Formate Utilization by the Crenarchaeon Desulfurococcus amylolyticus

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals
2.2. Microorganism and Medium Composition
2.3. Closed Batch Cultivations
2.4. Whole Cell Conversion Experiments
2.5. Cell Counting
2.6. Gas Chromatography
2.7. Formic Acid Analysis
2.8. HPLC
2.9. Data Analysis
2.10. Gibbs Free Energy Calculations
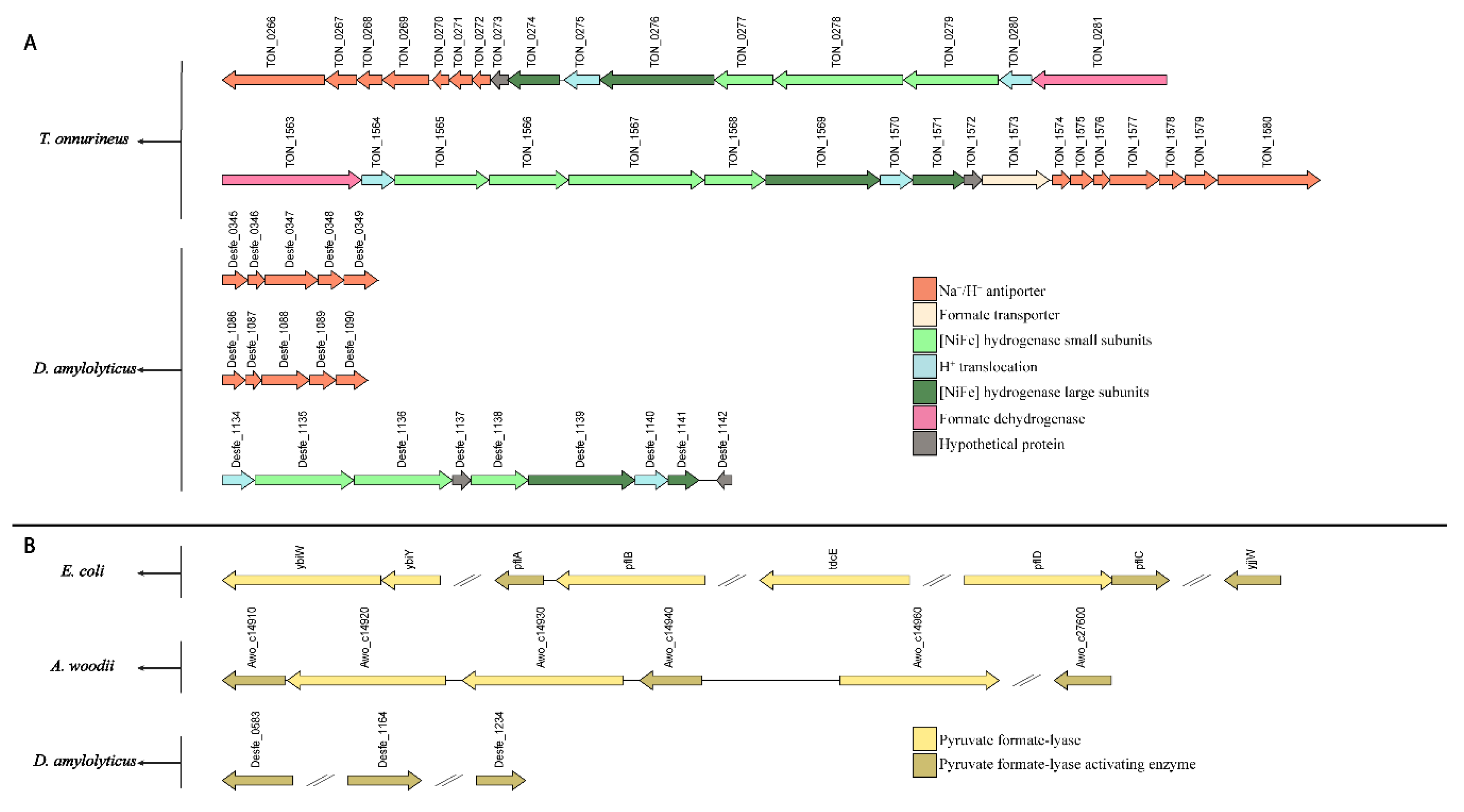
2.11. Genome Analysis
3. Results
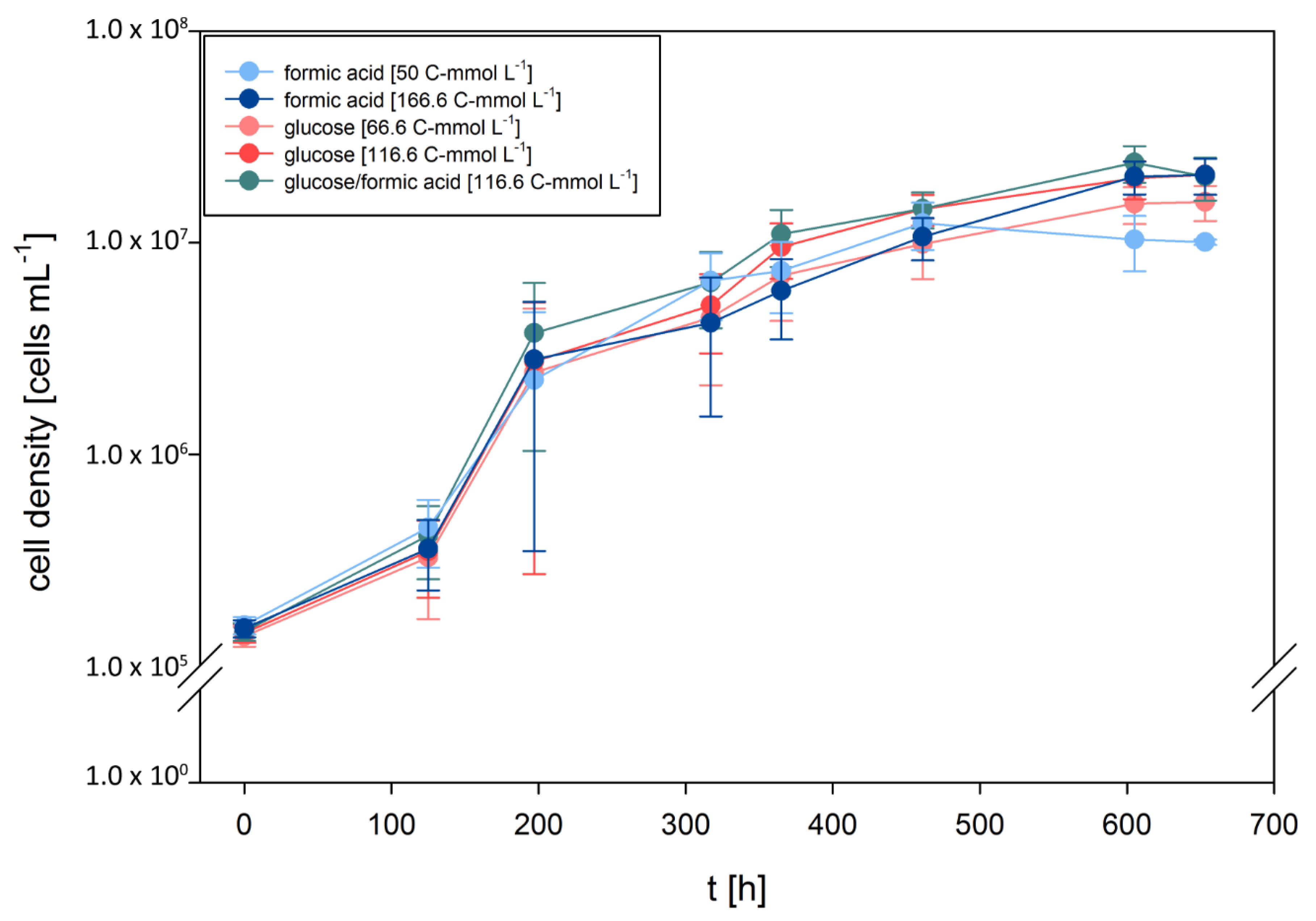
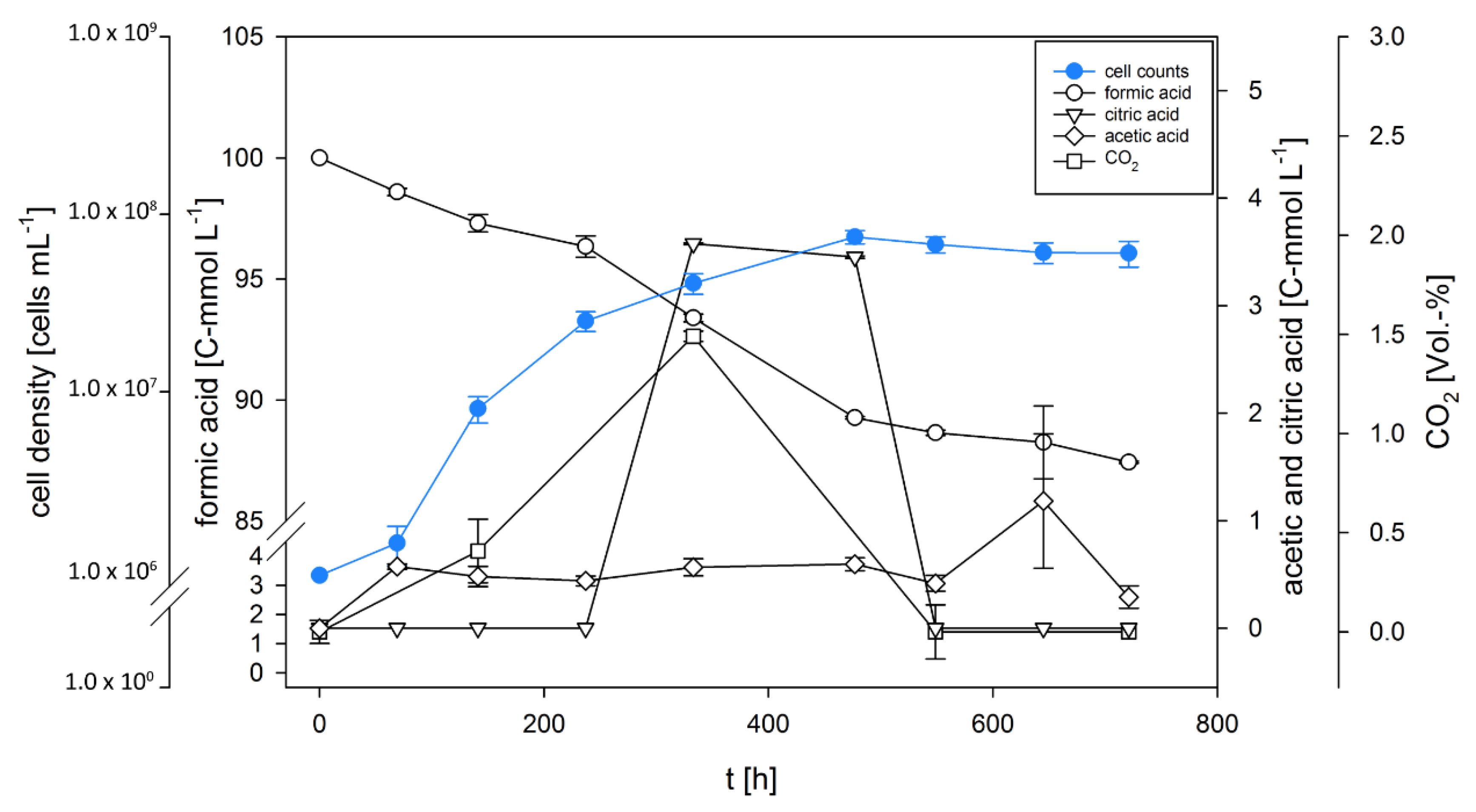
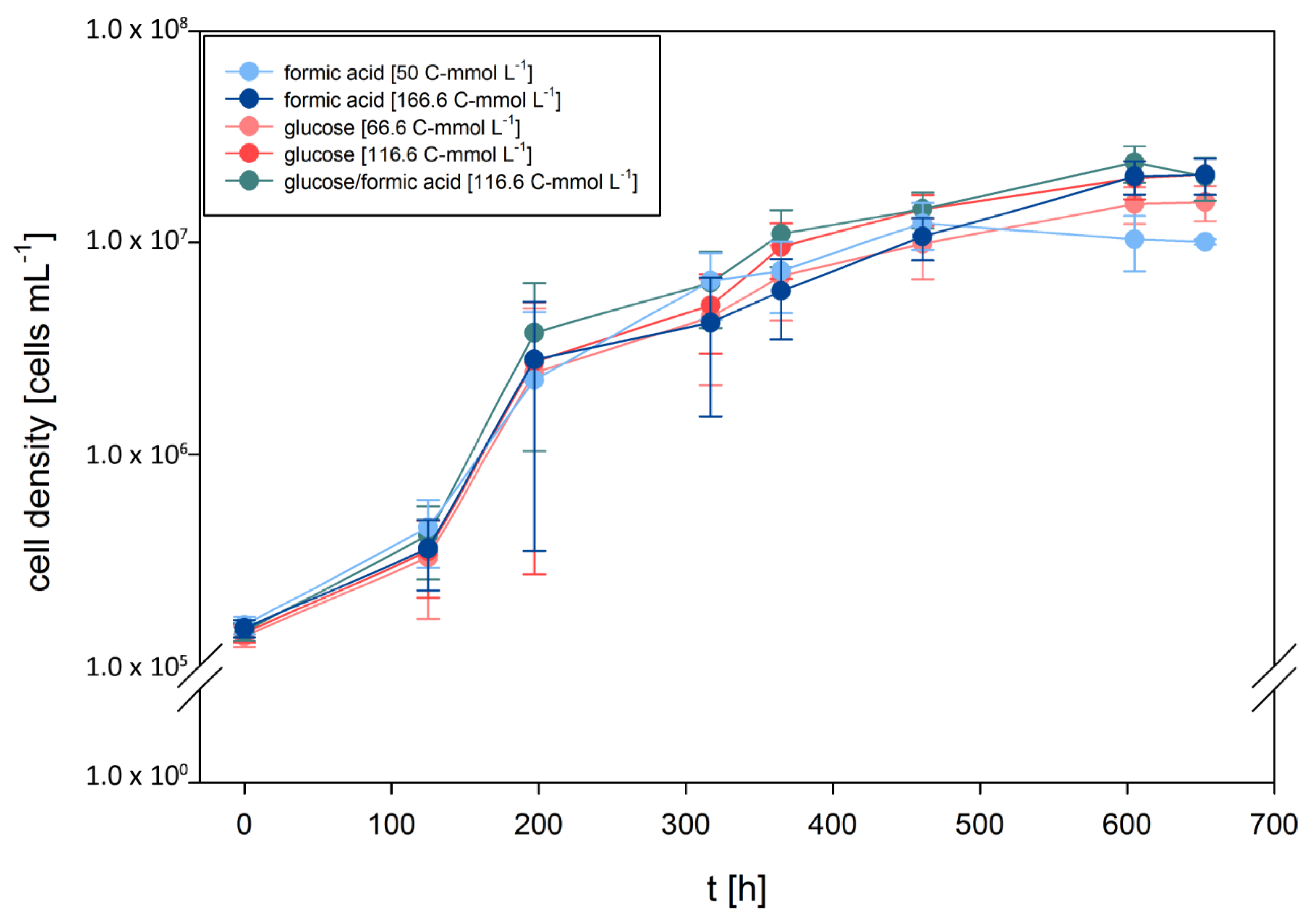
3.1. D. amylolyticus Grows on Formate
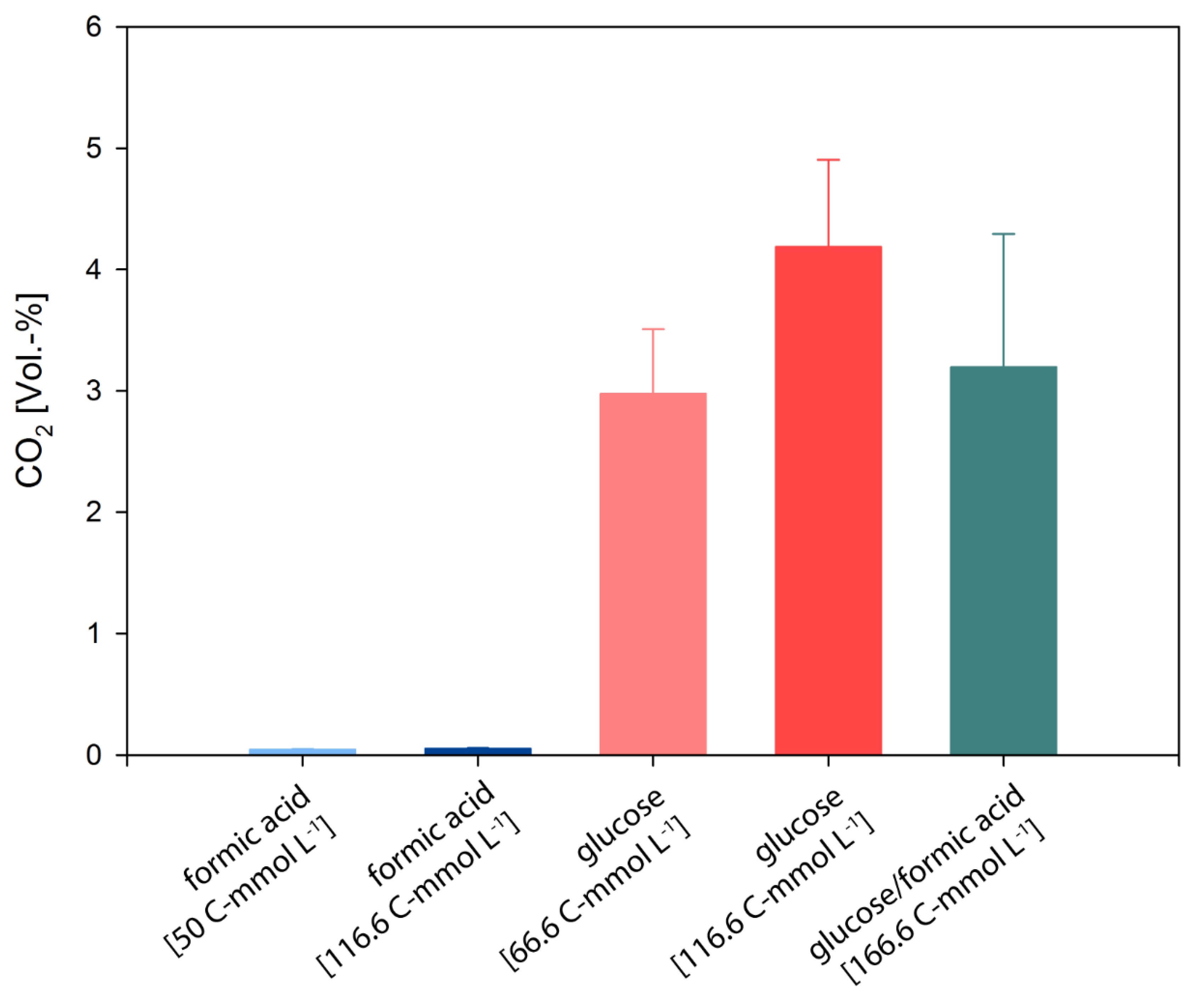
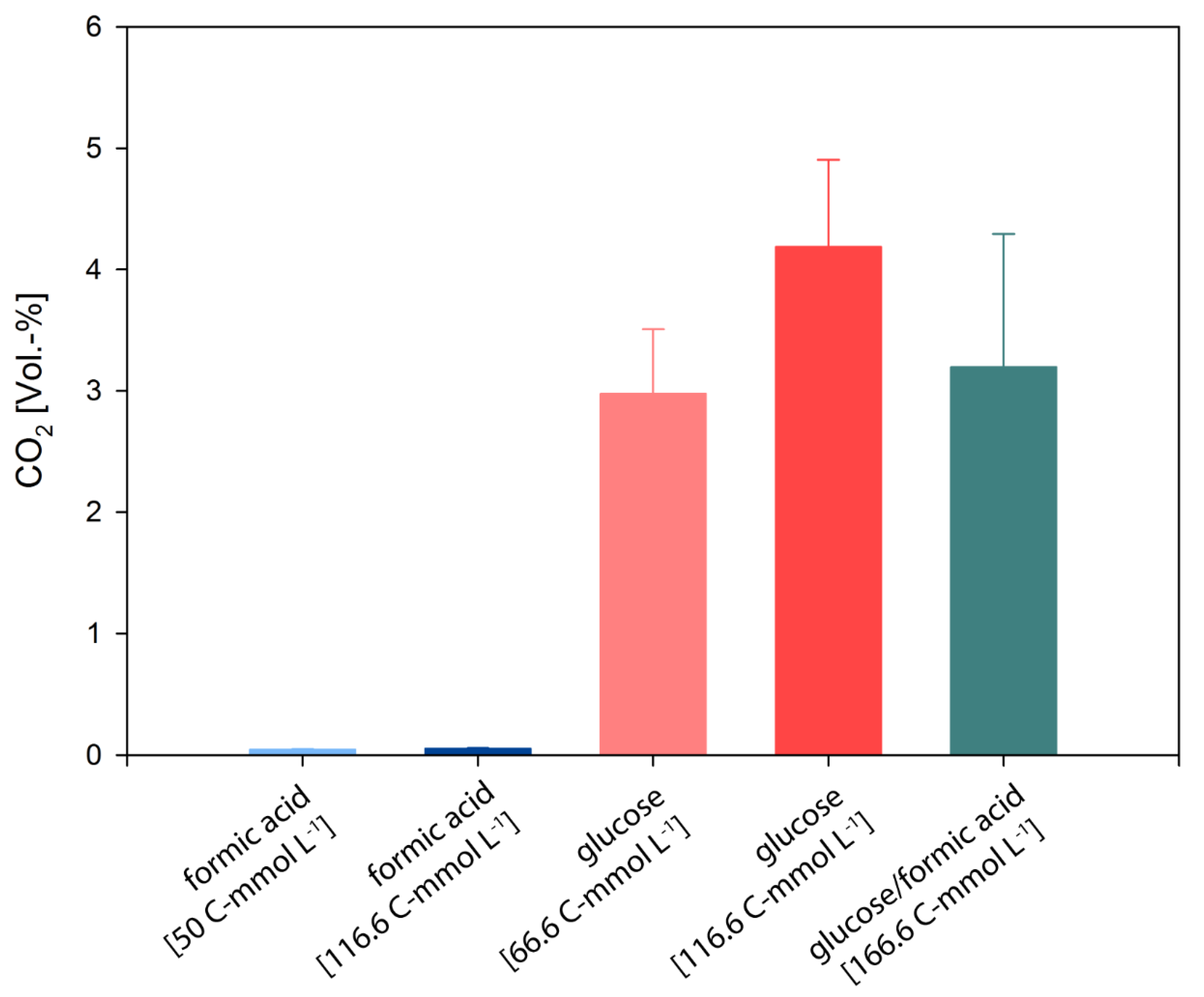
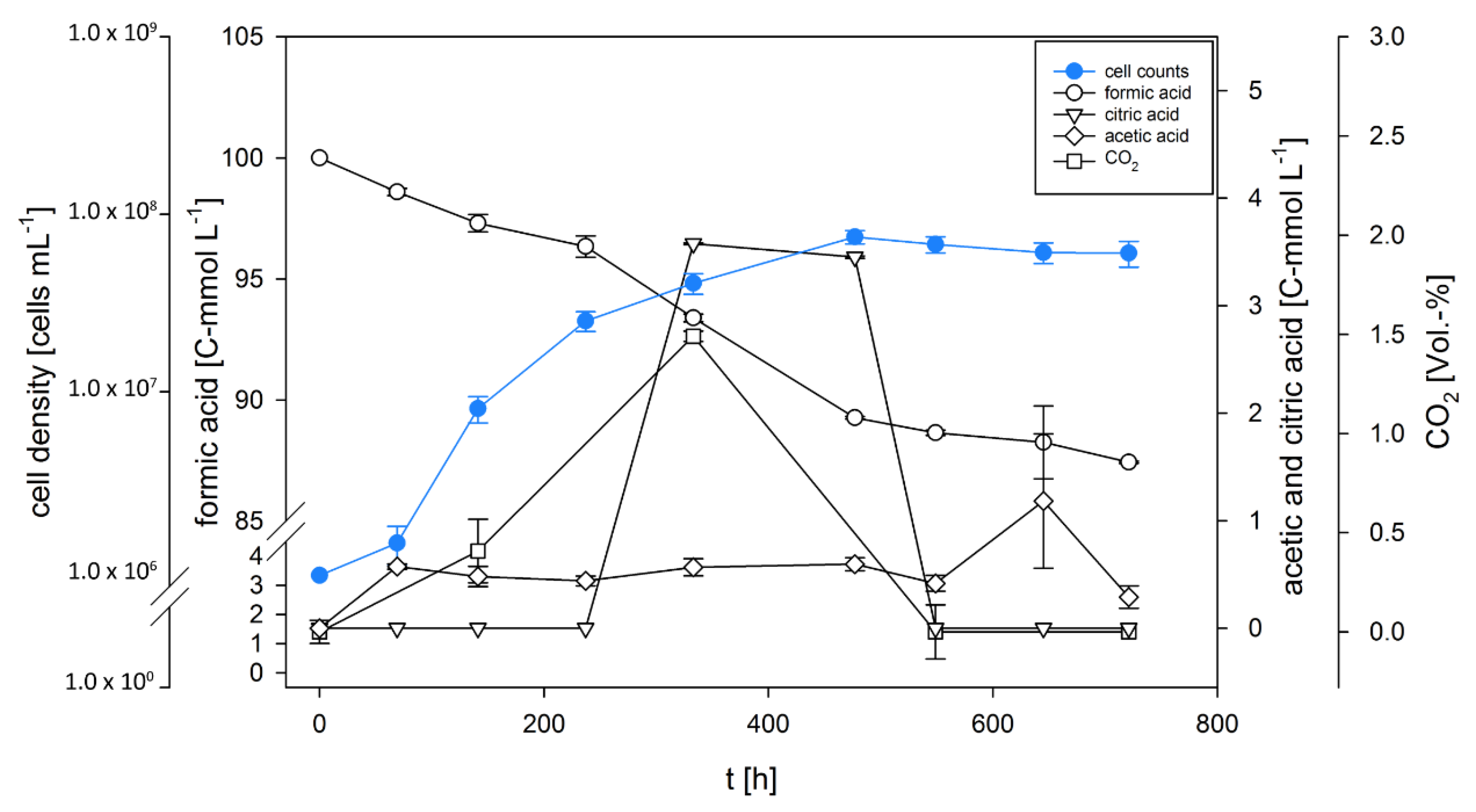
3.2. D. amylolyticus Converts Formate to CO2, Acetate, Citrate, and Ethanol
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gibson, H.W. Chemistry of formic acid and its simple derivatives. Chem. Rev. 1969, 69, 673–692. [Google Scholar] [CrossRef]
- Guillory, J.K. Dissociation Constants of Organic Acids and Bases. J. Med. Chem. 2009, 52, 5560. [Google Scholar] [CrossRef]
- Agarwal, A.S.; Zhai, Y.; Hill, D.; Sridhar, N. The Electrochemical Reduction of Carbon Dioxide to Formate/Formic Acid: Engineering and Economic Feasibility. ChemSusChem 2011, 4, 1301–1310. [Google Scholar] [CrossRef]
- Kopljar, D.; Inan, A.; Vindayer, P.; Wagner, N.; Klemm, E. Electrochemical reduction of CO2 to formate at high current density using gas diffusion electrodes. J. Appl. Electrochem. 2014, 44, 1107–1116. [Google Scholar] [CrossRef]
- Wang, Q.; Dong, H.; Yu, H. Fabrication of a novel tin gas diffusion electrode for electrochemical reduction of carbon dioxide to formic acid. RSC Adv. 2014, 4, 59970–59976. [Google Scholar] [CrossRef]
- Lu, X.; Leung, D.Y.C.; Wang, H.; Leung, M.K.H.; Xuan, J. Electrochemical Reduction of Carbon Dioxide to Formic Acid. ChemElectroChem 2014, 1, 836–849. [Google Scholar] [CrossRef]
- Taheri, A.; Berben, L.A. Making C–H bonds with CO2: Production of formate by molecular electrocatalysts. ChemComm 2016, 52, 1768–1777. [Google Scholar] [CrossRef]
- Albert, J.; Wölfel, R.; Bösmann, A.; Wasserscheid, P. Selective oxidation of complex, water-insoluble biomass to formic acid using additives as reaction accelerators. Energy Environ. Sci. 2012, 5, 7956–7962. [Google Scholar] [CrossRef]
- Sorokin, A.B.; Kudrik, E.V.; Alvarez, L.X.; Afanasiev, P.; Millet, J.M.M.; Bouchu, D. Oxidation of methane and ethylene in water at ambient conditions. Catal. Today 2010, 157, 149–154. [Google Scholar] [CrossRef]
- Shukla, R.S.; Bhatt, S.D.; Thorat, R.B.; Jasra, R.V. A novel effective hydration of carbon monoxide in liquid phase by a water-soluble ruthenium complex catalyst at moderate pressures in aqueous medium. Appl. Catal. A Gen. 2005, 294, 111–118. [Google Scholar] [CrossRef]
