Inactivation of the sfgtr4 Gene of Shigella flexneri Induces Biofilm Formation and Affects Bacterial Pathogenicity


Abstract
:1. Introduction
2. Material and Methods
2.1. Bacterial Strains and Growth Conditions
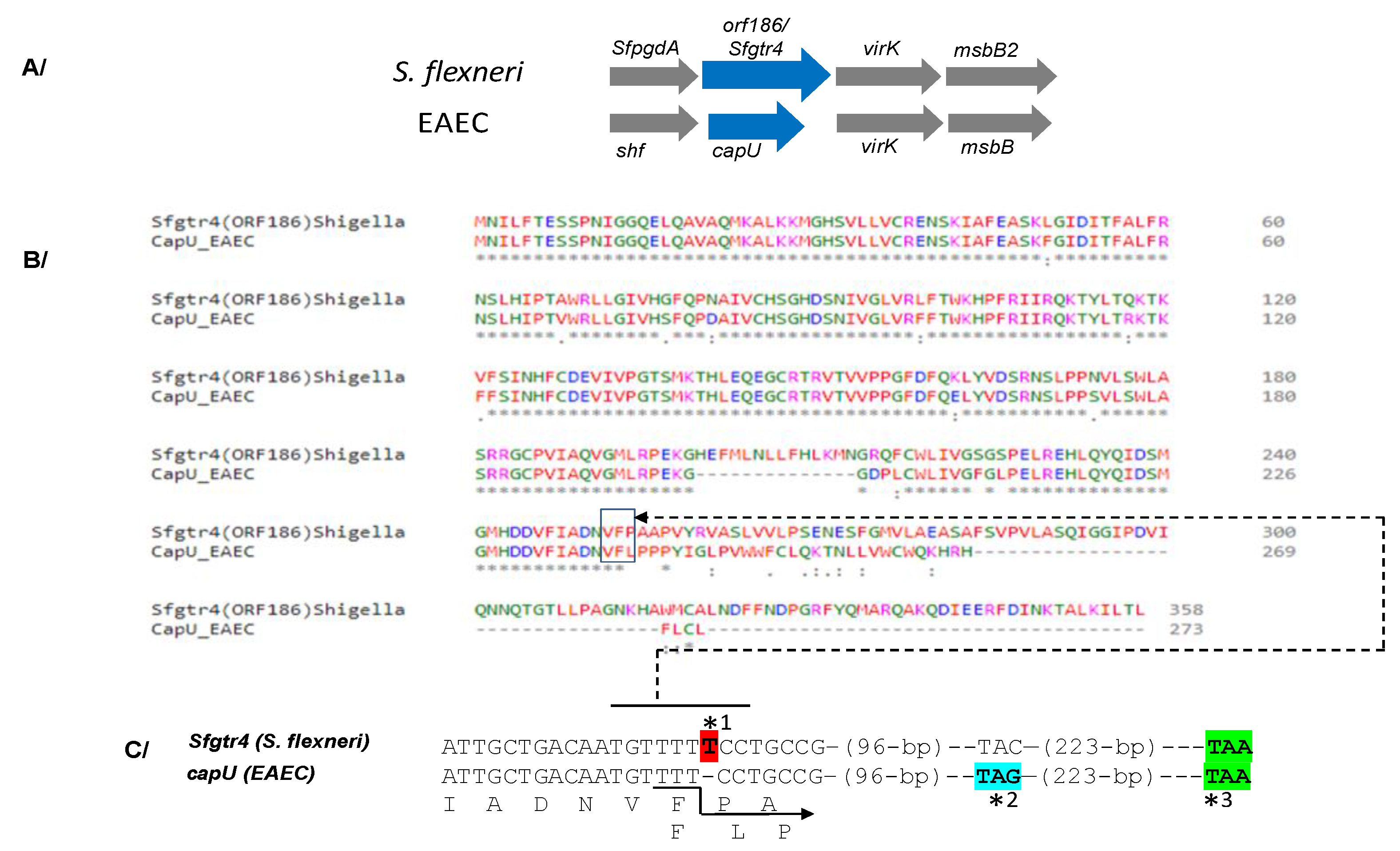
2.2. Construction of Single sfgtr4 and Double sfpgdA-sfgtr4 Mutants
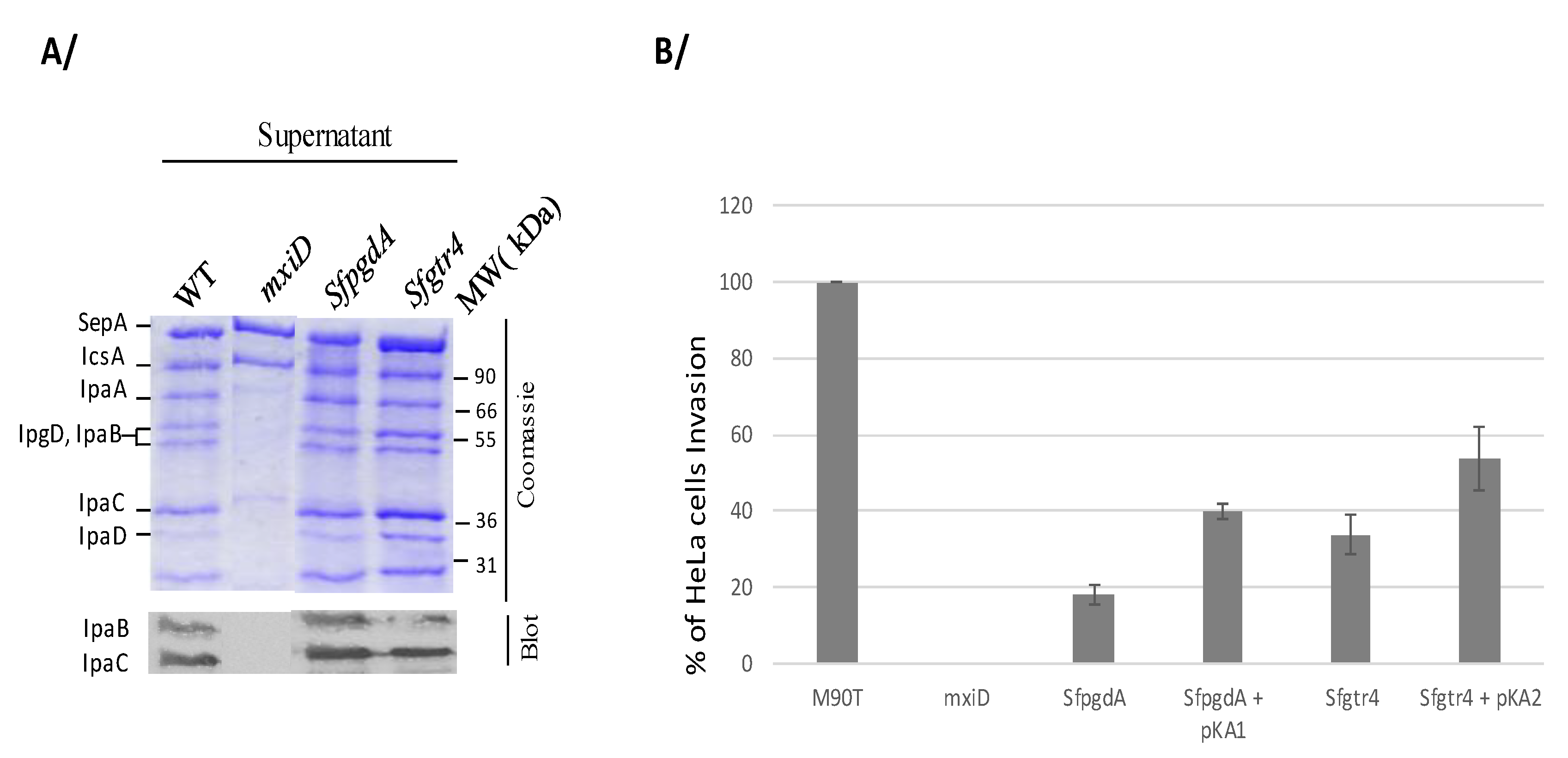
2.3. Proteins Secretion and HeLa Cells Invasion
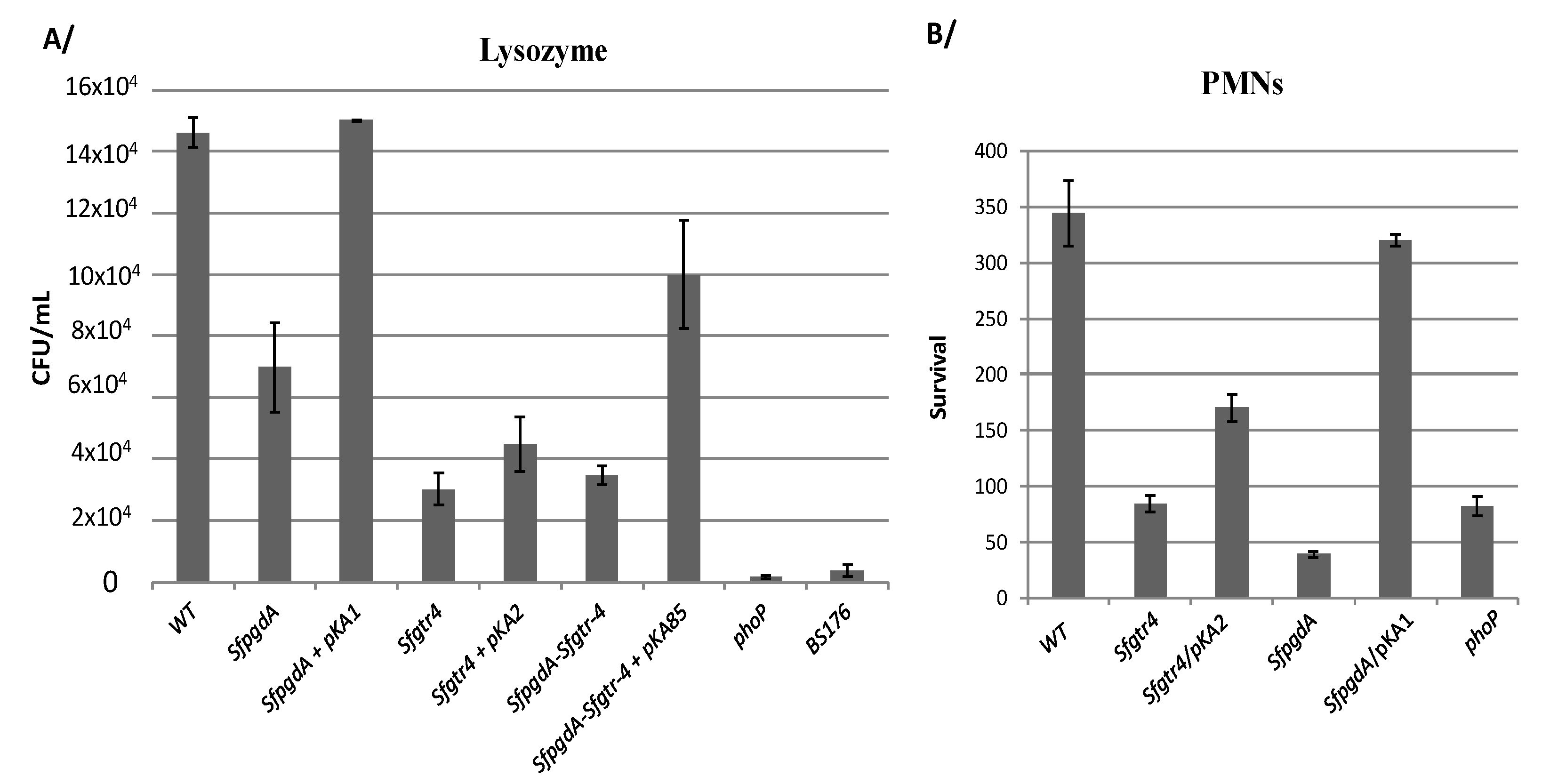
2.4. Lysozyme Sensitivity Assay and Polymorphonuclear Neutrophils (PMNs) Bacterial Infection
2.5. Measuring the Biofilm Formation by Shigella
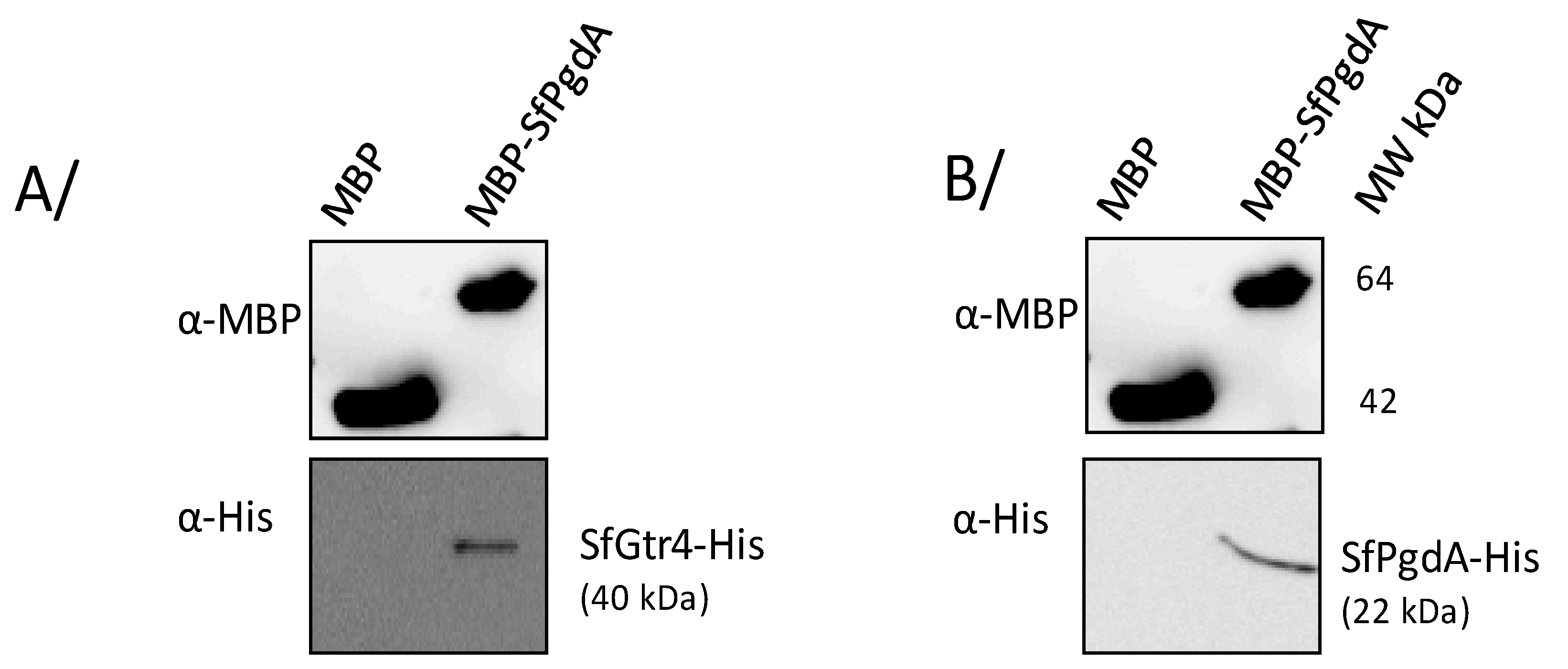
2.6. Protein Purification and Maltose Binding Protein (MBP) Pull-Down Assay
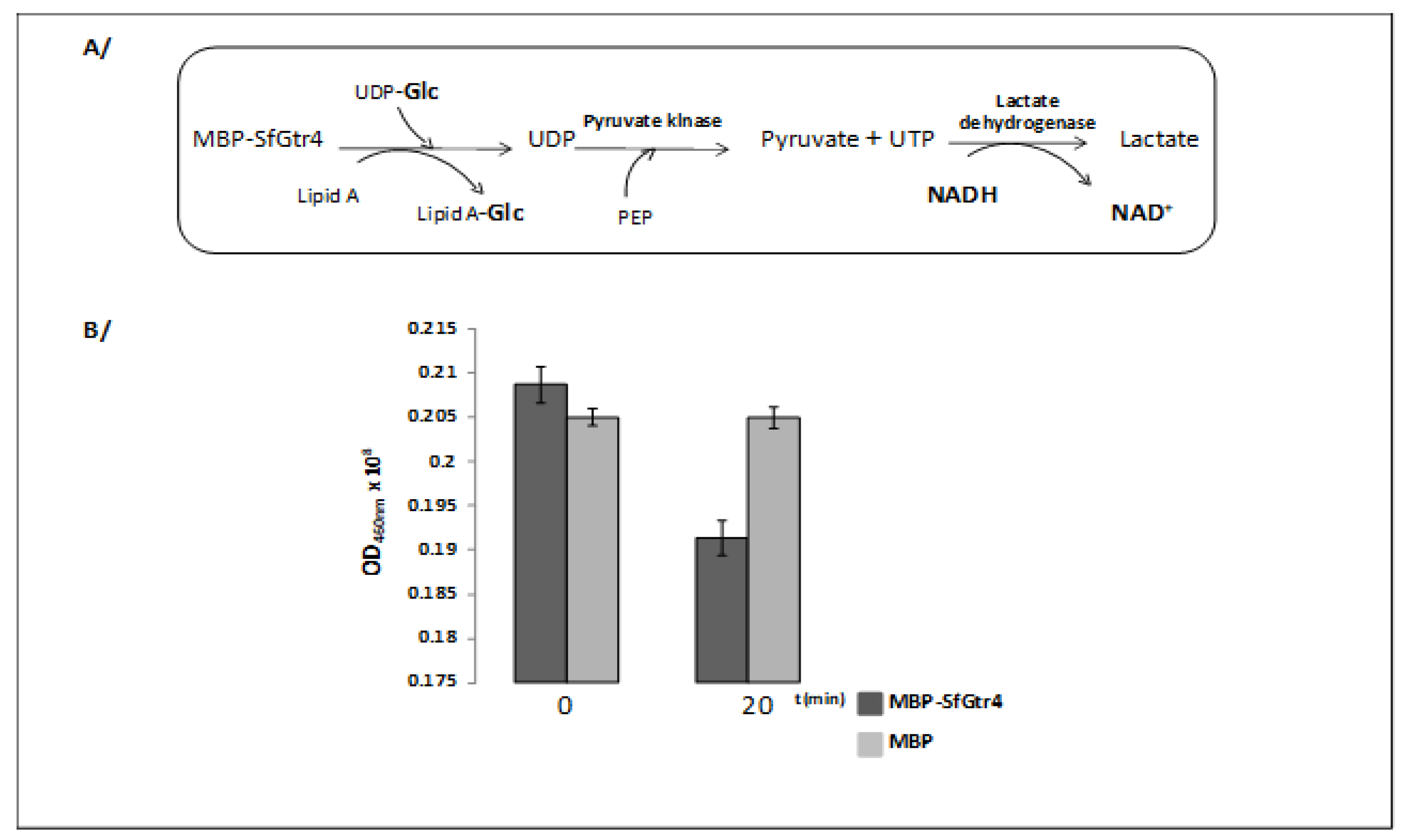
2.7. Detection of the Glycosyltransferase Enzymatic Activity
3. Results
3.1. Purified SfGtr4 Exhibits a Glycosyltransferase Enzymatic Activity
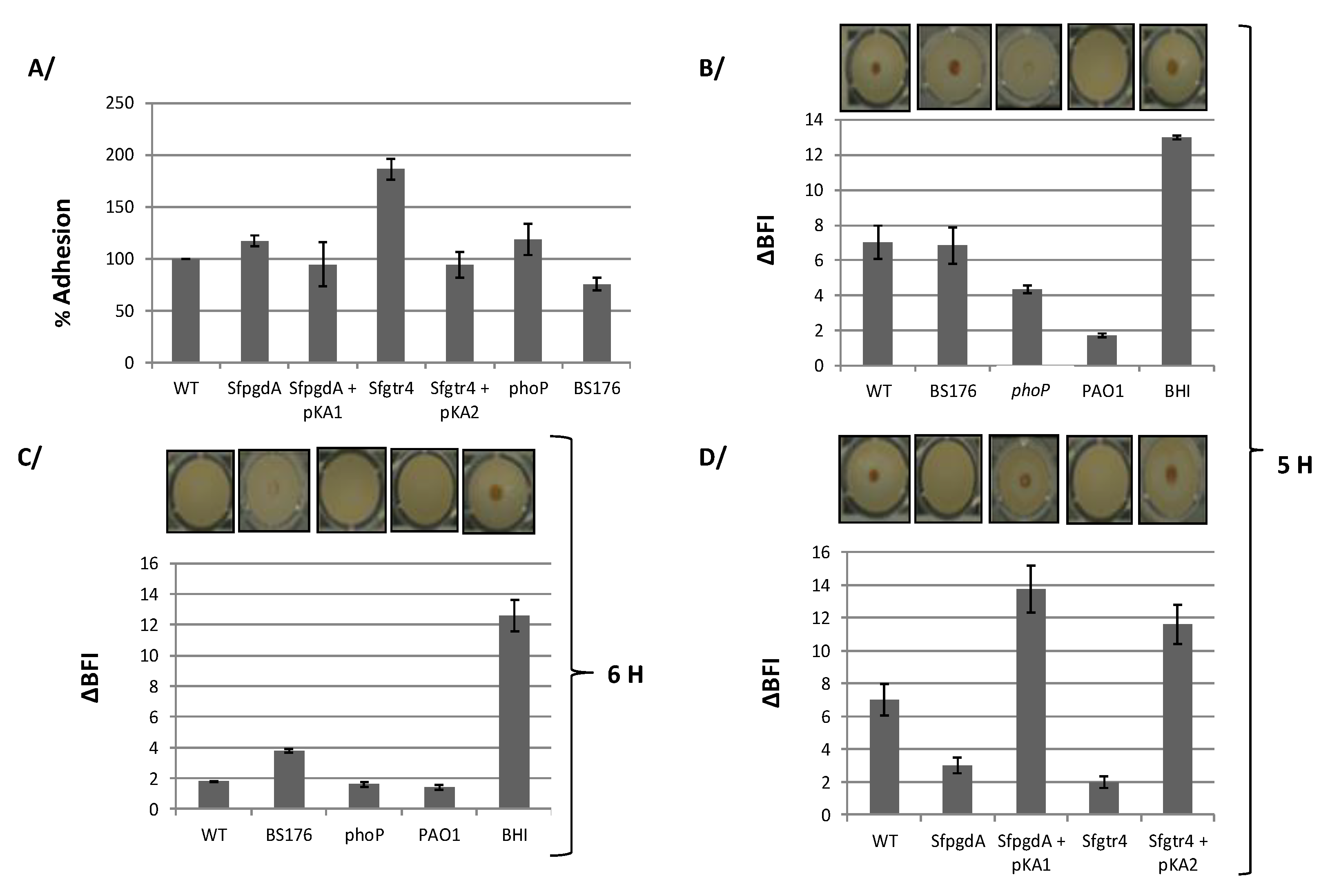
3.2. Inactivation of Sfgtr4 or SfpgdA Gene Reduces Shigella Entry into HeLa Cells Without Impairing Effectors Secretion via the T3SA
3.3. Inactivation of Sfgtr4 Gene Enhances Sensitivity to Lysozyme Action and Increases Bacterial Killing by Polymorphonuclear Neutrophils (PMNs)
3.4. Inactivation of Sfgtr4 or SfpgdA Accelerates Biofilm Appearance
3.5. The phoP Mutant, but not the pWR100 Shigella Cured Strain, Increases Biofilm Formation
3.6. SfGtr4 Interacts with SfPgdA
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buchrieser, C.; Glaser, P.; Rusniok, C.; Nedjari, H.; D’Hauteville, H.; Kunst, F.; Sansonetti, P.; Parsot, C. The virulence plasmid pWR100 and the repertoire of proteins secreted by the type III secretion apparatus of Shigella flexneri. Mol. Microbiol. 2000, 38, 760–771. [Google Scholar] [CrossRef] [PubMed]
