Transcriptomic and Ultrastructural Signatures of K+-Induced Aggregation in Phytophthora parasitica Zoospores

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zoospore Suspension Preparation and K+ Gradient Generation
2.2. RNA Extraction and Sequencing
2.3. Transcriptomic Analysis
2.4. cDNA Preparation and RT-qPCR Analysis
2.5. Statistical Analysis
2.6. Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM)
2.7. Protonography of Carbonic Anhydrase (CA) Enzymatic Activity
2.8. Immunolocalization
2.9. Verapamil Pharmacological Assay
2.10. Image Analysis
3. Results
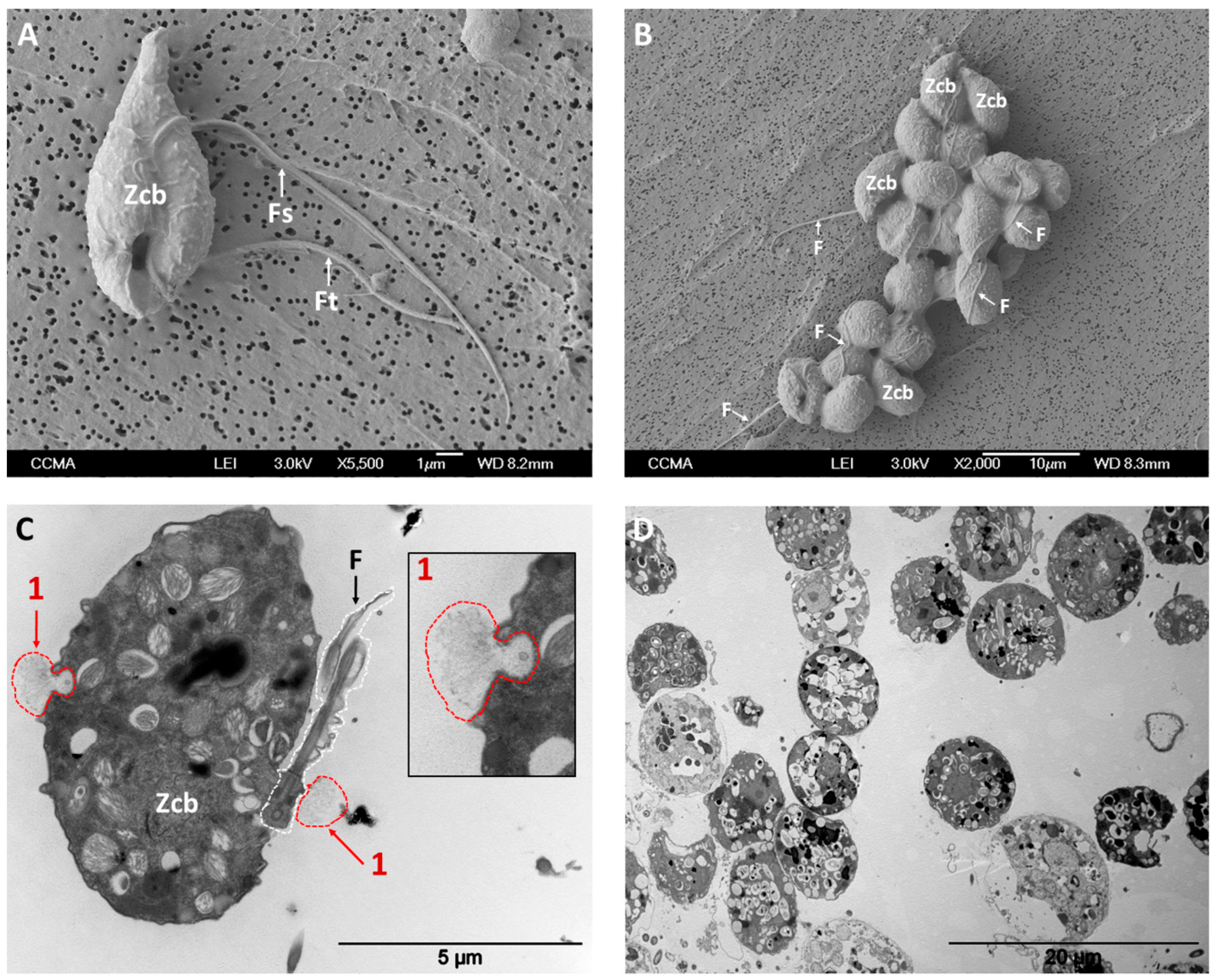
3.1. Ultrastructures of Zoospore Aggregates
3.2. Overview of the Transcriptome of Swimming Zoospores
3.3. Molecular Patterns Underlying Coordinated Behavior and Cell-to-Cell Adhesion of P. parasitica Zoospores
3.4. P. parasitica Zoospore Transcriptome Dynamics and Cell Response to K+ Gradient Application
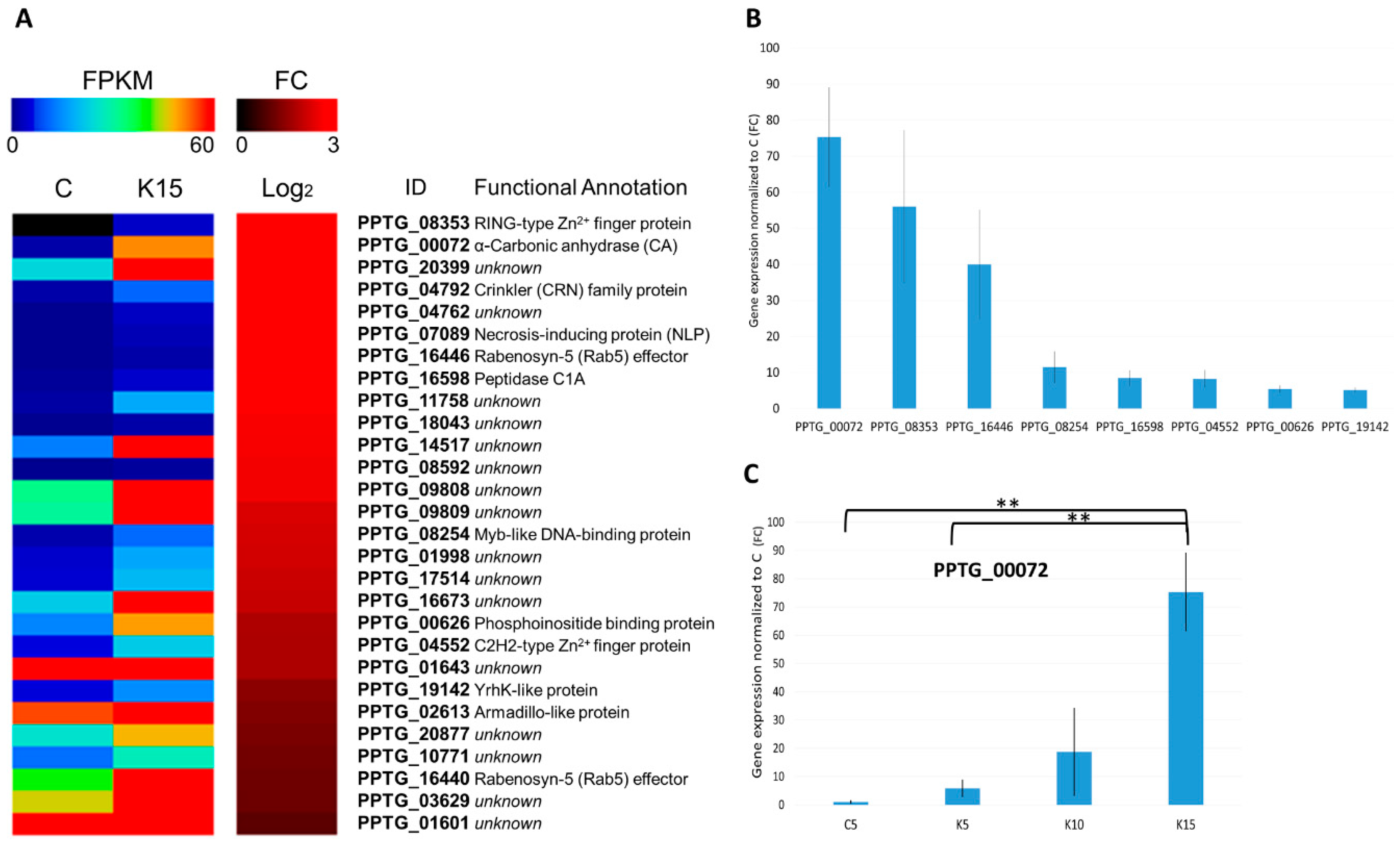
3.4.1. Overview of K+-Induced Transcriptome Dynamics
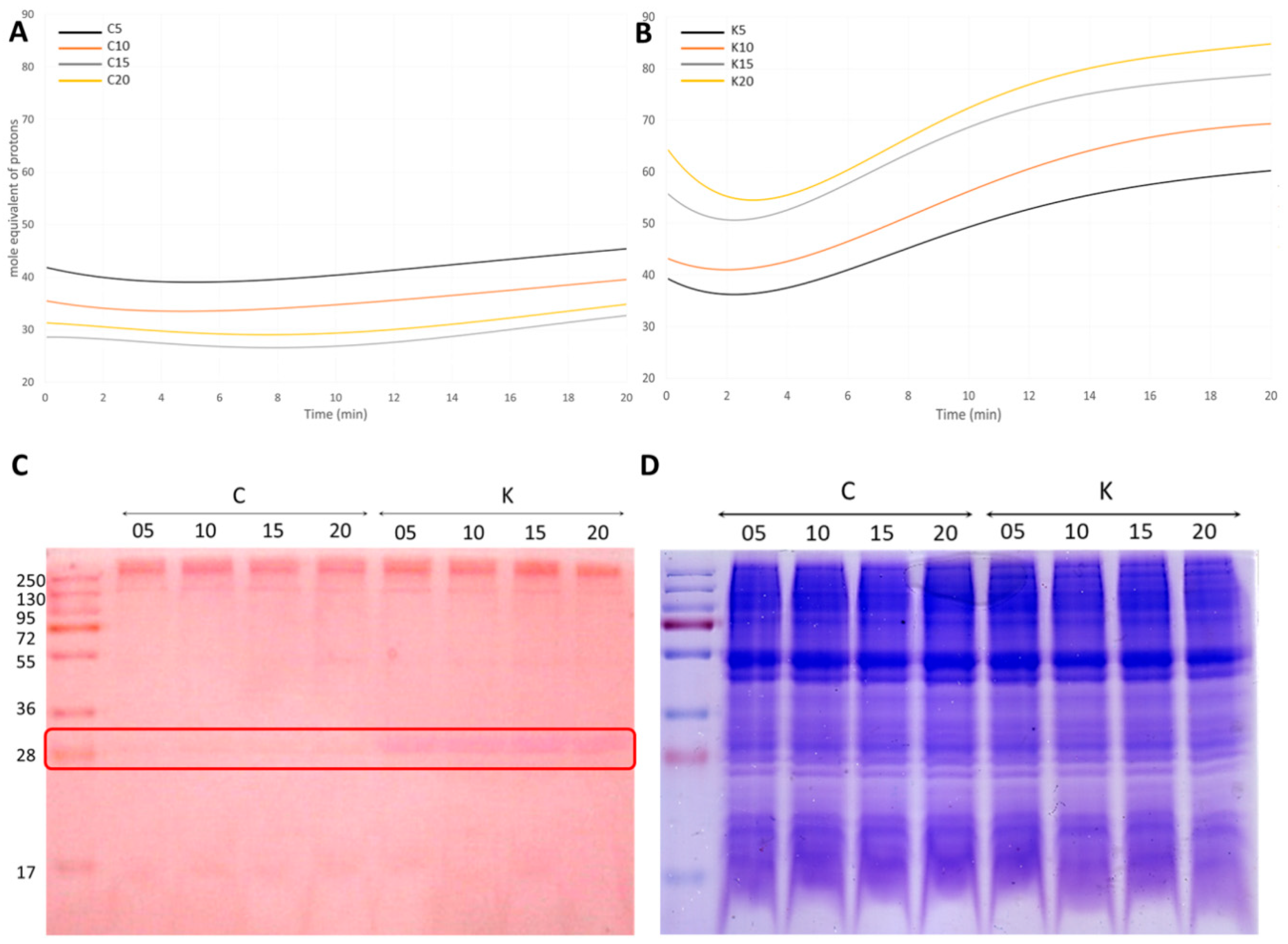
3.4.2. A Carbonic Anhydrase (CA) Is Strongly Induced upon K+ Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beakes, G.W.; Glockling, S.L.; Sekimoto, S. The evolutionary phylogeny of the oomycete “fungi”. Protoplasma 2012, 249, 3–19. [Google Scholar] [CrossRef]
- Judelson, H.S.; Blanco, F.A. The spores of Phytophthora: Weapons of the plant destroyer. Nat. Rev. Microbiol. 2005, 3, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, S.; Furzer, O.; Jones, J.D.G.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.D.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabières, F.; et al. The Top 10 oomycete pathogens in molecular plant pathology. Mol. Plant Pathol. 2015, 16, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.A.; van West, P. Zoospore development in the oomycetes. Fungal Biol. Rev. 2007, 21, 10–18. [Google Scholar] [CrossRef]
- Galiana, E.; Fourré, S.; Engler, G. Phytophthora parasitica biofilm formation: Installation and organization of microcolonies on the surface of a host plant. Environ. Microbiol. 2008, 10, 2164–2171. [Google Scholar] [CrossRef] [PubMed]
- Larousse, M.; Govetto, B.; Séassau, A.; Etienne, C.; Industri, B.; Theodorakopoulos, N.; Deleury, E.; Ponchet, M.; Panabières, F.; Galiana, E. Characterization of PPMUCL1/2/3, three members of a new oomycete-specific mucin-like protein family residing in Phytophthora parasitica biofilm. Protist 2014, 165, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Galiana, E.; Cohen, C.; Thomen, P.; Etienne, C.; Noblin, X. Guidance of zoospores by potassium gradient sensing mediates aggregation. J. R. Soc. Interface 2019, 16, 20190367. [Google Scholar] [CrossRef] [Green Version]