- Li, H.; Opgenorth, P.H.; Wernick, D.G.; Rogers, S.; Wu, T.-Y.; Higashide, W.; Malati, P.; Huo, Y.-X.; Cho, K.M.; Liao, J.C. Integrated Electromicrobial Conversion of CO2 to Higher Alcohols. Science 2012, 335, 1596. [Google Scholar] [CrossRef]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Christensen, K.E.; MacKenzie, R.E. Chapter 14 Mitochondrial Methylenetetrahydrofolate Dehydrogenase, Methenyltetrahydrofolate Cyclohydrolase, and Formyltetrahydrofolate Synthetases. In Vitam Horm; Elsevier: Amsterdam, The Netherlands, 2008; Volume 79, pp. 393–410. [Google Scholar]
- Stairs, C.W.; Roger, A.J.; Hampl, V. Eukaryotic Pyruvate Formate Lyase and Its Activating Enzyme Were Acquired Laterally from a Firmicute. Mol. Biol. Evol. 2011, 28, 2087–2099. [Google Scholar] [CrossRef] [Green Version]
- Wray, J. Extract of a Letter, Written by Mr. John Wray to the Publisher Concerning some uncommon observations and experiments made with an acid juice to be found in ants. Philos. Trans. 1670, 2063–2066. [Google Scholar]
- LeBrun, E.G.; Jones, N.T.; Gilbert, L.E. Chemical Warfare Among Invaders: A Detoxification Interaction Facilitates an Ant Invasion. Science 2014, 343, 1014–1017. [Google Scholar] [CrossRef]
- Thompson, J.L.; Hinton, M. Antibacterial activity of formic and propionic acids in the diet of hens on salmonellas in the crop. Br. Poult. Sci. 1997, 38, 59–65. [Google Scholar] [CrossRef]
- Hietala, J.; Vuori, A.; Johnsson, P.; Pollari, I.; Reutemann, W.; Kieczka, H. Formic Acid. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2016; pp. 1–22. [Google Scholar]
- Zhou, W.; Lee, J.Y. Particle Size Effects in Pd-Catalyzed Electrooxidation of Formic Acid. J. Phys. Chem. 2008, 112, 3789–3793. [Google Scholar] [CrossRef]
- Enthaler, S.; von Langermann, J.; Schmidt, T. Carbon dioxide and formic acid—The couple for environmental-friendly hydrogen storage? Energy Environ. Sci. 2010, 3, 1207. [Google Scholar] [CrossRef]
- Grasemann, M.; Laurenczy, G. Formic acid as a hydrogen source—Recent developments and future trends. Energy Environ. Sci. 2012, 5, 8171. [Google Scholar] [CrossRef]
- Yishai, O.; Lindner, S.N.; Gonzalez de la Cruz, J.; Tenenboim, H.; Bar-Even, A. The formate bio-economy. Curr. Opin. Chem. Biol. 2016, 35, 1–9. [Google Scholar] [CrossRef]
- Drake, H.L.; Küsel, K.; Matthies, C. Acetogenic Prokaryotes. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 3–60. [Google Scholar]
- Thauer, R.K.; Kaster, A.-K.; Seedorf, H.; Buckel, W.; Hedderich, R. Methanogenic archaea: Ecologically relevant differences in energy conservation. Nat. Rev. Microbiol. 2008, 6, 579–591. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, H.S.; Kim, E.S.; Bae, S.S.; Lim, J.K.; Matsumi, R.; Lebedinsky, A.V.; Sokolova, T.G.; Kozhevnikova, D.A.; Cha, S.-S.; et al. Formate-driven growth coupled with H2 production. Nature 2010, 467, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Dolfing, J.; Jiang, B.; Henstra, A.M.; Stams, A.J.M.; Plugge, C.M. Syntrophic Growth on Formate: A New Microbial Niche in Anoxic Environments. Appl. Environ. Microbiol. 2008, 74, 6126–6131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Schuchmann, K.; Müller, V. Direct and Reversible Hydrogenation of CO2 to Formate by a Bacterial Carbon Dioxide Reductase. Science 2013, 342, 1382–1385. [Google Scholar] [CrossRef]