- Parsot, C. Shigella type III secretion effectors: How, where, when, for what purposes? Curr. Opin. Microbiol. 2009, 12, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Fujiyama, R.; Nishi, J.; Imuta, N.; Tokuda, K.; Manago, K.; Kawano, Y. The shf Gene of a Shigella flexneri homologue on the virulent plasmid pAA2 of enteroaggregative escherichia coli 042 Is required for firm biofilm formation. Curr. Microbiol. 2008, 56, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Forman, S.; Bobrov, A.G.; Kirillina, O.; Craig, S.K.; Abney, J.; Fetherston, J.D.; Perry, R.D. Identification of critical amino acid residues in the plague biofilm Hms proteins. Microbiology 2006, 152, 3399–3410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, Y.; Rice, J.D.; Goller, C.; Pannuri, A.; Taylor, J.; Meisner, J.; Beveridge, T.J.; Preston, J.F.; Romeo, T. Roles of pgaABCD Genes in synthesis, modification, and export of the escherichia coli biofilm adhesin Poly-β-1,6-N-Acetyl-d-Glucosamine. J. Bacteriol. 2008, 190, 3670–3680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, R.D.; Bobrov, A.G.; Kirillina, O.; Jones, H.A.; Pedersen, L.; Abney, J.; Fetherston, J.D. Temperature regulation of the Hemin storage (Hms+) phenotype of Yersinia pestis is posttranscriptional. J. Bacteriol. 2004, 186, 1638–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Cromie, M.J.; Hsu, F.-F.; Turk, J.; Groisman, E.A. PhoP-regulated Salmonella resistance to the antimicrobial peptides magainin 2 and polymyxin B. Mol. Microbiol. 2004, 53, 229–241. [Google Scholar] [CrossRef]