- Thomson, S.V. Mechanisms of Survival of Zoospores of Phytophthora parasitica in Irrigation Water. Phytopathology 1976, 66, 1198. [Google Scholar] [CrossRef]
- Filho, A.C.C.; Duniway, J.M. Dispersal of Phytophthora capsici and P. parasitica in furrow-irrigated rows of bell pepper, tomato and squash. Plant Pathol. 1995, 44, 1025–1032. [Google Scholar] [CrossRef]
- Kemen, E. Microbe-microbe interactions determine oomycete and fungal host colonization. Curr. Opin. Plant Biol. 2014, 20, 75–81. [Google Scholar] [CrossRef]
- Larousse, M.; Galiana, E. Microbial Partnerships of Pathogenic Oomycetes. PLoS Pathog. 2017, 13, e1006028. [Google Scholar] [CrossRef]
- Sebé-Pedrós, A.; Irimia, M.; del Campo, J.; Parra-Acero, H.; Russ, C.; Nusbaum, C.; Blencowe, B.J.; Ruiz-Trillo, I. Regulated aggregative multicellularity in a close unicellular relative of metazoa. eLife 2013, 2, e01287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Q.; Kawabe, Y.; Schilde, C.; Chen, Z.H.; Schaap, P. The Evolution of Aggregative Multicellularity and Cell-Cell Communication in the Dictyostelia. J. Mol. Biol. 2015, 427, 3722–3733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, N.; Hittinger, C.T.; Carroll, S.B. Evolution of key cell signaling and adhesion protein families predates animal origins. Science 2003, 301, 361–363. [Google Scholar] [CrossRef]
- King, N.; Westbrook, M.J.; Young, S.L.; Kuo, A.; Abedin, M.; Chapman, J.; Fairclough, S.; Hellsten, U.; Isogai, Y.; Letunic, I.; et al. The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans. Nature 2008, 451, 783–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Judelson, H.S.; Ah-Fong, A.M.V. Exchanges at the plant-oomycete interface that influence disease. Plant Physiol. 2019, 179, 1198–1211. [Google Scholar] [CrossRef] [Green Version]
- Stanghellini, M.E.; Rasmussen, S.L. Hydroponics: A solution for zoosporic pathogens. Plant Dis. 1994, 78, 1129–1138. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Howe, E.; Holton, K.; Nair, S.; Schlauch, D.; Sinha, R.; Quackenbush, J. MeV: MultiExperiment viewer. In Biomedical Informatics for Cancer Research; Springer: Boston, MA, USA, 2010; pp. 267–277. [Google Scholar]