- Poehlein, A.; Schmidt, S.; Kaster, A.-K.; Goenrich, M.; Vollmers, J.; Thürmer, A.; Bertsch, J.; Schuchmann, K.; Voigt, B.; Hecker, M.; et al. An Ancient Pathway Combining Carbon Dioxide Fixation with the Generation and Utilization of a Sodium Ion Gradient for ATP Synthesis. PLoS ONE 2012, 7, e33439. [Google Scholar] [CrossRef] [Green Version]
- Crain, A.V.; Broderick, J.B. Pyruvate Formate-lyase and Its Activation by Pyruvate Formate-lyase Activating Enzyme. J. Biol. Chem. 2014, 289, 5723–5729. [Google Scholar] [CrossRef] [Green Version]
- Broderick, J.B.; Duffus, B.R.; Duschene, K.S.; Shepard, E.M. Radical S-Adenosylmethionine Enzymes. Chem. Rev. 2014, 114, 4229–4317. [Google Scholar] [CrossRef]
- Zelcbuch, L.; Lindner, S.N.; Zegman, Y.; Vainberg Slutskin, I.; Antonovsky, N.; Gleizer, S.; Milo, R.; Bar-Even, A. Pyruvate Formate-Lyase Enables Efficient Growth of Escherichia coli on Acetate and Formate. Biochemistry 2016, 55, 2423–2426. [Google Scholar] [CrossRef]
- Klibanov, A.M.; Alberti, B.N.; Zale, S.E. Enzymatic synthesis of formic acid from H2 and CO2 and production of hydrogen from formic acid. Biotechnol. Bioeng. 1982, 24, 25–36. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, C.; Zhao, H.; Xing, X.-H. Improvement of Hydrogen Productivity by Introduction of NADH Regeneration Pathway in Clostridium paraputrificum. Appl. Biochem. Biotechnol. 2012, 167, 732–742. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Nishimura, Y. Hydrogen production by fermentation using acetic acid and lactic acid. J. Biosci. Bioeng. 2007, 103, 236–241. [Google Scholar] [CrossRef]
- Bakonyi, P.; Nemestóthy, N.; Lövitusz, É.; Bélafi-Bakó, K. Application of Plackett–Burman experimental design to optimize biohydrogen fermentation by E. coli (XL1-BLUE). Int. J. Hydrogen Energy 2011, 36, 13949–13954. [Google Scholar] [CrossRef]
- Hill, S.; Viollet, S.; Smith, A.T.; Anthony, C. Roles for enteric Formate Utilization by the Crenarchaeo d-type cytochrome oxidase in N2 fixation and microaerobiosis. J. Bacteriol. 1990, 172, 2071–2078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, T.; Sanchez-Torres, V.; Wood, T.K. Metabolic engineering to enhance bacterial hydrogen production. Microb. Biotechnol. 2008, 1, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Pakes, W.C.C.; Jollyman, W.H. XL.—The bacterial decomposition of formic acid into carbon dioxide and hydrogen. J. Chem. Soc. Trans. 1901, 79, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Seol, E.; Kim, S.; Raj, S.M.; Park, S. Comparison of hydrogen-production capability of four different Enterobacteriaceae strains under growing and non-growing conditions. Int. J. Hydrogen Energy 2008, 33, 5169–5175. [Google Scholar] [CrossRef]
- Seol, E.; Manimaran, A.; Jang, Y.; Kim, S.; Oh, Y.-K.; Park, S. Sustained hydrogen production from formate using immobilized recombinant Escherichia coli SH5. Int. J. Hydrogen Energy 2011, 36, 8681–8686. [Google Scholar] [CrossRef]