- Vuong, C.; Kocianova, S.; Voyich, J.M.; Yao, Y.; Fischer, E.R.; DeLeo, F.R.; Otto, M. A Crucial role for exopolysaccharide modification in bacterial biofilm formation, immune evasion, and virulence. J. Boil. Chem. 2004, 279, 54881–54886. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, G.A.; Kaplan, H.B.; Kolter, R. Biofilm formation as microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef]
- Goldman, S.R.; Tu, Y.; Goldberg, M.B. Differential regulation by magnesium of the two MsbB paralogs of Shigella flexneri. J. Bacteriol. 2008, 190, 3526–3537. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Cai, X.; Chen, M.; Ye, L.; Wu, Y.; Wang, X.; Lv, Z.; Shang, Y.; Qu, D. Virulence and stress responses of Shigella flexneri regulated by PhoP/PhoQ. Front. Microbiol. 2018, 8, 2689. [Google Scholar] [CrossRef] [PubMed]
- Kaoukab-Raji, A.; Biskri, L.; Bernardini, M.; Allaoui, A. Characterization of SfPgdA, a Shigella flexneri peptidoglycan deacetylase required for bacterial persistence within polymorphonuclear neutrophils. Microbes Infect. 2012, 14, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Makino, K.; Ishii, K.; Yasunaga, T.; Hattori, M.; Yokoyama, K.; Yutsudo, C.H.; Kubota, Y.; Yamaichi, Y.; Iida, T.; Yamamoto, K.; et al. Complete nucleotide sequences of 93-kb and 3.3-kb plasmids of an enterohemorrhagic Escherichia coli O157:H7 derived from Sakai outbreak. Curr. Neuropharmacol. 1998, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yethon, J.A.; Vinogradov, E.; Perry, M.B.; Whitfield, C. Mutation of the lipopolysaccharide core glycosyltransferase encoded by waaG destabilizes the outer membrane of escherichia coli by interfering with core phosphorylation. J. Bacteriol. 