- Bateman, A. The Pfam protein families database. Nucleic Acids Res. 2004, 32, 138–141. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbon, S.; Douglass, E.; Dunn, N.; Good, B.M.; Harris, N.L.; Lewis, S.E.; Mungall, C.J.; Basu, S.N.; Chisholm, R.L.; Dodson, R.J.; et al. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bellono, N.W.; Leitch, D.B.; Julius, D. Molecular basis of ancestral vertebrate electroreception. Nature 2017, 543, 391–396. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Smirnov, S.; Nikolskaya, A.N.; et al. The COG database: An updated vesion includes eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, N.D.; Waller, M.; Barrett, A.J.; Bateman, A. MEROPS: The database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2014, 42, D503–D509. [Google Scholar] [CrossRef] [Green Version]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakai, K.; Horton, P. PSORT: A program for detecting sorting signals in proteins and predicting their subcellular localization. Trends Biochem. Sci. 1999, 24, 34–35. [Google Scholar] [CrossRef]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Evangelisti, E.; Govetto, B.; Minet-Kebdani, N.; Kuhn, M.L.; Attard, A.; Ponchet, M.; Panabières, F.; Gourgues, M. The Phytophthora parasitica RXLR effector Penetration-Specific Effector 1 favours Arabidopsis thaliana infection by interfering with auxin physiology. New Phytol. 2013, 199, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Dalio, R.J.D.; Maximo, H.J.; Oliveira, T.S.; Dias, R.O.; Breton, M.C.; Felizatti, H.; Machado, M. Phytophthora parasitica Effector PpRxLR2 Suppresses Nicotiana benthamiana Immunity. Mol. Plant Microbe Interact. 2018, 31, 481–493. [Google Scholar] [CrossRef] [Green Version]
- le Berre, J.-Y.; Gourgues, M.; Samans, B.; Keller, H.; Panabières, F.; Attard, A. Transcriptome dynamic of Arabidopsis roots infected with Phytophthora parasitica identifies VQ29, a gene induced during the penetration and involved in the restriction of infection. PLoS ONE 2017, 12, e0190341. [Google Scholar] [CrossRef] [Green Version]
- Team, R. R: A Language and Environment for Statistical Computing; Austria R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open software development for computational biology and bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.Y.; Dillies, M.A. SARTools: A DESeq2- and edgeR-based R pipeline for comprehensive differential analysis of RNA-Seq