- Shin, J.-H.; Yoon, J.H.; Lee, S.H.; Park, T.H. Hydrogen production from formic acid in pH-stat fed-batch operation for direct supply to fuel cell. Bioresour. Technol. 2010, 101, S53–S58. [Google Scholar] [CrossRef]
- Yoshida, A.; Nishimura, T.; Kawaguchi, H.; Inui, M.; Yukawa, H. Enhanced Hydrogen Production from Formic Acid by Formate Hydrogen Lyase-Overexpressing Escherichia coli Strains. Appl. Environ. Microbiol. 2005, 71, 6762–6768. [Google Scholar] [CrossRef]
- Yoshida, A.; Nishimura, T.; Kawaguchi, H.; Inui, M.; Yukawa, H. Efficient induction of formate hydrogen lyase of aerobically grown Escherichia coli in a three-step biohydrogen production process. Appl. Microbiol. Biotechnol. 2007, 74, 754–760. [Google Scholar] [CrossRef]
- Bagramyan, K.; Mnatsakanyan, N.; Poladian, A.; Vassilian, A.; Trchounian, A. The roles of hydrogenases 3 and 4, and the F0F1-ATPase, in H2 production by Escherichia coli at alkaline and acidic pH. FEBS Lett. 2002, 516, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T.; Woods, D.R. Acetone-butanol fermentation revisited. Microbiol. Rev. 1986, 50, 484–524. [Google Scholar] [CrossRef] [Green Version]
- Herrero, A.A.; Gomez, R.F.; Snedecor, B.; Tolman, C.J.; Roberts, M.F. Growth inhibition of Clostridium thermocellum by carboxylic acids: A mechanism based on uncoupling by weak acids. Appl. Microbiol. Biotechnol. 1985, 22, 53–62. [Google Scholar] [CrossRef]
- Bagramyan, K.; Trchounian, A. Structural and functional features of formate hydrogen lyase, an enzyme of mixed-acid fermentation from Escherichia coli. Biochemistry 2003, 68, 1159–1170. [Google Scholar] [CrossRef]
- Lim, J.K.; Bae, S.S.; Kim, T.W.; Lee, J.-H.; Lee, H.S.; Kang, S.G. Thermodynamics of Formate-Oxidizing Metabolism and Implications for H2 Production. Appl. Environ. Microbiol. 2012, 78, 7393–7397. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Seol, E.; Kaur, G.; Oh, Y.-K.; Park, S. Hydrogen production from C1 compounds by a novel marine hyperthermophilic archaeon Thermococcus onnurineus NA1. Int. J. Hydrogen Energy 2012, 37, 11113–11121. [Google Scholar] [CrossRef]
- Jackson, B.E.; McInerney, M.J. Anaerobic microbial metabolism can proceed close to thermodynamic limits. Nature 2002, 415, 454–456. [Google Scholar] [CrossRef]
- Mayer, F.; Müller, V. Adaptations of anaerobic archaea to life under extreme energy limitation. FEMS Microbiol. Rev. 2014, 38, 449–472. [Google Scholar] [CrossRef] [Green Version]
- Schink, B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Mol. Biol. Rev. 1997, 61, 262–280. [Google Scholar] [CrossRef] [Green Version]
- Stams, A.J.M.; Plugge, C.M. Electron transfer in syntrophic communities of anaerobic bacteria and archaea. Nat. Rev. Microbiol. 2009, 7, 568–577. [Google Scholar] [CrossRef]
- Lee, H.S.; Kang, S.G.; Bae, S.S.; Lim, J.K.; Cho, Y.; Kim, Y.J.; Jeon, J.H.; Cha, S.-S.; Kwon, K.K.; Kim, H.-T.; et al. The Complete Genome Sequence of Thermococcus onnurineus NA1 Reveals a Mixed Heterotrophic and Carboxydotrophic Metabolism. J. Bacteriol. 2008, 190, 7491–7499. [Google Scholar] [CrossRef] [Green Version]