2000, 182, 5620–5623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S. Glycosyltransferases: Structures, Functions, and Mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allaoui, A.; Sansonetti, P.J.; Parsot, C. MxiJ, a lipoprotein involved in secretion of Shigella Ipa invasins, is homologous to YscJ, a secretion factor of the Yersinia Yop proteins. J. Bacteriol. 1992, 174, 7661–7669. [Google Scholar] [CrossRef] [Green Version]
- Demarre, G.; Guerout, A.-M.; Matsumoto-Mashimo, C.; Rowe-Magnus, D.; Marlière, P.; Mazel, D. A new family of mobilizable suicide plasmids based on broad host range R388 plasmid (IncW) and RP4 plasmid (IncPα) conjugative machineries and their cognate Escherichia coli host strains. Res. Microbiol. 2005, 156, 245–255. [Google Scholar] [CrossRef]
- Sansonetti, P.J.; Ryter, A.; Clerc, P.; Maurelli, A.T.; Mounier, J. Multiplication of Shigella flexneri within HeLa cells: Lysis of the phagocytic vacuole and plasmid-mediated contact hemolysis. Infect. Immun. 1986, 51, 461–469. [Google Scholar] [CrossRef] [Green Version]
- Allaoui, A.; Sansonetti, P.J.; Parsot, C. MxiD, an outer membrane protein necessary for the secretion of the Shigella flexneri Ipa invasins. Mol. Microbiol. 1993, 7, 59–68. [Google Scholar] [CrossRef]
- Ménard, R.; Sansonetti, P.J.; Parsot, C. Nonpolar mutagenesis of the ipa genes defines IpaB, IpaC, and IpaD as effectors of Shigella flexneri entry into epithelial cells. J. Bacteriol. 1993, 175, 5899–5906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magdalena, J.; Hachani, A.; Chamekh, M.; Jouihri, N.; Gounon, P.; Blocker, A.; Allaoui, A. Spa32 regulates a switch in substrate specificity of the Type III secreton of Shigella flexneri from needle components to Ipa Proteins. J. Bacteriol. 