data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Yekutieli, D. The Control of the False Discovery Rate in Multiple Testing under Dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Rancurel, C.; van Tran, T.; Elie, C.; Hilliou, F. SATQPCR: Website for statistical analysis of real-time quantitative PCR data. Mol. Cell Probes 2019, 46, 101418. [Google Scholar] [CrossRef] [PubMed]
- De Luca, V.; del Prete, S.; Supuran, C.T.; Capasso, C. Protonography, a new technique for the analysis of carbonic anhydrase activity. J. Enzym. Inhib. Med. Chem. 2015, 30, 277–282. [Google Scholar] [CrossRef]
- Zhang, W.; Blackman, L.M.; Hardham, A.R. Transient fusion and selective secretion of vesicle proteins in Phytophthora nicotianae zoospores. PeerJ 2013, 1, e221. [Google Scholar] [CrossRef] [Green Version]
- Niklas, K.J.; Newman, S.A. The origins of multicellular organisms. Evol. Dev. 2013, 15, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Hulpiau, P.; Gul, I.S.; van Roy, F. New insights into the evolution of metazoan cadherins and catenins. In Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2013; Volume 116, pp. 71–94. [Google Scholar]
- Ah-Fong, A.M.V.; Kim, K.S.; Judelson, H.S. RNA-seq of life stages of the oomycete Phytophthora infestans reveals dynamic changes in metabolic, signal transduction, and pathogenesis genes and a major role for calcium signaling in development. BMC Genom. 2017, 18, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, R.H.Y.; Tyler, B.M.; Whisson, S.C.; Hardham, A.R.; Govers, F. Ancient Origin of Elicitin Gene Clusters in Phytophthora Genomes. Mol. Biol. Evol. 2006, 23, 338–351. [Google Scholar] [CrossRef] [Green Version]
- Panabieres, F.; Marais, A.; le Berre, J.Y.; Penot, I.; Fournier, D.; Ricci, P. Characterization of a gene cluster of Phytophthora cryptogea which codes for elicitins, proteins inducing a hypersensitive-like response in tobacco. Mol. Plant Microbe Interact. 1995, 8, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Amaro, T.M.M.M.; Thilliez, G.J.A.; Motion, G.B.; Huitema, E. A Perspective on CRN Proteins in the Genomics Age: Evolution, Classification, Delivery and Function Revisited. Front. Plant Sci. 2017, 8, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.H.; Yan, H.Z.; Liou, R.F. A novel elicitor protein from Phytophthora parasitica induces plant basal immunity and systemic acquired resistance. Mol. Plant Pathol. 2015, 16, 123–136. [Google Scholar] [CrossRef]
