Microbial Communities in Permafrost Soils of Larsemann Hills, Eastern Antarctica: Environmental Controls and Effect of Human Impact



Abstract
:1. Introduction
- -
- To characterize taxonomic diversity and soil-physical properties of soils of Larsemann Hills;
- -
- To assess soil microbiome composition and its variability across landscapes of Larsemann Hills, as well as the role of anthropogenic factor effect on soil microbiome structure and composition;
- -
- To reveal the relationship between soil microbiome composition and diversity with soil physical–chemical properties.
2. Materials and Methods
Soil Sampling and Laboratory Analyses
3. Results and Discussion
3.1. Soil Physical–Chemical Properties
3.2. Microbiome Composition of Studied Soils
3.3. Anthropogenically-Affected Sites
3.4. Near-Natural Sites
3.5. Relationship between Soil Microbiome and Soil Physical–Chemical Properties
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vincent, W. Evolutionary origins of Antarctic microbiota: Invasion, selection, and endemism. Antarct. Sci. 2000, 12, 374–385. [Google Scholar] [CrossRef] [Green Version]
- Wynn-Williams, D. Antarctic microbial diversity: The basis of polar ecosystem processes. Biodivers. Conserv. 1996, 5, 1271–1293. [Google Scholar] [CrossRef]
- Wolińska, A.; Gałazka, A.; Kuzniar, A.; Goraj, W.; Jastrzębska, N.; Grzadziel, J.; Stepniewska, Z. Catabolic fingerprinting and diversity of bacteria in Mollic Gleysol contaminated with petroleum substances. Appl. Sci. 2018, 8, 1970. [Google Scholar] [CrossRef] [Green Version]
- Bockheim, J.; Hall, K. Permafrost, active-layer dynamics and periglacial environments of continental Antarctica. S. Afr. J. Sci. 2002, 98, 82–90. [Google Scholar]
- Mergelov, N.; Konyushkov, D.; Lupachev, A.; Goryachkin, S. Soils of Mac Robertson land. In The Soils of Antarctica; Bockheim, G., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 65–86. [Google Scholar] [CrossRef]
- Lysak, V.; Maksimova, I.A.; Nikitin, D.A.; Ivanova, A.E.; Kudinova, A.G.; Soina, V.S.; Marfenina, O.E. Soil microbial communities of eastern Antarctica. Mosc. Univ. Biol. Sci. Bull. 2018, 73, 104–112. [Google Scholar] [CrossRef]
- Friedman, E.I. Endolithic microorganisms in the Antarctic cold desert. Science 1982, 215, 1045–1053. [Google Scholar] [CrossRef]
- Kudinova, A.G.; Lysak, L.V.; Soina, V.S.; Mergelov, N.S.; Dolgikh, A.V.; Shorkunov, I.G. Bacterial communities in the soils of cryptogamic barrens of East Antarctica (the Larsemann Hills and Thala Hills oases). Eurasian Soil Sci. 2015, 48, 276–287. [Google Scholar] [CrossRef]
- Wang, N.F.; Zhang, T.; Zhang, F.; Wang, E.T.; He, J.F.; Ding, H.; Zhang, B.T.; Liu, J.; Ran, X.B.; Zan, J.Y. Diversity and structure of soil bacterial communities in the Fildes Region (maritime Antarctica) as revealed by 454 pyrosequencing. Front. Microbiol. 2015, 6, 1188. [Google Scholar] [CrossRef] [Green Version]
- Coleine, C.; Pombubpa, N.; Zucconi, L.; Onofri, S.; Turchetti, B.; Buzzini, P.; Stajich, J.E.; Selbmann, L. Uncovered Microbial Diversity in Antarctic Cryptoendolithic Communities Sampling three Representative Locations of the Victoria Land. Microorganisms 2020, 8, 942. [Google Scholar] [CrossRef]
- Coleine, C.; Pombubpa, N.; Zucconi, L.; Onofri, S.; Stajich, J.E.; Selbmann, L. Endolithic Fungal Species Markers for Harshest Conditions in the McMurdo Dry Valleys, Antarctica. Life 2020, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- De Los Rios, A.; Wierzchos, J.; Sancho, L.; Ascaso, C. Acid microenvironments in microbial biofilms of Antarctic endolithic microecosystems. Environ. Microbiol. 2003, 5, 231–237. [Google Scholar] [CrossRef] [PubMed]
- De Los Ríos, A.; Grube, M.; Sancho, L.; Ascaso, C. Ultrastructural and genetic characteristics of endolithic cyanobacterial biofilms colonizing Antarctic Granite Rocks. FEMS Microbiol. Ecol. 2007, 59, 386–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottos, E.; Scarrow, J.; Archer, S.; McDonald, I.; Cary, S. Bacterial community structures of Antarctic soils. In Antarctic Terrestrial Microbiology: Physical and Biological Properties of Antarctic Soils; Cowan, D., Ed.; Springer: Heidelberg, Germany, 2014; pp. 9–33. [Google Scholar]
- Foong, C.; Ling, C.; Gonzalez, M. Metagenomic analyses of the dominant bacterial community in the Fildes Peninsula, King George Island (South Shetland Islands). Polar Sci. 2010, 4, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Wang, G.; Zhang, Y.; Cai, M.; He, J.; Yang, H. Denaturing gradient gel electrophoresis fingerprinting of soil bacteria in the vicinity of the Chinese Great Wall Station, King George Island, Antarctica. J. Environ. Sci. 2013, 25, 1649–1655. [Google Scholar] [CrossRef]
- Chong, C.W.; Pearce, D.A.; Convey, P.; Tan, G.Y.A.; Wong, R.C.S.; Tan, I.K.P. High levels of spatial heterogeneity in the biodiversity of soil prokaryotes on Signy Island, Antarctica. Soil Biol. Biochem. 2010, 42, 601–610. [Google Scholar] [CrossRef]
- Brooks, S.T.; Jabour, J.; Bergstrom, D. What is ‘footprint’ in Antarctica: Proposing a set of definitions. Antarctic Sci. 2018, 30, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Brooks, S.T.; Jabour, J.; Van Den Hoff, J.; Bergstrom, D. Our footprint on Antarctica competes with nature for rare ice-free land. Nat. Sustain. 2019, 2, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Pertierra, L.; Hughes, K.; Vega, G.; Ollala-Tarraga, M. High Resolution Spatial Mapping of Human Footprint across Antarctica and Its Implications for the Strategic Conservation of Avifauna. PLoS ONE 2017, 12, e0168280. [Google Scholar]
- Secretariat of the Antarctic Treaty. The Protocol on Environmental Protection to the Antarctic Treaty. Available online: https://www.ats.aq/e/ep.html (accessed on 30 June 2020).