2002, 184, 3433–3441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barzu, S.; Nato, F.; Rouyre, S.; Mazie, J.C.; Sansonetti, P.; Phalipon, A. Characterization of B-cell epitopes on IpaB, an invasion-associated antigen of Shigella flexneri: Identification of an immunodominant domain recognized during natural infection. Infect. Immun. 1993, 61, 3825–3831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phalipon, A.; Arondel, J.; Nato, F.; Rouyre, S.; Mazie, J.C.; Sansonetti, P.J. Identification and characterization of B-cell epitopes of IpaC, an invasion-associated protein of Shigella flexneri. Infect. Immun. 1992, 60, 1919–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; Singh, V.; More, T. Electrophoretic detection of myeloperoxidase, protease, lactoferrin and lysozyme in buffalo polymorphonuclear granular acid extracts. Veter- Res. Commun. 1997, 21, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Pycock, J.; Allen, W.E.; Morris, T. Rapid, single-step isolation of equine neutrophils on a discontinuous Percoll density gradient. Res. Veter- Sci. 1987, 42, 411–412. [Google Scholar] [CrossRef]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Chavant, P.; Gaillard-Martinie, B.; Talon, R.; Hebraud, M.; Bernardi, T. A new device for rapid evaluation of biofilm formation potential by bacteria. J. Microbiol. Methods 2007, 68, 605–612. [Google Scholar] [CrossRef]
- Chen, L.; Men, H.; Ha, S.; Ye, X.-Y.; Brunner, L.; Hu, Y.; Walker, S. Intrinsic Lipid Preferences and Kinetic Mechanism ofEscherichia coliMurG†. Biochemistry 2002, 41, 6824–6833. [Google Scholar] [CrossRef]