- Blackman, L.M.; Cullerne, D.P.; Hardham, A.R. Bioinformatic characterisation of genes encoding cell wall degrading enzymes in the Phytophthora parasitica genome. BMC Genom. 2014, 15, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Ambikapathy, J.; Marshall, J.S.; Hocart, C.H.; Hardham, A.R. The role of proline in osmoregulation in Phytophthora nicotianae. Fungal Genet. Biol. 2002, 35, 287–299. [Google Scholar] [CrossRef]
- Škalamera, D.; Hardham, A.R. PnCcp, a Phytophthora nicotianae protein containing a single complement control protein module, is sorted into large peripheral vesicles in zoospores. Australas. Plant Pathol. 2006, 35, 593–603. [Google Scholar] [CrossRef]
- De Valck, D.; Heyninck, K.; van Criekinge, W.; Contreras, R.; Beyaert, R.; Fiers, W. A20, an inhibitor of cell death, self-associates by its zinc finger domain. FEBS Lett. 1996, 384, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.D.; Cho, Y.H.; Yoo, S.D. Regulatory functions of evolutionarily conserved AN1/A20-like Zinc finger family proteins in Arabidopsis stress responses under high temperature. Biochem. Biophys. Res. Commun. 2015, 457, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Di Vietro, L.; Daghino, S.; Abbà, S.; Perotto, S. Gene expression and role in cadmium tolerance of two PLAC8-containing proteins identified in the ericoid mycorrhizal fungus Oidiodendron maius. Fungal Biol. 2014, 118, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, U.M.; Manohar, M.; Gaur, V.S. Calcium transport from source to sink: Understanding the mechanism(s) of acquisition, translocation, and accumulation for crop biofortification. Acta Physiol. Plant. 2015, 37, 1722. [Google Scholar] [CrossRef]
- Fletcher, K.I.G.; van West, P.; Gachon, C.M.M. Nonagonal cadherins: A new protein family found within the Stramenopiles. Gene 2016, 593, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Humphries, J.; Xiong, L.; Liu, J.; Prindle, A.; Yuan, F.; Arjes, H.A.; Tsimring, L.; Süel, G.M. Species-Independent Attraction to Biofilms through Electrical Signaling. Cell 2017, 168, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Borden, K.L.B.; Freemont, P.S. The RING finger domain: A recent example of a sequence-structure family. Curr. Opin. Struct. Biol. 1996, 6, 395–401. [Google Scholar] [CrossRef]