- Bajerski, F.; Wagner, D. Bacterial succession in Antarctic soils of two glacier forefields on Larsemann Hills, East Antarctica. FEMS Microbiol. Ecol. 2013, 85, 128–142. [Google Scholar] [CrossRef]
- Teixeira, L.C.; Peixoto, R.S.; Cury, J.C.; Sul, W.J.; Pellizari, V.H.; Tiedje, J.; Rosaldo, A.S. Bacterial diversity in rhizosphere soil from Antarctic vascular plants of Admiralty Bay, maritime Antarctica. ISME J. 2010, 4, 989–1001. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pershina, E.; Ivanova, E.; Abakumov, E.; Andronov, E. The impacts of deglaciation and human activity on the taxonomic structure of prokaryotic communities in Antarctic soils on King George Island. Antarct. Sci. 2018, 30, 278–288. [Google Scholar] [CrossRef]
- Mergelov, N.; Mueller, C.W.; Prater, I.; Shorkunov, I.; Dolgikh, A.; Zazovskaya, E.; Shishkov, V.; Krupskaya, V.; Abrosimov, K.; Cherkinsky, A.; et al. Alteration of rocks by endolithic organisms is one of the pathways for the beginning of soils on Earth. Sci. Rep. 2018, 8, 3367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arctic and Antarctic Research Institute database. Available online: http://www.aari.nw.ru/projects/Antarctic/data/data.asp?lang=0&station=7 (accessed on 15 June 2020).
- Mergelov, N. Soils of Wet Valleys in the Larsemann Hills and Vestfold Hills Oases (Princess Elizabeth Land, East Antarctica). Eurasian Soil Sci. 2014, 47, 845–862. [Google Scholar] [CrossRef]
- Alekseev, I.; Abakumov, E. Permafrost table depth in soils of Eastern Antarctica oases, King George and Ardley Islands (South Shetland Islands). Czech Polar Rep. 2020, 10, 7–22. [Google Scholar]
- Kachynskyi, N.A. Soil Physics; Vsshaya shkola: Moscow, Russia, 1970; 358p. [Google Scholar]
- Food and Agriculture Organization. WRB—World Reference Base for Soil Resources 2014; Update 2015; FAO: Rome, Italy, 2015; p. 203. [Google Scholar]
- Bates, S.T.; Berg-Lyons, D.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. Examining the global distribution of dominant archaeal populations in soil. ISME J. 2010, 5, 908–917. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.; Costello, E.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucl. Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Abakumov, E.; Alekseev, I. Stability of soil organic matter in Cryosols of the maritime Antarctic: Insights from 13C NMR and electron spin resonance spectroscopy. Solid Earth 2018, 9, 1329–1339. [Google Scholar] [CrossRef] [Green Version]
- Michel, R.F.M.; Schaefer, C.E.; Dias, L.E.; Simas, F.; De Melo Benites, V.; Mendonca, E.S. Ornithogenic gelisols (cryosols) from Maritime Antarctica: Pedogenesis, vegetation and carbon studies. Soil Sci. Soc. Am. J. 2006, 70, 1370–1376. [Google Scholar] [CrossRef]
- Simas, N.; Schaefer, C.; Mendonca, E.; Silva, I.; Ribeiro, A. Organic carbon stocks in permafrost-affected soils from Admiralty Bay, Antarctica. J. Res. U.S. Geol. Surv. 2007, 1047, 76–79. [Google Scholar]
- Schaefer, C.; Simas, F.; Gilkes, R.; Mathison, C.; Costa, L.; Albuquerque, M. Micromorphology and microchemistry of selected Cryosols from Maritime Antarctica. Geoderma 2008, 144, 104–115. [Google Scholar] [CrossRef]
- Abakumov, E.; Mukhametova, N. Microbial biomass and basal respiration of selected Sub-Antarctic and Antarctic soils in the areas of some Russian polar stations. Solid Earth 2014, 5, 705–712. [Google Scholar] [CrossRef] [Green Version]
- Gilichinsky, D.; Abakumov, E.; Abramov, A.; Fyodorov-Davydov, D.; Goryachkin, S.; Lupachev, A.; Mergelov, N.; Zazovskaya, E. Soils of mid and low antarctic: Diversity, geography, temperature regime. In Proceedings of the 19th World Congress of Soil Science, Soil Solutions for a Changing World, Brisbane, Australia, 1–6 August 2010; pp. 32–35. [Google Scholar]
- Espejo, W.; Celis, J.; Sandoval, M.; Gonzalez-Acuna, D.; Barra, D.; Capulin, J. The Impact of Penguins on the Content of Trace Elements and Nutrients in Coastal Soils of North Western Chile and the Antarctic Peninsula Area. Water Air Soil Pollut. 2017, 228, 116. [Google Scholar] [CrossRef]
- Metcheva, R.; Yurukova, L.; Teodorova, S. Biogenic and toxic elements in feathers, eggs, and excreta of Gentoo penguin (Pygoscelis papua ellsworthii) in the Antarctic. Environ. Monit. Assess. 2011, 182, 571–585. [Google Scholar] [CrossRef]
- Santamans, A.C.; Boluda, R.; Picazo, A.; Gil, C.; Ramos-Miras, J.; Tejedo, P.; Pertierra, L.R.; Benayas, J.; Camacho, A. Soil features in rookeries of Antarctic penguins reveal sea to land biotransport of chemical pollutants. PLoS ONE 2017, 12, e0181901. [Google Scholar] [CrossRef] [Green Version]