- Moon, Y.-J.; Kwon, J.; Yun, S.-H.; Lim, H.L.; Kim, M.-S.; Kang, S.G.; Lee, J.-H.; Choi, J.-S.; Kim, S.I.; Chung, Y.-H. Proteome Analyses of Hydrogen-producing Hyperthermophilic Archaeon Thermococcus onnurineus NA1 in Different One-carbon Substrate Culture Conditions. Mol. Cell. Proteomics 2012, 11, M111.015420. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.K.; Mayer, F.; Kang, S.G.; Muller, V. Energy conservation by oxidation of formate to carbon dioxide and hydrogen via a sodium ion current in a hyperthermophilic archaeon. Proc. Natl. Acad. Sci. USA 2014, 111, 11497–11502. [Google Scholar] [CrossRef] [Green Version]
- Bar-Even, A. Formate Assimilation: The Metabolic Architecture of Natural and Synthetic Pathways. Biochemistry 2016, 55, 3851–3863. [Google Scholar] [CrossRef]
- Heim, L.E.; Konnerth, H.; Prechtl, M.H.G. Future perspectives for formaldehyde: Pathways for reductive synthesis and energy storage. Green Chem. 2017, 19, 2347–2355. [Google Scholar] [CrossRef] [Green Version]
- Flamholz, A.; Noor, E.; Bar-Even, A.; Milo, R. eQuilibrator—The biochemical thermodynamics calculator. Nucleic Acids Res. 2012, 40, D770–D775. [Google Scholar] [CrossRef] [Green Version]
- Ogushi, S.; Ando, M.; Tsuru, D. Formaldehyde dehydrogenase from Pseudomonas putida: A zinc metalloenzyme. J. Biochem. 1984, 96, 1587–1591. [Google Scholar] [CrossRef]
- Yoshimoto, M.; Yamashita, T.; Kinoshita, S. Thermal stabilization of formaldehyde dehydrogenase by encapsulation in liposomes with nicotinamide adenine dinucleotide. Enzyme Microb. Technol. 2011, 49, 209–214. [Google Scholar] [CrossRef]
- Perevalova, A.A. Desulfurococcus fermentans sp. nov., a novel hyperthermophilic archaeon from a Kamchatka hot spring, and emended description of the genus Desulfurococcus. Int. J. Syst. Evol. Microbiol. 2005, 55, 995–999. [Google Scholar] [CrossRef]
- Reischl, B.; Ergal, İ.; Rittmann, S.K.-M.R. Biohydrogen production characteristics of Desulfurococcus amylolyticus DSM 16532. Int. J. Hydrogen Energy 2018, 43, 8747–8753. [Google Scholar] [CrossRef]
- Reischl, B.; Ergal, İ.; Rittmann, S.K.-M.R. Metabolic reconstruction and experimental verification of glucose utilization in Desulfurococcus amylolyticus DSM 16532. Folia Microbiol. 2018, 63, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Rittmann, S.K.-M.R.; Lee, H.S.; Lim, J.K.; Kim, T.W.; Lee, J.-H.; Kang, S.G. One-carbon substrate-based biohydrogen production: Microbes, mechanism, and productivity. Biotechnol. Adv. 2015, 33, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Ergal, İ.; Fuchs, W.; Hasibar, B.; Thallinger, B.; Bochmann, G.; Rittmann, S.K.-M.R. The physiology and biotechnology of dark fermentative biohydrogen production. Biotechnol. Adv. 2018, 36, 2165–2186. [Google Scholar] [CrossRef] [PubMed]
- Kublanov, I.V.; Bidzhieva, S.K.; Perevalova, A.A.; Lebedinsky, A.V.; Mukhopadhyay, B.; Bonch-Osmolovskaya, E.A. Reclassification of Desulfurococcus mobilis as a synonym of Desulfurococcus mucosus, Desulfurococcus fermentans and Desulfurococcus kamchatkensis as synonyms of Desulfurococcus amylolyticus, and emendation of the D. mucosus and D. amylolyticus species descriptions. Int. J. Syst. Evol. Microbiol. 2016, 66, 514–517. [Google Scholar]