- Maira-Litrán, T.; Kropec, A.; Abeygunawardana, C.; Joyce, J.; Mark, G.; Goldmann, D.A.; Pier, G.; Iii, G.M. Immunochemical properties of the staphylococcal Poly-N-Acetylglucosamine surface polysaccharide. Infect. Immun. 2002, 70, 4433–4440. [Google Scholar] [CrossRef] [Green Version]
- Cramton, S.E.; Gerke, C.; Schnell, N.F.; Nichols, W.W.; Götz, F. The Intercellular Adhesion (ica) Locus Is present in staphylococcus aureus and is required for biofilm formation. Infect. Immun. 1999, 67, 5427–5433. [Google Scholar] [CrossRef] [Green Version]
- Mercier, C.; Durrieu, C.; Briandet, R.; Domakova, E.; Tremblay, J.; Buist, G.; Kulakauskas, S. Positive role of peptidoglycan breaks in lactococcal biofilm formation. Mol. Microbiol. 2002, 46, 235–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelis, G.R. The type III secretion injectisome. Nat. Rev. Genet. 2006, 4, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Hueck, C.J. Type III protein secretion systems in bacterial pathogens of animals and plants. Microbiol. Mol. Boil. Rev. 1998, 62, 379–433. [Google Scholar] [CrossRef] [Green Version]
- West, N.P.; Sansonetti, P.; Mounier, J.; Exley, R.M.; Parsot, C.; Guadagnini, S.; Prévost, M.-C.; Prochnicka-Chalufour, A.; Delepierre, M.; Tanguy, M.; et al. Optimization of virulence functions through glucosylation of Shigella LPS. Science 2005, 307, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.B.; Mahapatra, S.; Crick, D.C.; Pavelka, M.S. Identification of thenamHGene, encoding the hydroxylase responsible for the N-Glycolylation of the mycobacterial peptidoglycan. J. Boil. Chem. 2004, 280, 326–333. [Google Scholar] [CrossRef] [Green Version]