- Schutte, U.M.E.; Abdo, Z.; Bent, S.J.; Williams, C.J.; Schneider, G.M.; Solheim, B.; Forney, L.J. Bacterial succession in a glacier foreland of the High Arctic. ISME J. 2009, 3, 1258–1268. [Google Scholar] [CrossRef] [Green Version]
- Cannone, N.; Wagner, D.; Hubberten, H.W.; Guglielmin, M. Biotic and abiotic factors influencing soil properties across a latitudinal gradient in Victoria Land, Antarctica. Geoderma 2008, 144, 50–65. [Google Scholar] [CrossRef] [Green Version]
- Pointing, S.B.; Chan, Y.; Lacap, D.C.; Lau, M.C.Y.; Jurgens, J.A.; Farrel, R.L. Highly specialized microbial diversity in hyper-arid polar desert. Proc. Natl. Acad. Sci. USA 2009, 106, 19964–19969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yung, C.C.M.; Chan, Y.; Lacap, D.C.; Perez-Ortega, S.; de los Rios-Murillo, A.; Lee, C.K.; Cary, S.K.; Pointing, S.B. Characterization of chasmoendolithic community in Miers Valley, McMurdo DryValleys, Antarctica. Microb. Ecol. 2014, 68, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Archer, D.; de los Rios, A.; Lee, K.; Niederberger, T.; Cary, S.; Coyne, K.; Douglas, S.; Lacap-Bugler, D.; Pointing, S. Endolithic microbial diversity in sandstone and granite from the McMurdo Dry Valleys, Antarctica. Polar Biol. 2016, 40, 997–1006. [Google Scholar] [CrossRef]
- Aislabie, J.M.; Chhour, K.L.; Saul, D.J.; Miyauchi, S.; Ayton, J.; Paetzold, R.F.; Balks, M.R. Dominant bacteria in soils of Marble Point and Wright Valley, Victoria Land, Antarctica. Soil Biol. Biochem. 2006, 38, 3041–3056. [Google Scholar] [CrossRef]
- Niederberger, T.D.; McDonald, I.R.; Hacker, A.L.; Soo, R.M.; Barrett, J.E.; Wall, D.H.; Cary, S.C. Microbial community composition in soils of Northern Victoria Land, Antarctica. Environ. Microbiol. 2008, 10, 1713–1724. [Google Scholar] [CrossRef] [PubMed]
- Saul, D.J.; Aislabie, J.M.; Brown, C.E.; Harris, L.; Foght, J.M. Hydrocarbon contamination changes the bacterial diversity of soil from around Scott Base, Antarctica. FEMS Microbiol. Ecol. 2005, 53, 141–155. [Google Scholar] [CrossRef] [Green Version]
- Aislabie, J.M.; Jordan, S.; Barker, G.M. Relation between soil classification and bacterial diversity in soils of the Ross Sea region, Antarctica. Geoderma 2008, 144, 9–20. [Google Scholar] [CrossRef]
- Shivaji, S.; Reddy, G.S.; Aduri, R.P.; Kutty, R.; Ravenschlag, K. Bacterial diversity of a soil sample from Schirmacher Oasis, Antarctica. Cell. Mol. Biol. 2004, 50, 525–536. [Google Scholar]
- Christner, B.C.; Kvitko, B.H.; Reeve, J.N. Molecular identification of bacteria and eukarya inhabiting an Antarctic cryoconite hole. Extremophiles 2003, 7, 177–183. [Google Scholar] [CrossRef]
- Mummey, D.L.; Stahl, P.D. Candidate division BD:phylogeny, distribution and abundance in soil ecosystems. Syst. Appl. Microbiol. 2003, 26, 228–235. [Google Scholar] [CrossRef]
- Zhang, H.; Sekiguchi, Y.; Hanada, S.; Hugenholtz, P.; Kim, H.; Kamagata, Y.; Nakamura, K. Gemmatimonas aurantiaca gen. nov., sp. nov., a Gram-negative, aerobic, polyphosphate-accumulating micro-organism, the first cultured representative of the new bacterial phylum Gemmatimonadetes phyl. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Ganzert, L.; Lipski, A.; Hubberten, H.W.; Wagner, D. The impact of different soil parameters on the community structure of dominant bacteria from nine different soils located on Livingston Island, South Shetland Archipelago, Antarctica. FEMS Microbiol. Ecol. 2011, 76, 476–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Chen, L.; Zhang, J.B.; Yin, J.; Huang, S.M. Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Front. Microbiol. 2017, 8, 187. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Kuramae, E.E.; Klinkhamer, P.G.; van Veen, J.A. Revisiting the dilution procedure used to manipulate microbial biodiversity in terrestrial systems. Appl. Environ. Microbiol. 2015, 81, 4246–4252. [Google Scholar] [CrossRef] [Green Version]
- Alger, A.S.; McKnight, D.M.; Spaulding, S.A.; Tate, C.M.; Shupe, G.H.; Welch, K.A.; Edwards, R.; Andrews, E.D.; House, H.R. Ecological Processes in a Cold Desert Ecosystem: The Abundance and Species Distribution of Algae Mats in Glacial Meltwater Streams in Taylor Valley, Antarctica. Occasional paper, University of Colorado, Aurora, CO, 1997. Available online: https://instaar.colorado.edu/research/publications/occasional-papers/ecological-processes-in-a-cold-desert-ecosystem-the-abundance-and-species-d/ (accessed on 6 August 2020).
