Plasmid-Mediated Colistin Resistance (mcr-1) in Escherichia coli from Non-Imported Fresh Vegetables for Human Consumption in Portugal


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolate
2.2. Antimicrobial Susceptibility Testing
2.3. Transfer Experiments
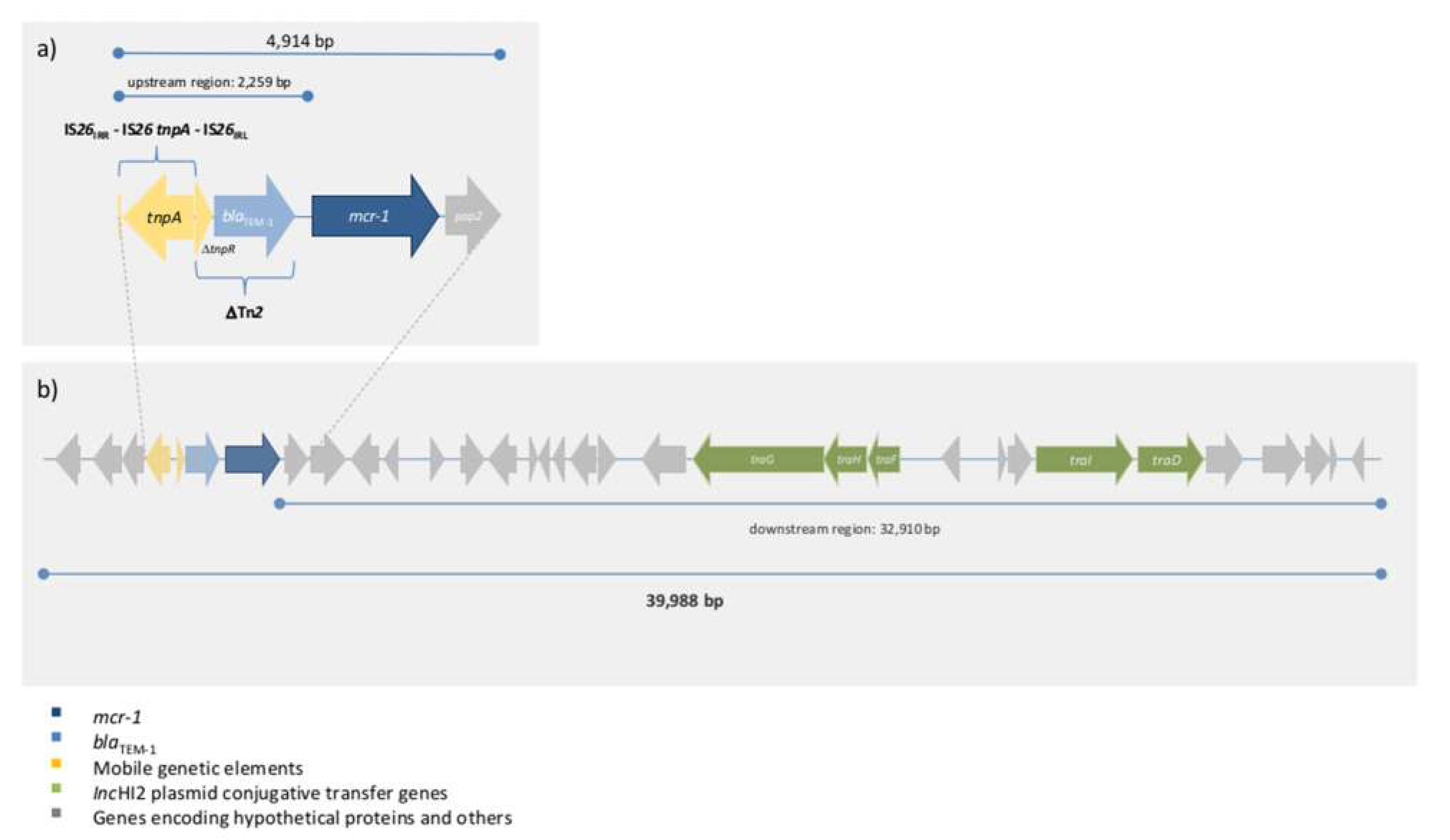
2.4. Genetic Background of E. coli INSali25
2.5. Genomic Epidemiological Analysis
3. Results and Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Paterson, D.L.; Harris, P.N. Colistin resistance: A major breach in our last line of defence. Lancet Infect. Dis. 2016, 16, 132–133. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L. Plasmid-mediated colistin resistance: An additional antibiotic resistance menace. Clin. Microbiol. Infect. 2016, 22, 398–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Zhang, H.; Liu, Y.-H.; Feng, Y. Towards understanding MCR-like colistin resistance. Trends Microbiol. 2018, 26, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Hölzel, C.S.; Tetens, J.L.; Schwaiger, K. Unraveling the role of vegetables in spreading antimicrobial-resistant bacteria: A need for quantitative risk assessment. Foodborne Pathog. Dis. 2018, 15, 671–688. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Yao, X.; Lv, L.; Doi, Y.; Huang, X.; Huang, S.; Liu, J.H. Emergence of mcr-1 in Raoultella ornithinolytica and Escherichia coli isolates from retail vegetables in China. Antimicrob. Agents Chemother. 2017, 61, e01139-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurfuh, K.; Poirel, L.; Nordmann, P.; Nüesch-Inderbinen, M.; Hächler, H.; Stephan, R. Occurrence of the plasmid-borne mcr-1 colistin resistance gene in extended-spectrum-β-lactamase-producing Enterobacteriaceae in river water and imported vegetable samples in Switzerland. Antimicrob. Agents Chemother. 2016, 60, 2594–2595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.T.; Li, X.; Zhang, Q.; Shan, H.; Zou, M.; Song, F.J. Colistin-resistant mcr-positive Enterobacteriaceae in fresh vegetables, an increasing infectious threat in China. Int. J. Antimicrob. Agents 2019, 54, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Jones-Dias, D.; Manageiro, V.; Ferreira, E.; Barreiro, P.; Vieira, L.; Moura, I.B.; Caniça, M. Architecture of class 1, 2, and 3 integrons from Gram negative bacteria recovered among fruits and vegetables. Front. Microbiol. 2016, 7, 1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manageiro, V.; Clemente, L.; Graça, R.; Correia, I.; Albuquerque, T.; Ferreira, E.; Caniça, M. New insights into resistance to colistin and third-generation cephalosporins of Escherichia coli in poultry, Portugal: Novel blaCTX-M-166 and blaESAC genes. Int. J. Food Microbiol. 