida albicans. Microorganisms 2020, 8, 299. https://doi.org/10.3390/microorganisms8020299
Lin G-Y, Chang C-F, Lan C-Y. The interaction between Carbohydrates and the Antimicrobial Peptide P-113Tri is Involved in the Killing of Candida albicans. Microorganisms. 2020; 8(2):299. https://doi.org/10.3390/microorganisms8020299
Chicago/Turabian StyleLin, Guan-Yu, Chuan-Fa Chang, and Chung-Yu Lan. 2020. "The interaction between Carbohydrates and the Antimicrobial Peptide P-113Tri is Involved in the Killing of Candida albicans" Microorganisms 8, no. 2: 299. https://doi.org/10.3390/microorganisms8020299
APA StyleLin, G.-Y., Chang, C.-F., & Lan, C.-Y. (2020). The interaction between Carbohydrates and the Antimicrobial Peptide P-113Tri is Involved in the Killing of Candida albicans. Microorganisms, 8(2), 299. https://doi.org/10.3390/microorganisms8020299