- Ward, N.L.; Challacombe, J.F.; Janssen, P.H.; Henrissat, B.; Coutinho, P.M.; Wu, M.; Xie, G.; Haft, D.H.; Sait, M.; Badger, M.; et al. Three genomes from the phylum Acidobacteria provide insight into the lifestyles of these microorganisms in soils. Appl. Environ. Microbiol. 2009, 75, 2046–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckley, D.; Schmidt, T. The structure of microbial communities in soil and the lasting impact of cultivation. Microb. Ecol. 2001, 42, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Liebner, S.; Harder, J.; Wagner, D. Bacterial diversity and community structure in polygonal tundra soils from Samoylov Island, Lena Delta, Siberia. Int. Microbiol. 2008, 11, 195–202. [Google Scholar] [PubMed]
- Brown, C.T.; Hug, L.A.; Thomas, B.C.; Sharon, I.; Castelle, C.J.; Singh, A.; Wilkins, M.J.; Wrighton, K.C.; Williams, K.H.; Banfield, J.F. Unusual biology across a group comprising more than 15% of domain Bacteria. Nature 2015, 523, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Rinke, C.; Schwientek, P.; Sczyrba, A.; Ivanova, N.N.; Anderson, I.J.; Cheng, J.F.; Darling, A.; Malfatti, S.; Swan, B.K.; Gies, E.A.; et al. Insights into the phylogeny and coding potential of microbial dark matter. Nature 2013, 499, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Buelow, H.N.; Winter, A.S.; Van Horn, D.J.; Barrett, J.E.; Gooseff, M.N.; Schwartz, E.; Takacs-Vesbach, C.D. Microbial Community Responses to Increased Water and Organic Matter in the Arid Soils of the McMurdo Dry Valleys, Antarctica. Front. Microbiol. 2016, 7, 1040. [Google Scholar] [CrossRef] [Green Version]