- Nickerson, K.P.; Chanin, R.B.; Sistrunk, J.R.; Rasko, D.A.; Fink, P.J.; Barry, E.M.; Nataro, J.P.; Faherty, C.S. Analysis of Shigella flexneri resistance, biofilm formation, and transcriptional profile in response to bile salts. Infect. Immun. 2017, 85, e01067:1–e01067:16. [Google Scholar] [CrossRef] [Green Version]
- Köseoğlu, V.K.; Hall, C.P.; Rodríguez-López, E.M.; Agaisse, H. The autotransporter IcsA promotes Shigella flexneri biofilm formation in the presence of bile salts. Infect. Immun. 2019, 87, e00861–e008618. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.M.; Jackson, M.D.; Akerley, B.J. Suppression of alternative lipooligosaccharide Glycosyltransferase activity by UDP-Galactose epimerase enhances murine lung infection and evasion of serum IgM. Front. Microbiol. 2019, 9, 160. [Google Scholar] [CrossRef]
- Weinrauch, Y.; Drujan, D.; Shapiro, S.D.; Weiss, J.; Zychlinsky, A. Neutrophil elastase targets virulence factors of enterobacteria. Nature 2002, 417, 91. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid | Characteristics | Reference |
| pKA1 | pTZ18R encoding native SfPgdA | [12] |
| pKA2 | pTZ18R encoding native SfGtr4 | TS |
| pKA85 | pTZ18R encoding native SfPgdA and SfGtr4 | TS |
| pKA4 | pSW23T- SfpgdA::aphA-3 (Suicide vector) | [12] |
| pKA8 | pSW23T- Sfgtr4::aphA-3 (Suicide vector) | TS |
| pKA121 | pSW23T- SfPgdA -Sfgtr4::aphA-3 (Suicide vector) | TS |
| pKA20 | pMALCRI expressing MBP-SfPgdA | TS |
| pKA22 | pQE60 expressing SfPgdA-His6 | TS |
| pKA23 | pQE60 expressing SfpgDA-His6 | TS |
| Strain | Genotype | Reference |
| M90T-Sm | Derivative of wild-type Shigella flexneri strain M90T | [16] |
| BS176 | M90T-Sm pWR100 cured strain | [18] |
| SF401 | M90T-Sm mxiD (mxiD::aphA-3) | [19] |
| SBkr1 | M90T-Sm SfpgdA (SfpgdA::aphA-3) | [12] |
| SBkr2 | M90T-Sm Sfgtr4 (Sfgtr4::aphA-3) | TS |