- Erickson, L.E. Biomass elemental composition and energy content. Biotechnol. Bioeng. 1980, 22, 451–456. [Google Scholar] [CrossRef]
- Amend, J.P.; Shock, E.L. Energetics of overall metabolic reactions of thermophilic and hyperthermophilic Archaea and bacteria. FEMS Microbiol. Rev. 2001, 25, 175–243. [Google Scholar] [CrossRef]
- Schulte, M.D.; Shock, E.L. Aldehydes in hydrothermal solution: Standard partial molal thermodynamic properties and relative stabilities at high temperatures and pressures. Geochim. Cosmochim. Acta 1993, 57, 3835–3846. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Tatusova, T.; Brown, G.R.; Maglott, D.R. NCBI Reference Sequences (RefSeq): Current status, new features and genome annotation policy. Nucleic Acids Res. 2012, 40, D130–D135. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16. [Google Scholar] [CrossRef] [Green Version]
- Bateman, A. The Pfam protein families database. Nucleic Acids Res. 2004, 32, 138–141. [Google Scholar] [CrossRef]
- Monod, J. Recherches Sur la Croissance Des Cultures Bactériennes; Hermann: Paris, France, 1941. [Google Scholar]
- Gaudy, A.F.; Obayashi, A.; Gaudy, E.T. Control of growth rate by initial substrate concentration at values below maximum rate. Appl. Microbiol. 1971, 22, 1041–1047. [Google Scholar] [CrossRef] [Green Version]
- Robb, F.T.; Shukla, H.D.; Clark, D.S. 10 Heat Shock Proteins in Hyperthermophiles. In Methods in Microbiology; Elsevier: Amsterdam, The Netherlands, 2006; Volume 35, pp. 233–252. [Google Scholar]
- Schäfer, T.; Schönheit, P. Pyruvate metabolism of the hyperthermophilic archaebacterium Pyrococcus furiosus: Acetate formation from acetyl-CoA and ATP synthesis are catalyzed by an acetyl-CoA synthetase (ADP forming). Arch. Microbiol. 1991, 155, 366–377. [Google Scholar] [CrossRef]
- Hansen, T.; Schönheit, P. Purification and properties of the first-identified, archaeal, ATP-dependent 6-phosphofructokinase, an extremely thermophilic non-allosteric enzyme, from the hyperthermophile Desulfurococcus amylolyticus. Arch. Microbiol. 2000, 173, 103–109. [Google Scholar] [CrossRef]
- Lin, B.; Tao, Y. Whole-cell biocatalysts by design. Microb. Cell Fact. 2017, 16, 106. [Google Scholar] [CrossRef] [Green Version]
- Fukui, T. Complete genome sequence of the hyperthermophilic archaeon Thermococcus kodakaraensis KOD1 and comparison with Pyrococcus genomes. Genome Res. 2005, 15, 352–363. [Google Scholar] [CrossRef] [Green Version]
- Mardanov, A.V.; Ravin, N.V.; Svetlitchnyi, V.A.; Beletsky, A.V.; Miroshnichenko, M.L.; Bonch-Osmolovskaya, E.A.; Skryabin, K.G. Metabolic Versatility and Indigenous Origin of the Archaeon Thermococcus sibiricus, Isolated from a Siberian Oil Reservoir, as Revealed by Genome Analysis. Appl. Environ. Microbiol. 2009, 75, 4580–4588. [Google Scholar] [CrossRef] [Green Version]