- Feeser, K.L.; Van Horn, D.J.; Buelow, H.N.; Colman, D.R.; Mchugh, T.A.; Okie, J.G.; Schwartz, E.; Takacs-Vesabach, C.D. Local and regional scale heterogeneity drive bacterial community diversity and composition in a polar desert. Front. Microbiol. 2018, 9, 1928. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, E.; Van Horn, D.J.; Buelow, H.N.; Okie, J.G.; Gooseff, M.N.; Barrett, J.E.; Takacs-Vesbach, C.D. Characterization of growing bacterial populations in McMurdo Dry Valley soils through stable isotope probing with (18) O-water. FEMS Microbiol. Ecol. 2014, 89, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Van Goethem, M.W.; Pierneef, R.; Bezuidt, O.; Van De Peer, Y.; Cowan, D.A.; Makhalanyane, T.P. A reservoir of ‘historical’ antibiotic resistance genes in remote pristine Antarctic soils. Microbiome 2018, 6, 40. [Google Scholar] [CrossRef] [Green Version]
- Koyama, A.; Wallenstein, M.D.; Simpson, R.T.; Moore, J.C. Soil bacterial community composition altered by increased nutrient availability in Arctic tundra soils. Front. Microbiol. 2014, 5, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousk, J.; Baath, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yergeau, E.; Bokhorst, S.; Kang, S.; Zhou, J.; Greer, C.W.; Aerts, R.; Kowalchuk, G.A. Shifts in soil microorganisms in response to warming are consistent across a range of Antarctic environments. ISME J. 2012, 6, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Cameron, R.E. Farthest south algae and associated bacteria. Phycologia 1972, 11, 133–139. [Google Scholar] [CrossRef]
- Wynn-Williams, D. Cyanobacteria in deserts—Life at the limit? In The Ecology of Cyanobacteria; Whitton, B.A., Potts, M., Eds.; Kluwer Academic Publishers: Boca Raton, FL, USA, 2002; pp. 341–366. [Google Scholar]