| SBkr3 | M90T-Sm SfpgdA Sfgtr4 (SfpgdA-Sfgtr4::aphA-3) | TS |
| PhoP | M90T-Sm phoP (phoP::aphA-3) | [12] |
| Name | Primers 5′–3′ | R.S |
|---|---|---|
| sfpgdA.1 | CGGGATCCTTTAAACGAAGGGGGCATTTTG * | BamHI |
| sfgtr4.1 | CGGGATCCGGAAAGTTGCGCATGGCTG | BamHI |
| sfgtr4.2 | ACGCGTCGACCCGATAAATGATAAGTTACTTAC | SalI |
| sfgtr4his-s * | GTCCATGGGATTAAACGAAGGGGGCATTTT | NcoI |
| sfgtr4his-as ** | GCTAGATCTCATCCGGTAATCTTGGCCCC | BglII |
| sfpgdAmal-s | CGGGATCCTTAAACGAAGGGGGCATT | BamHI |
| sfpgdAmal-as | AACTGCAGTTAATCATCCGGTAATCTTGGC | PstI |
| sfpgdAhis-s | GTCCATGGGAAATATACTATTTACGGAATCAT | NcoI |
| sfpgdAhis-as | GCTAGATCTCTTGTGCTTCGCTAATGTGAG | BglII |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaoukab-Raji, A.; Biskri, L.; Allaoui, A. Inactivation of the sfgtr4 Gene of Shigella flexneri Induces Biofilm Formation and Affects Bacterial Pathogenicity. Microorganisms 2020, 8, 841. https://doi.org/10.3390/microorganisms8060841
Kaoukab-Raji A, Biskri L, Allaoui A. Inactivation of the sfgtr4 Gene of Shigella flexneri Induces Biofilm Formation and Affects Bacterial Pathogenicity. Microorganisms. 2020; 8(6):841. https://doi.org/10.3390/microorganisms8060841
Chicago/Turabian StyleKaoukab-Raji, Abdelmoughit, Latéfa Biskri, and Abdelmounaaïm Allaoui. 2020. "Inactivation of the sfgtr4 Gene of Shigella flexneri Induces Biofilm Formation and Affects Bacterial Pathogenicity" Microorganisms 8, no. 6: 841. https://doi.org/10.3390/microorganisms8060841
APA StyleKaoukab-Raji, A., Biskri, L., & Allaoui, A. (2020). Inactivation of the sfgtr4 Gene of Shigella flexneri Induces Biofilm Formation and Affects Bacterial Pathogenicity. Microorganisms, 8(6), 841. https://doi.org/10.3390/microorganisms8060841