- Wishart, M.J.; Taylor, G.S.; Dixon, J.E. Phoxy lipids: Revealing PX domains as phosphoinositide binding modules. Cell 2001, 105, 817–820. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, E.; Christoforidis, S.; Uttenweiler-Joseph, S.; Miaczynska, M.; Dewitte, F.; Wilm, M.; Hoflack, B.; Zerial, M. Rabenosyn-5, a novel Rab5 effector, is complexed with hVPS45 and recruited to endosomes through a FYVE finger domain. J. Cell Biol. 2000, 151, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gijzen, M.; Nürnberger, T. Nep1-like proteins from plant pathogens: Recruitment and diversification of the NPP1 domain across taxa. Phytochemistry 2006, 67, 1800–1807. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef] [PubMed]
- Brayer, K.J.; Segal, D.J. Keep your fingers off my DNA: Protein-protein interactions mediated by C2H2 zinc finger domains. Cell Biochem. Biophys. 2008, 50, 111–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, L.A.; Latek, R.R.; Peterson, C.L. The SANT domain: A unique histone-tail-binding module? Nat. Rev. Mol. Cell Biol. 2004, 5, 158–163. [Google Scholar] [CrossRef]
- Purkerson, J.M.; Schwartz, G.J. The role of carbonic anhydrases in renal physiology. Kidney Int. 2007, 71, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Appiah, A.A.; van West, P.; Osborne, M.C.; Gow, N.A.R. Potassium homeostasis influences the locomotion and encystment of zoospores of plant pathogenic oomycetes. Fungal Genet. Biol. 2005, 42, 213–223. [Google Scholar] [CrossRef]
- Donaldson, S.P.; Deacon, J.W. Role of calcium in adhesion and germination of zoospore cysts of Pythium: A model to explain infection of host plants. J. Gen. Microbiol. 1992, 138, 2051–2059. [Google Scholar] [CrossRef] [Green Version]
- Warburton, A.J.; Deacon, J.W. Transmembrane Ca2+ fluxes associated with zoospore encystment and cyst germination by the phytopathogen Phytophthora parasitica. Fungal Genet. Biol. 1998, 25, 54–62. [Google Scholar] [CrossRef]
- Henry, R.P. Multiple Roles of Carbonic Anhydrase in Cellular Transport and Metabolism. Annu. Rev. Physiol. 1996, 58, 523–538. [Google Scholar] [CrossRef]
- Alvarez, B.V.; Loiselle, F.B.; Supuran, C.T.; Schwartz, G.J.; Casey, J.R. Direct Extracellular Interaction between Carbonic Anhydrase IV and the Human NBC1 Sodium/Bicarbonate Co-Transporter. Biochemistry 2003, 42, 12321–12329. [Google Scholar] [CrossRef]