- Engelskjøn, T. Botany of two Antarctic mountain ranges: Gjelsvikfjella and Muhlig-Hofmannfjella, Dronning Maud Land. Polar Res. 1986, 4, 205–224. [Google Scholar] [CrossRef] [Green Version]
- Stomeo, F.; Makhalanyane, T.P.; Valverde, A.; Pointing, S.; Stevens, M.; Cary, C.; Tuffin, M.; Cowan, D. Abiotic factors influence microbial diversity in permanently cold soil horizons of a maritime-associated Antarctic Dry Valley. FEMS Microbiol. Ecol. 2012, 82, 326–340. [Google Scholar] [CrossRef]
- Newsham, K.K.; Pearce, D.A.; Bridge, P. Minimal influence of water and nutrient content on the bacterial community composition of a maritime Antarctic soil. Microbiol. Res. 2010, 165, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Tin, T.; Fleming, Z.L.; Hughes, K.A.; Ainley, D.G.; Convey, P.; Moreno, C.A.; Pfeiffer, S.; Scott, J.; Snape, I. Impacts of local human activities on the Antarctic environment. Antarct. Sci. 2009, 21, 3–33. [Google Scholar] [CrossRef] [Green Version]
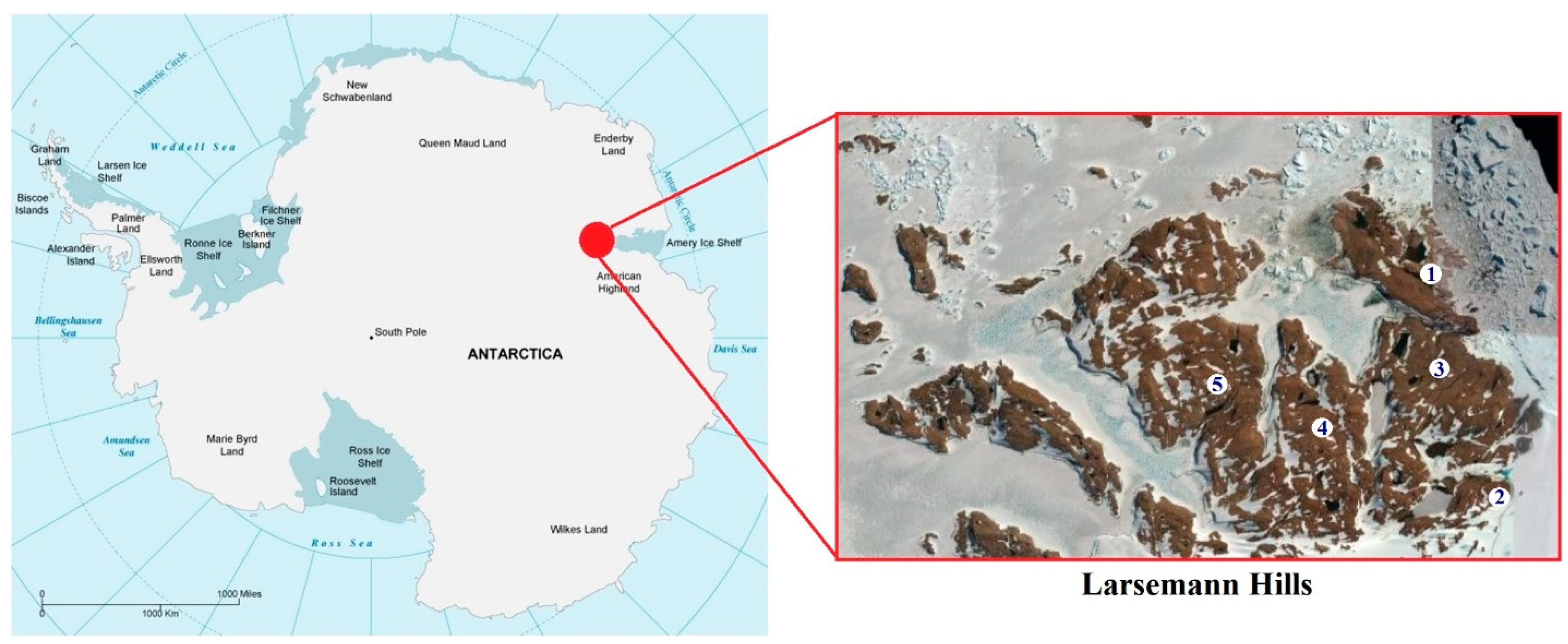





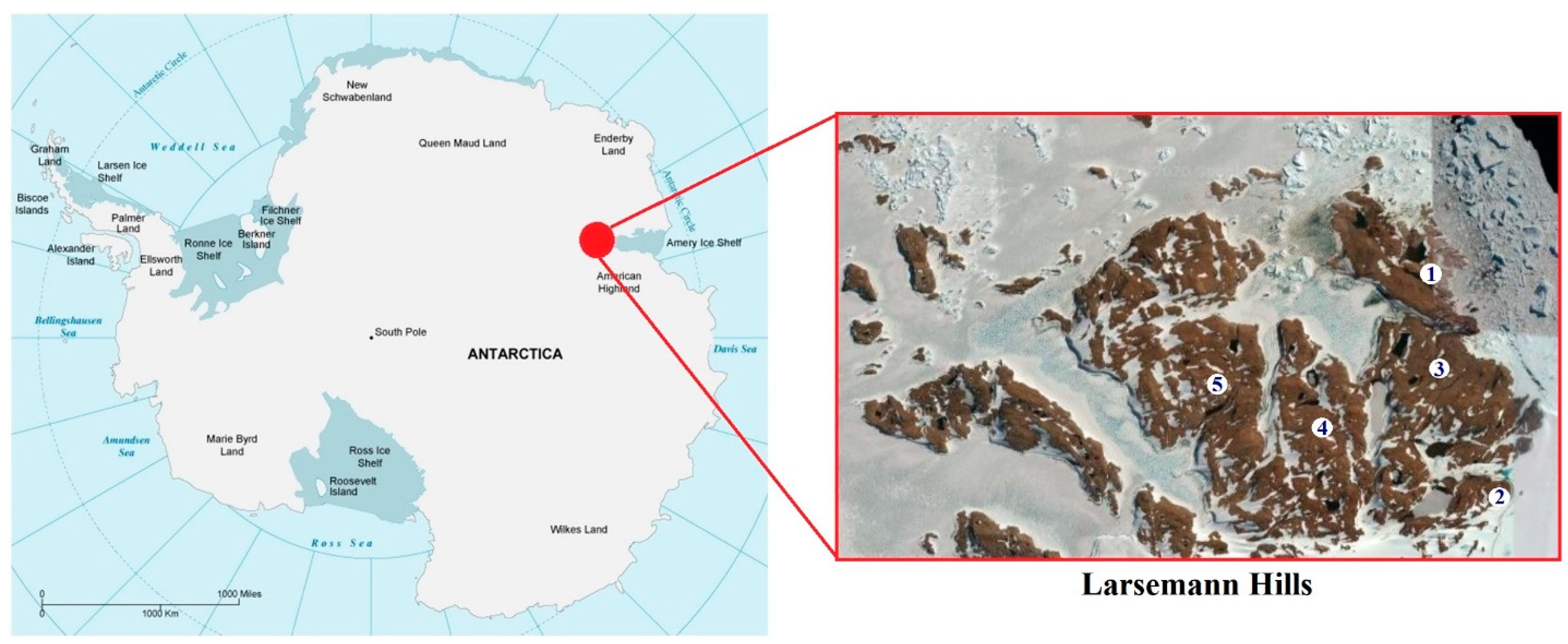{kind=link}
{kind=link}
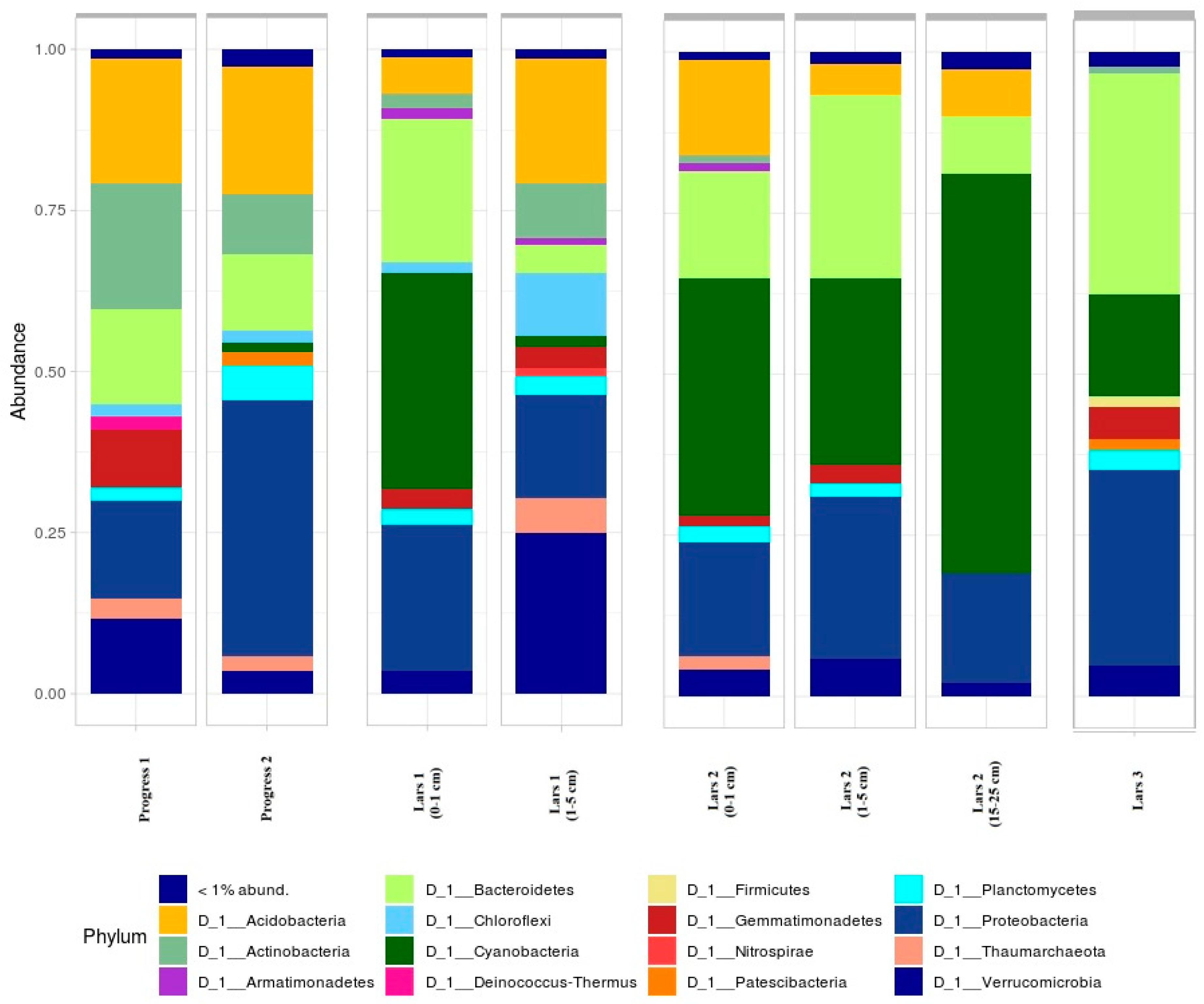{kind=link}
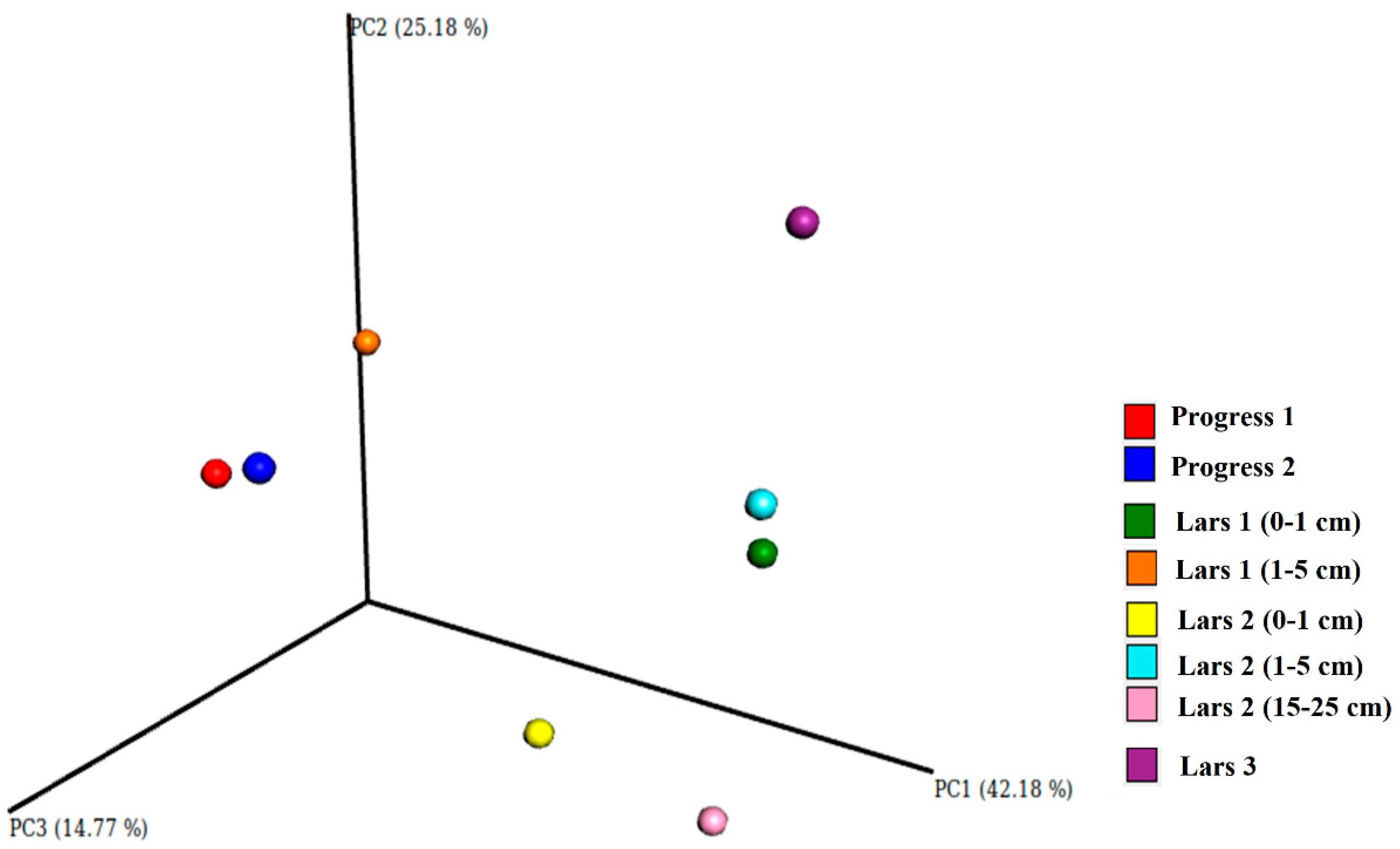{kind=link}
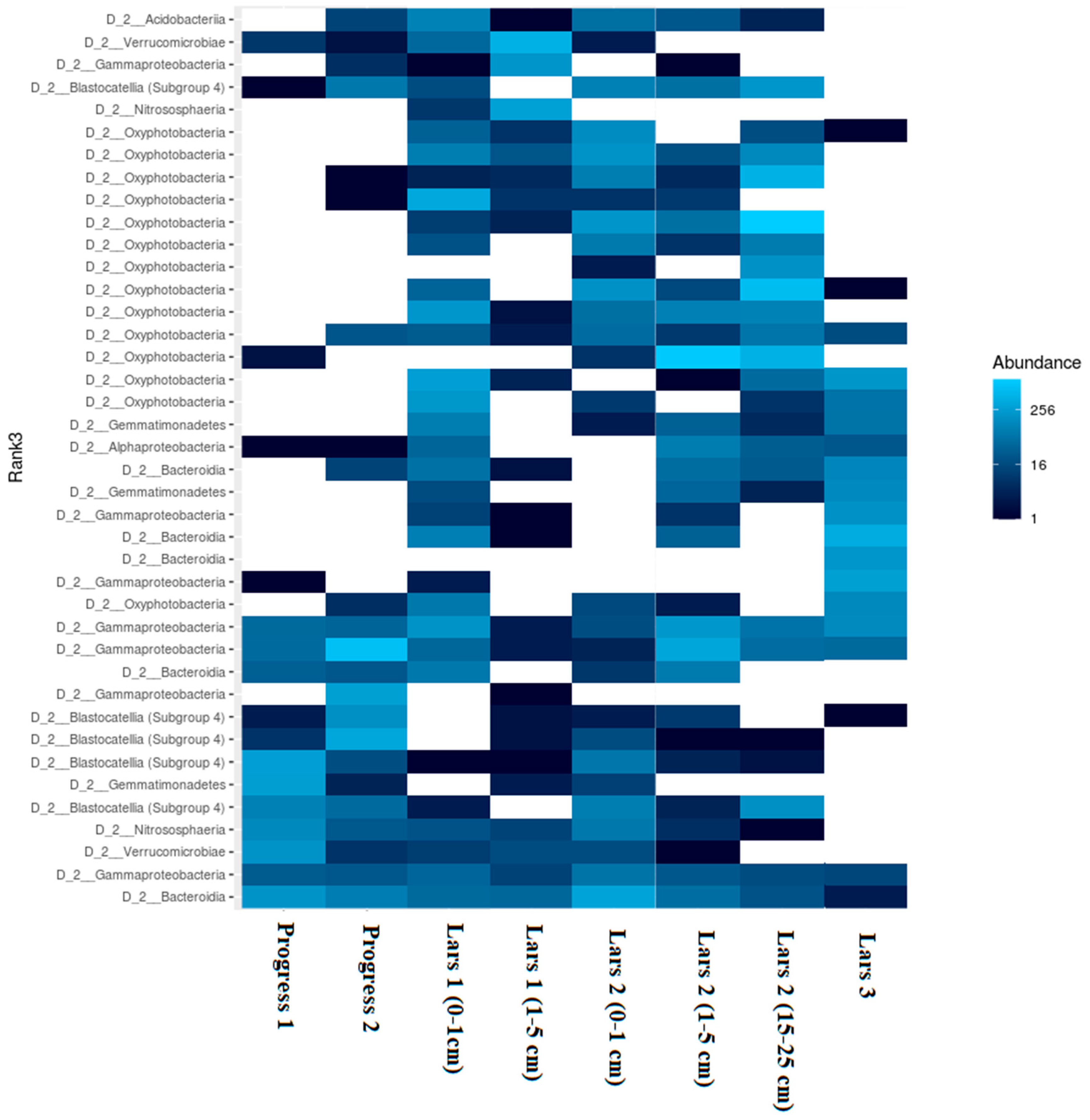{kind=link}
| Coordinates | MAAT, °C | MAST, °C | MAGT, °C | Mean Annual Wind Speed, m s−1 | Annual Precipitation, mm |
|---|---|---|---|---|---|
| Larsemann Hills | |||||
| S 69°40′23.2″ E 76°34′21.2″ | −9.6 | −10.4 | −9.1 | 6.7 | 250 |
| Site | Geographical Coordinates | Location and Horizon Description | Soil Name (WRB) |
|---|---|---|---|
| Anthropogenically-Affected Sites | |||
| Progress 1 | 69°22′39.7″S 76°23′15.9″E | Area between office and residential buildings; surface horizon | Hyperskeletic Turbic Technosol |