2017, 263, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Z.; Feng, Y. BacWGSTdb, a database for genotyping and source tracking bacterial pathogens. Nucleic Acids Res. 2016, 44, D682–D687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veldman, K.; van Essen-Zandbergen, A.; Rapallini, M.; Wit, B.; Heymans, R.; van Pelt, W.; Mevius, D. Location of colistin resistance gene mcr-1 in Enterobacteriaceae from livestock and meat. J. Antimicrob. Chemother. 2016, 71, 2340–2342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjadj, L.; Riziki, T.; Zhu, Y.; Li, J.; Diene, S.M.; Rolain, J.M. Study of mcr-1 gene-mediated colistin resistance in Enterobacteriaceae isolated from humans and animals in different countries. Genes (Basel) 2017, 8, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, C.N.; Sodha, S.V.; Shaw, R.K.; Griffin, P.M.; Pink, D.; Hand, P.; Frankel, G. Fresh fruit and vegetables as vehicles for the transmission of human pathogens. Environ. Microbiol. 2010, 12, 2385–2397. [Google Scholar] [CrossRef] [PubMed]
- Zurfluh, K.; Klumpp, J.; Nüesch-Inderbinen, M.; Stephan, R. Full-length nucleotide sequences of mcr-1-harboring plasmids isolated from extended-spectrum-β-lactamase-producing Escherichia coli isolates of different origins. Antimicrob. Agents Chemother. 2016, 60, 5589–5591. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| IncHI2-Type Plasmid (bp) | Strain (MLST 1) | Isolation Source/ Country/Year | Identity(%) | Alignment Length (bp) | Antimicrobial Resistance Gene 2 | Plasmid GenBank Acc. No. |
|---|---|---|---|---|---|---|
| pSE08-00436-1 (264914) | S. enterica 08-00436 (ST28) | Chicken skin/Germany/2008 | 99.997 | 32115 | aadA1, aadA2-type, aph(3’)-Ia-type, aph(3’’)-Ib-type, aph(6)-Id, aac(3)-IIa, blaTEM-1B, mcr-1.1, cml-type, catA1-type, sul1, sul2, sul3, tet(A), dfrA1-type | NZ_CP020493.1 |
| p1rc4-mcr1 (239098) | E. coli 1RC4 (ST155) | Hajj pilgrim Stool/France/2014 | 99.994 | 32115 | aadA1, aadA12-type, aph(3’)-Ia, aph(3’’)-Ib, aph(6)-Id, blaTEM-1B, mcr-1.1, mph(A), cml-type, floR-type, sul3-type, tet(A), dfrA4 | NZ_CM008266.1 |
| pSA186_MCR1 (241600) | E. coli SA186 (ST131) | Human patient urine/ Saudi Arabia (Riyadh)/2012 | 99.993 | 30522 | aadA1, aadA2-type, aph(3’)-Ia, aph(3’’)-Ib, aph(6)-Id, blaTEM-1B, mcr-1.1, mph(A), cml-type, floR-type, sul3-type, tet(A), dfrA14 | NZ_CP022735.1 |
| pRS571-MCR-1.1 (257270) | E. coli RS571 (ST648) | rectal swab/Bangladesh: Dhaka/2018 | 99.993 | 29648 | aadA1, aadA2-type, aac(3)-IId-type, aph(3’)-Ia, aph(3’’)-Ib, aph(6)-Id, blaTEM-1B, mcr-1.1, mph(A), cml-type, floR-type, sul3-type, tet(A), dfrA14 | NZ_CP034390.1 |
| pG3X16-2-2 (265575) | E. coli G3X16-2 (ST1196) | Human feces/China: Guangxi/2017 | 99.977 | 30043 | aadA22, aph(3’)-Ia-type, aph(3’’)-Ib, aph(6)-Id, aph(4)-Ia-type, aac(3)-IV, blaTEM-1B, blaCTX-M-65, mcr-1.1, oqxAB-type, Inu(F)-type, mph(A), floR-type, sul1-type, sul2, tet(A)-type, tet(M)-type | NZ_CP038139.1 |