- Yang, Z.; Alvarez, B.V.; Chakarova, C.; Jiang, L.; Karan, G.; Frederick, J.M.; Zhao, Y.; Sauvé, Y.; Li, X.; Zrenner, E.; et al. Mutant carbonic anhydrase 4 impairs pH regulation and causes retinal photoreceptor degeneration. Hum. Mol. Genet. 2005, 14, 255–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csaderova, L.; Debreova, M.; Radvak, P.; Stano, M.; Vrestiakova, M.; Kopacek, J.; Pastorekova, S.; Svastova, E. The effect of carbonic anhydrase IX on focal contacts during cell spreading and migration. Front. Physiol. 2013, 4, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Occhipinti, R.; Boron, W.F. Mathematical modeling of acid-base physiology. Prog. Biophys. Mol. Biol. 2015, 117, 43–58. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, H.J.; Hardham, A.R. Characterisation of the water expulsion vacuole in Phytophthora nicotianae zoospores. Protoplasma 1999, 206, 118–130. [Google Scholar] [CrossRef]
- Rodan, G.A.; Martin, T.J. Therapeutic approaches to bone diseases. Science 2000, 289, 1508–1514. [Google Scholar] [CrossRef]
- Vargas, L.A.; Díaz, R.G.; Swenson, E.R.; Pérez, N.G.; Álvarez, B.V. Inhibition of carbonic anhydrase prevents the Na+/H+ exchanger 1-dependent slow force response to rat myocardial stretch. Am. J. Physiol. Heart. Circ. Physiol. 2013, 305, H228–H237. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bassani, I.; Rancurel, C.; Pagnotta, S.; Orange, F.; Pons, N.; Lebrigand, K.; Panabières, F.; Counillon, L.; Noblin, X.; Galiana, E. Transcriptomic and Ultrastructural Signatures of K+-Induced Aggregation in Phytophthora parasitica Zoospores. Microorganisms 2020, 8, 1012. https://doi.org/10.3390/microorganisms8071012
Bassani I, Rancurel C, Pagnotta S, Orange F, Pons N, Lebrigand K, Panabières F, Counillon L, Noblin X, Galiana E. Transcriptomic and Ultrastructural Signatures of K+-Induced Aggregation in Phytophthora parasitica Zoospores. Microorganisms. 2020; 8(7):1012. https://doi.org/10.3390/microorganisms8071012
Chicago/Turabian StyleBassani, Ilaria, Corinne Rancurel, Sophie Pagnotta, François Orange, Nicolas Pons, Kevin Lebrigand, Franck Panabières, Laurent Counillon, Xavier Noblin, and Eric Galiana. 2020. "Transcriptomic and Ultrastructural Signatures of K+-Induced Aggregation in Phytophthora parasitica Zoospores" Microorganisms 8, no. 7: 1012. https://doi.org/10.3390/microorganisms8071012
APA StyleBassani, I., Rancurel, C., Pagnotta, S., Orange, F., Pons, N., Lebrigand, K., Panabières, F., Counillon, L., Noblin, X., & Galiana, E. (2020). Transcriptomic and Ultrastructural Signatures of K+-Induced Aggregation in Phytophthora parasitica Zoospores. Microorganisms, 8(7), 1012. https://doi.org/10.3390/microorganisms8071012