| Progress 2 | 69°24′02.6″S 76°24′05.2″E | Vicinities of field base “Progress-3”, wet valley, surface horizon | Hyperskeletic Lithic Leptosol |
| Near-Natural Sites | |||
| Lars 1 (0–1 cm) | 69°23′11.4″S 76°23′16.7″E | Surface “algal–mineral” horizon, wet valley of Larsemann Hills | Hyperskeletic Leptosol (Lithic) |
| Lars 1 (1–5 cm) | Moss–lichen–algae horizon in bottom of the wet valley of Larsemann Hills | ||
| Lars 2 (0–1 cm) | 69°23′45.1″S 76°21’08.7″E | Surface coarse horizon under “gravel pavement” in flank of the valley with ephemeral water supply from melting snow patches | Hyperskeletic Leptosol (Lithic, Gleyic) |
| Lars 2 (1–5 cm) | “Algal–mineral” mineral horizon | ||
| Lars 2 (15–25 cm) | Transitional (BC) horizon, with stagnic and oximorphic features | ||
| Lars 3 | 69°23′12.9″S 76°19′26.6″E | Gravel pavement in high “dry” flank of the valley | Hyperskeletic Leptosol (Lithic) |
| Site | pHH2O | pHKCl | TOC, % | N, % | C:N | Basal Respiration, mg g−1 day−1 | Fine Earth (<2 mm), % |
|---|---|---|---|---|---|---|---|
| Anthropogenically-Affected Sites | |||||||
| Progress 1 | 5.85 ± 0.53 | 5.23 ± 0.42 | 0.87 ± 0.10 | 0.083± 0.005 | 10.48 ± 1.27 | 0.009 ± 0.001 | 18.99 ± 1.23 |
| Progress 2 | 5.32 ± 0.56 | 4.89± 0.32 | 0.76 ±0.16 | 0.062 ±0.005 | 12.26 ± 2.09 | 0.008 ± 0.001 | 21.91 ± 1.97 |
| Near-Natural Sites | |||||||
| Lars 1 (0–1 cm) | 6.42 ± 0.56 | 5.76 ± 0.45 | 0.89 ± 0.12 | 0.08 ± 0.010 | 11.13 ± 1.56 | 0.010 ± 0.002 | 21.78 ± 1.86 |
| Lars 1 (1–5 cm) | 6.01 ±0.43 | 5.54 ± 0.31 | 1.50 ± 0.12 | 0.12± 0.01 | 12.50 ± 1.23 | 0.101 ± 0.006 | 31.98 ± 1.98 |
| Lars 2 (0–1 cm) | 6.21 ± 0.42 | 5.86 ± 0.23 | 0.98 ± 0.12 | 0.06 ± 0.01 | 16.33 ± 1.86 | 0.009 ± 0.001 | 22.03 ± 1.54 |