- Rossmann, R.; Sawers, G.; Böck, A. Mechanism of regulation of the formate-hydrogenlyase pathway by oxygen, nitrate, and pH: Definition of the formate regulon. Mol. Microbiol. 1991, 5, 2807–2814. [Google Scholar] [CrossRef]
- Goldberg, I.; Rock, J.S.; Ben-Bassat, A.; Mateles, R.I. Bacterial yields on methanol, methylamine, formaldehyde, and formate. Biotechnol. Bioeng. 1976, 18, 1657–1668. [Google Scholar] [CrossRef]
- Bar-Even, A.; Noor, E.; Flamholz, A.; Milo, R. Design and analysis of metabolic pathways supporting formatotrophic growth for electricity-dependent cultivation of microbes. Biochim. Biophys. Acta Bioenerg. 2013, 1827, 1039–1047. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Edlich-Muth, C.; Lindner, S.N.; Bar-Even, A. Ribulose Monophosphate Shunt Provides Nearly All Biomass and Energy Required for Growth of E. coli. ACS Synth. Biol. 2018, 7, 1601–1611. [Google Scholar] [CrossRef]
- Pickl, A.; Schönheit, P. The oxidative pentose phosphate pathway in the haloarchaeon Haloferax volcanii involves a novel type of glucose-6-phosphate dehydrogenase—The archaeal Zwischenferment. FEBS Lett. 2015, 589, 1105–1111. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Imanaka, H.; Rashid, N.; Fukui, T.; Atomi, H.; Imanaka, T. Genetic Evidence Identifying the True Gluconeogenic Fructose-1,6-Bisphosphatase in Thermococcus kodakaraensis and Other Hyperthermophiles. J. Bacteriol. 2004, 186, 5799–5807. [Google Scholar] [CrossRef] [Green Version]
- Orita, I.; Yurimoto, H.; Hirai, R.; Kawarabayasi, Y.; Sakai, Y.; Kato, N. The Archaeon Pyrococcus horikoshii Possesses a Bifunctional Enzyme for Formaldehyde Fixation via the Ribulose Monophosphate Pathway. J. Bacteriol. 2005, 187, 3636–3642. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Inui, M.; Yukawa, H.; Kanao, T.; Tomizawa, K.; Atomi, H.; Imanaka, T. Phototrophic growth of a Rubisco-deficient mesophilic purple nonsulfur bacterium harboring a Type III Rubisco from a hyperthermophilic archaeon. J. Biotechnol. 2006, 124, 532–544. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
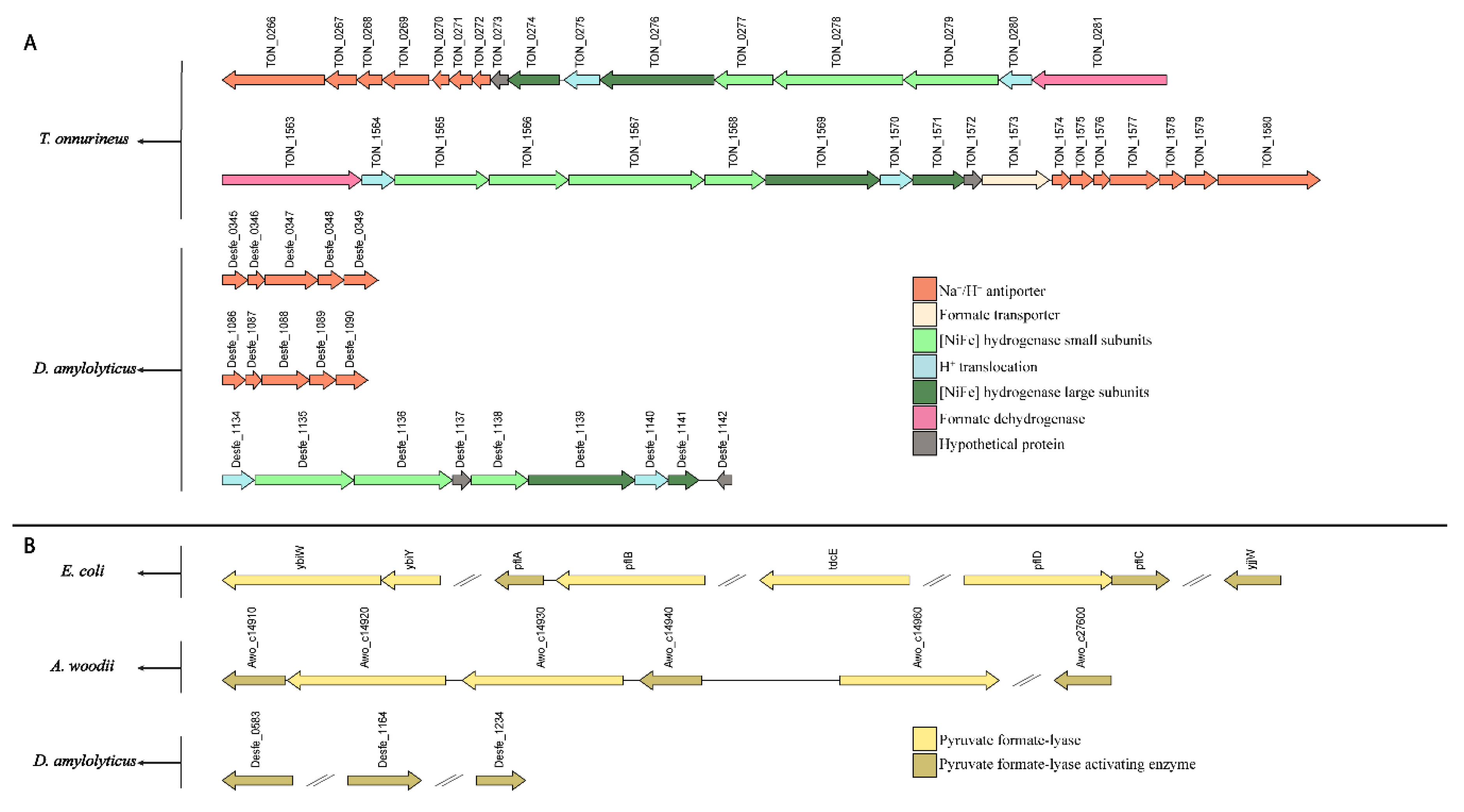{kind=link}
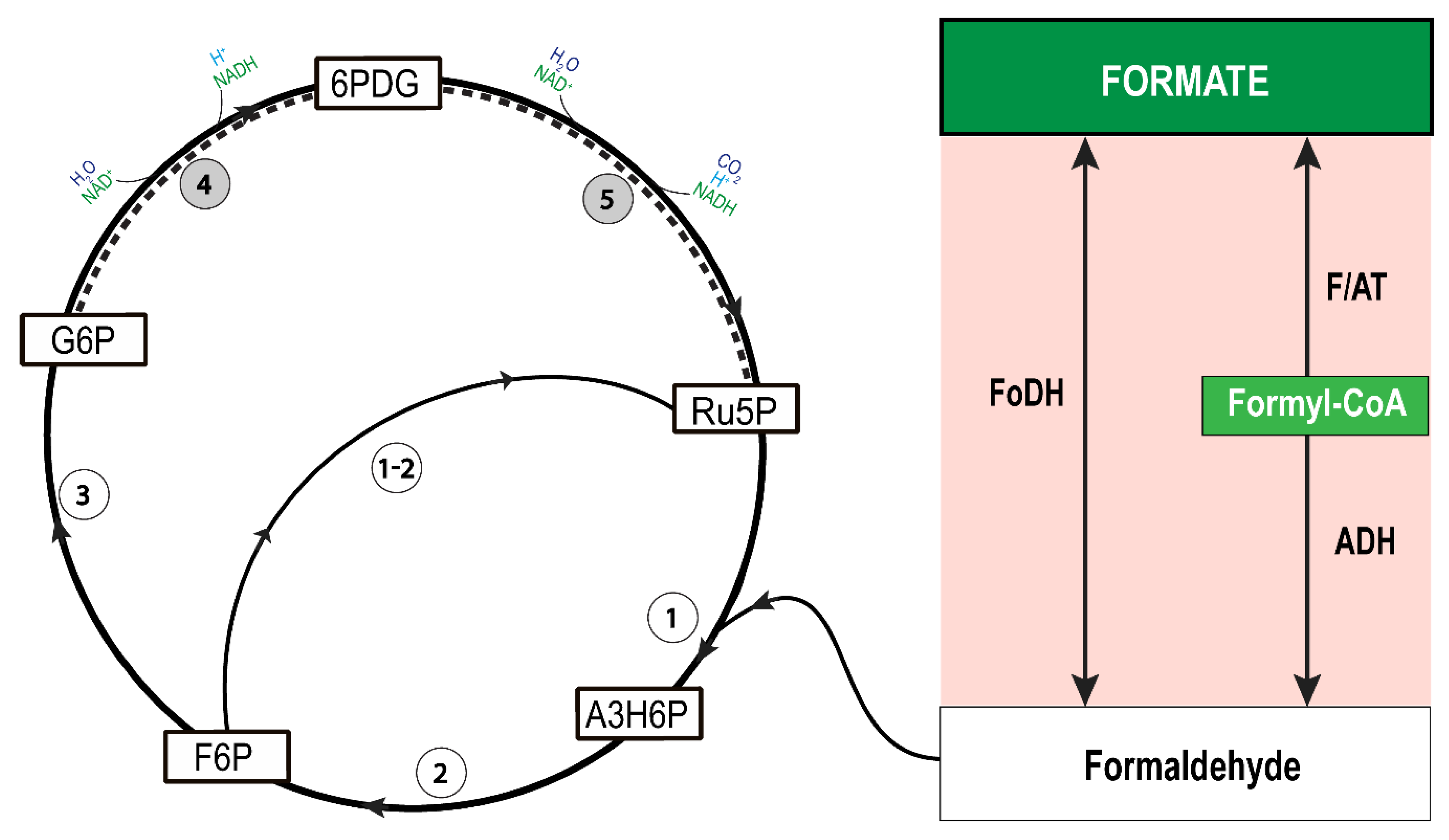{kind=link}
| Compound and Concentration | μmax [h−1] | μmean [h−1] | Final Cell Concentration [cells per mL] |
|---|---|---|---|
| Glucose [66.6 C-mmol L−1] | 0.033 | 0.011 | 1.55·107 |
| Glucose [116.6 C-mmol L−1] | 0.035 | 0.012 | 2.09·107 |
| Formic acid [50 C-mmol L−1] | 0.032 | 0.010 | 1.24·107 |
| Formic acid [116.6 C-mmol L−1] | 0.032 | 0.011 | 2.08·107 |
| Glucose/Formic acid [66 and 50 C-mmol L−1] | 0.036 | 0.012 | 2.38·107 |
| Compound and Concentration | CER [mmol L−1 h−1] (C-Molar) | qCO2 [pmol h−1 g−1] (C-Molar) | Y(CO2/s) [C-mol/C-mol] * | Y(Lact/s) [C-mol/C-mol] * | Y(Ac/s) [C-mol/C-mol] * | Y(Form/s) [C-mol/C-mol] * | Y(Glu/s) [C-mol/ C-mol] * | Y(Buty/s) [C-mol/C-mol] * | Y(Citr/s) [C-mol/C-mol] * | Y(Eth/s) [C-mol/C-mol] * | Y(x/s) [C-mol/C-mol] | C-Balance + | DoR-Balance # |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Glucose [66.6 C-mmol L−1] | 3.34·10−5 | 2.41·10−3 | 2.64·10−2 | 2.96·10−1 | 5.64·10−1 | 6.14·10−2 | 1.29·10−4 | 94.80% | 89.08% | ||||