| pMCR1_025943 (265538) | E. coli WCHEC025943 (ST410) | Sewage/China: Sichuan, Chengdu/2017 | 99.977 | 30043 | aadA22, aph(3’)-Ia-type, aph(3’’)-Ib, aph(6)-Id, aph(4)-Ia-type, aac(3)-IV, blaTEM-1B, blaCTX-M-65, mcr-1.1, oqxAB-type, Inu(F)-type, mph(A), floR-type, sul1, sul2, tet(A), tet(M)-type | NZ_CP027202.2 |
| E. coli Strain (MLST a) | Isolation Source/Country/Year | Antimicrobial Resistance Genes b | Plasmid (bp) | Plasmid Type | Accession Number | Reference |
|---|---|---|---|---|---|---|
| INSali25 (ST1716) | Lettuce/Portugal/2015 | aadA1, aac(3)-Iv, aph(4)-Ia,aph(6)-Ia, aph(6)-Id, blaTEM-1B, mcr-1.1, sul2, tet(A), floR-type | pINSali25-MCR (≈250000) | IncHI2/ST4 | LSRK00000000 | [8]; This study |
| CTX148 (untypable) | tomato | mcr-1.1 | pT-CTX148 (57764) | IncI2 | MK754161 | [7] |
| TO89 (ST713) | Tomato/China: Huimin/2017 | mcr-1.1 | pT-89 (∼33) | IncX4 | SRMK00000000 | [7] |
| SQB-1-1 (ST2705) | Romaine lettuce/China: Qingdao/2017 | aph(3’)-Ia, aac(3)-IV-type, aph(4)-Ia, aadA1, aadA2-type, blaCTX-M-14, mcr-1.1, fosA3, mdf(A)-type, mph(A), cmlA1-type, floR-type, tet(M)-type, sul2, sul3 | pSQB-1-1 (≈250000) | IncHI2/ST3 | SRML00000000 | [7] |
| H226B (ST167) | Cha-om imported from Thailand/2014 | mcr-1.1 | pH226B (209401) | IncHI1 | KX129784 | [6,14] |
| 2SK1 (ST4683) | Basil leaves imported from Vietnam/2014 | mcr-1, blaCTX-M-65 | - | - | - | [14] |
| HS20eCTX (ST795) | Lettuce/China: Guangzhou/2016 | aph(3’)-Ia, aac(3)-IV-type, aph(4)-Ia, aadA1, aadA2-type, blaCTX-M-14, mcr-1.1, fosA3, cmlA1-type, floR-type, sul2, sul3 | pHNHS20EC (250827) | IncHI2/ST3 | MF135536 | [5] |
| BS21Ectx (ST2505) | Lettuce/China: Guangzhou/2016 | mcr-1 | - (∼60) | IncI2 | - | [5] |
| 6BF21eCTX (ST69) | Tomato/China: Guangzhou/2016 | mcr-1, blaCTX-M-14, floR, fosA3, oqxAB | - (∼244) | IncHI2/ST3 | - | [5] |
| TS62CTX (ST156) | Lettuce/China: Guangzhou/2015 | mcr-1 | - (∼60) | IncI2 | - | [5] |
| 6HS20E (ST48) | Lettuce/China: Guangzhou/2016 | mcr-1 | - (∼33) | IncX4 | - | [5] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manageiro, V.; Jones-Dias, D.; Ferreira, E.; Caniça, M. Plasmid-Mediated Colistin Resistance (mcr-1) in Escherichia coli from Non-Imported Fresh Vegetables for Human Consumption in Portugal. Microorganisms 2020, 8, 429. https://doi.org/10.3390/microorganisms8030429
Manageiro V, Jones-Dias D, Ferreira E, Caniça M. Plasmid-Mediated Colistin Resistance (mcr-1) in Escherichia coli from Non-Imported Fresh Vegetables for Human Consumption in Portugal. Microorganisms. 2020; 8(3):429. https://doi.org/10.3390/microorganisms8030429
Chicago/Turabian StyleManageiro, Vera, Daniela Jones-Dias, Eugénia Ferreira, and Manuela Caniça. 2020. "Plasmid-Mediated Colistin Resistance (mcr-1) in Escherichia coli from Non-Imported Fresh Vegetables for Human Consumption in Portugal" Microorganisms 8, no. 3: 429. https://doi.org/10.3390/microorganisms8030429