| Glucose [116.6 C-mmol L−1] | 4.04·10−5 | 2.59·10−3 | 4.43·10−1 | 2.48·10−1 | 5.89·10−1 | 8.10·10−2 | 2.42·10−3 | 136.12% | 87.82% | ||||
| Formic acid [50 C-mmol L−1] | 1.20·10−6 | 1.16·10−2 | 7.20·10−4 | 1.45·10−1 | 8.69·10−3 | 2.15·10−4 | 5.59·10−2 | 5.17·10−1 | 6.34·10−5 | 72.79% | 194.37% | ||
| Formic acid [116.6 C-mmol L−1] | 3.48·10−6 | 2.23·10−4 | 5.02·10−4 | 6.21·10−1 | 5.12·10−2 | 1.02·10−2 | 1.59·10−2 | 3.18·10−5 | 69.84% | 140.65% |
| Time [h] | CER [mmol L−1 h−1] (C-Molar) | qCO2 [pmol h−1 g−1] (C-Molar) | Y(CO2/s) [C-mol/C-mol] * | Y(Ac/s) [C-mol/C-mol] * | Y(Citr/s) [C-mol/C-mol] * | Y(x/s) [C-mol/C-mol] * | C-Balance + | DoR-Balance # |
|---|---|---|---|---|---|---|---|---|
| 69 | 4.09·10−1 | 1.05·10−5 | 40.93% | 81.87% | ||||
| 141 | 2.12·10−5 | 2.65·10−6 | 2.32·10−3 | 3.73·10−1 | 6.57·10−5 | 37.55% | 74.67% | |
| 237 | 4.57·10−1 | 2.75·10−4 | 45.75% | 91.53% | ||||
| 333 | 5.35·10−5 | 1.31·10−6 | 6.04·10−3 | 1.93·10−1 | 1.21·100 | 1.46·10−4 | 141.12% | 220.45% |
| 477 | 1.44·10−1 | 8.36·10−1 | 1.91·10−4 | 98.05% | 154.30% | |||
| 549 | NA § | NA§ | 1.82·10−3 | 6.74·10−1 | 1.15·10−3 | 66.53% | 134.93% | |
| 645 | 3.03·100 | 1.65·10−3 | 302.83% | 90.15% | ||||
| 721 | NA § | NA§ | 3.53·10−3 | 3.54·10−1 | 7.76·10−4 | 34.11% | 70.77% |
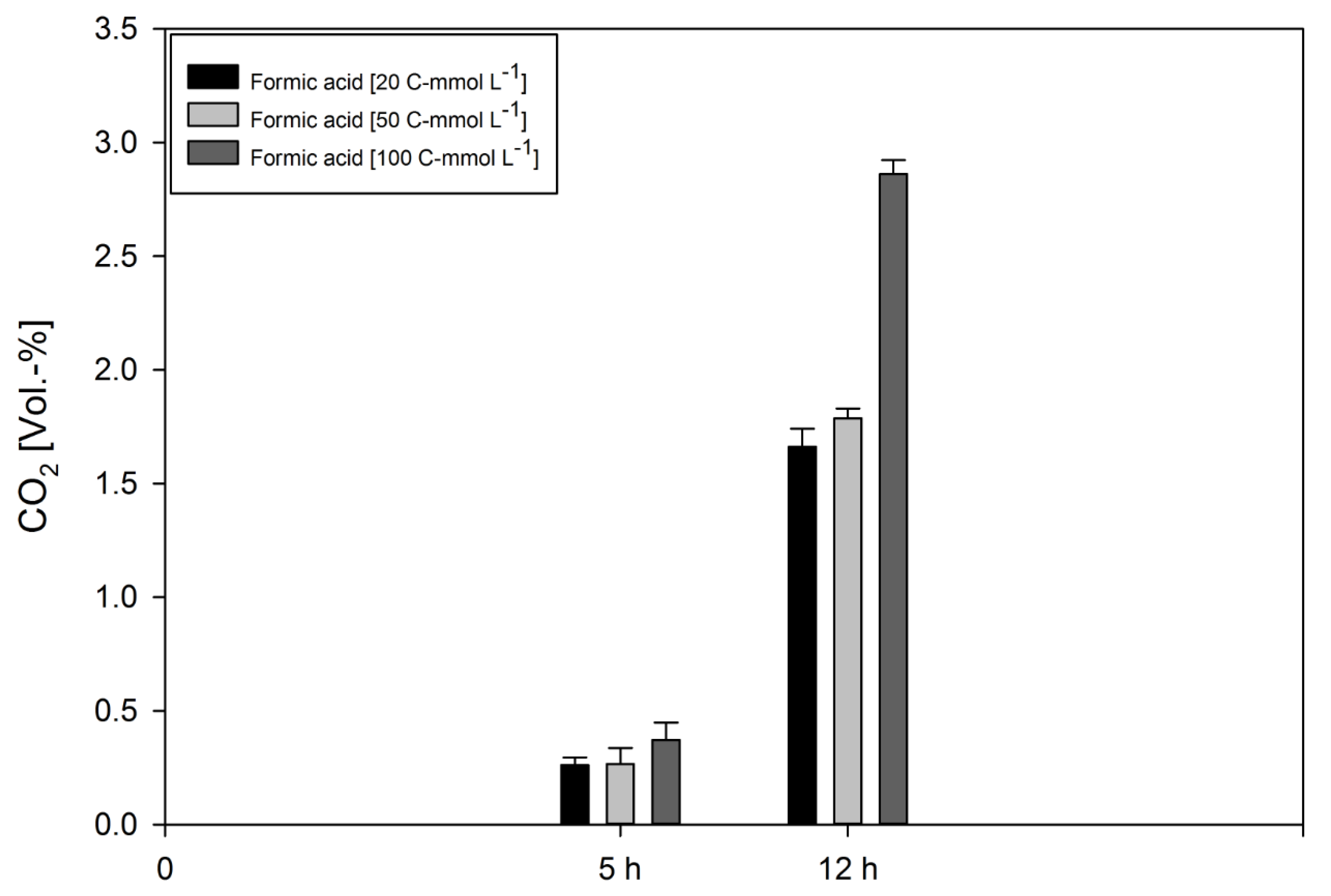
| Time [h] | Concentration [C-mmol L−1] | Formic acid Consumption [%] | CER [mmol L−1 h−1] (C-Molar) | qCO2 [mmol h−1 g−1] (C-Molar) | Y(CO2/s) * [C-mol/C-mol] | Y(Ac/s) * [C-mol/C-mol] | Y(Buty/s) * [C-mol/C-mol] | Y(Citr/s) * [C-mol/C-mol] | Y(Eth/s) * [C-mol/C-mol] |
|---|---|---|---|---|---|---|---|---|---|
| 5 | 100 | 1.42 | 2.50·10−5 | 1.60·10−15 | 8.78·10−5 | 3.41·10−3 | 5.72·10−2 | ||
| 5 | 50 | 0.71 | 1.79·10−5 | 1.14·10−15 | 2.53·10−4 | 1.97·10−3 | 2.95·10−2 | ||
| 5 | 20 | 0.08 | 1.76·10−5 | 1.13·10−15 | 5.41·10−3 | ||||
| 12 | 100 | 41.05 | 1.93·10−4 | 1.23·10−14 | 5.63·10−5 | 3.06·10−1 | 1.43·10−4 | 4.63·10−4 | |
| 12 | 50 | 9.35 | 1.20·10−4 | 7.71·10−15 | 3.09·10−4 | 7.72·10−3 | 2.81·10−2 | ||
| 12 | 20 | 9.36 | 1.12·10−4 | 7.17·10−15 | 7.18·10−5 | 1.92·10−4 | 1.26·10−2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ergal, I.; Reischl, B.; Hasibar, B.; Manoharan, L.; Zipperle, A.; Bochmann, G.; Fuchs, W.; Rittmann, S.K.-M.R. Formate Utilization by the Crenarchaeon Desulfurococcus amylolyticus. Microorganisms 2020, 8, 454. https://doi.org/10.3390/microorganisms8030454
Ergal I, Reischl B, Hasibar B, Manoharan L, Zipperle A, Bochmann G, Fuchs W, Rittmann SK-MR. Formate Utilization by the Crenarchaeon Desulfurococcus amylolyticus. Microorganisms. 2020; 8(3):454. https://doi.org/10.3390/microorganisms8030454
Chicago/Turabian StyleErgal, Ipek, Barbara Reischl, Benedikt Hasibar, Lokeshwaran Manoharan, Aaron Zipperle, Günther Bochmann, Werner Fuchs, and Simon K.-M. R. Rittmann. 2020. "Formate Utilization by the Crenarchaeon Desulfurococcus amylolyticus" Microorganisms 8, no. 3: 454. https://doi.org/10.3390/microorganisms8030454