| Lars 2 (1–5 cm) | 5.98 ± 0.32 | 5.54 ± 0.27 | 1.65 ± 0.17 | 0.15 ± 0.03 | 10.84 ± 0.87 | 0.150 ± 0.010 | 29.87 ± 1.50 |
| Lars 2 (15–25 cm) | 6.03 ± 0.24 | 5.65 ± 0.23 | 0.54 ± 0.08 | 0.04 ± 0.01 | 13.50 ± 1.24 | 0.006 ± 0.001 | 19.87 ± 1.65 |
| Lars 3 | 6.04 ± 0.21 | 5.76 ± 0.21 | 0.87 ± 0.10 | 0.08 ± 0.02 | 10.88 ± 1.26 | 0.007 ± 0.001 | 21.09 ± 1.78 |
| Site | OTUs | PD_Whole_Tree | Simpson Index | Shannon Index |
|---|---|---|---|---|
| Anthropogenically-Affected Sites | ||||
| Progress 1 | 504 ± 32 | 40 ± 3.5 | 0.985 ± 0.12 | 5.02 ± 0.78 |
| Progress 2 | 760 ± 28 | 58 ± 4.1 | 0.981 ± 0.15 | 5.31 ± 0.85 |
| Near-Natural Sites | ||||
| Lars 1 (0–1 cm) | 566 ± 26 | 45 ± 3.5 | 0.984 ± 0.17 | 5.10 ± 1.02 |
| Lars 1 (1–5 cm) | 513 ± 25 | 46 ± 5.2 | 0.967 ± 0.11 | 4.85 ± 0.65 |
| Lars 2 (0–1 cm) | 476 ± 22 | 40 ± 4.2 | 0.981 ± 0.14 | 4.85 ± 0.98 |
| Lars 2 (1–5 cm) | 590 ± 29 | 46 ± 6.1 | 0.959 ± 0.15 | 4.75 ± 0.76 |
| Lars 2 (15–25 cm) | 310 ± 21 | 28 ± 5.3 | 0.945 ± 0.23 | 4.02 ± 0.73 |
| Lars 3 | 403 ± 22 | 41.5 ± 4.5 | 0.979 ± 0.25 | 4.63 ± 0.56 |
| pH | TOC | N | C/N | Basal Respiration | Fine Earth | OTUs | Shannon | Simpson | PD_Tree | |
|---|---|---|---|---|---|---|---|---|---|---|
| pH | — | |||||||||
| TOC | 0.273 | — | ||||||||
| N | −0.034 | 0.758 *** | — | |||||||
| C/N | 0.285 | −0.093 | −0.588 ** | — | ||||||
| Basal Respiration | 0.176 | 0.876 *** | 0.806 *** | −0.242 | — | |||||
| Fine Earth | 0.148 | 0.757 *** | 0.522 ** | 0.258 | 0.723 *** | — | ||||
| OTUs | −0.244 | 0.245 | 0.436 * | −0.312 | 0.555 ** | 0.397 | — | |||
| Shannon | −0.056 | 0.032 | 0.156 | −0.101 | 0.283 | 0.163 | 0.795 *** | — | ||
| Simpson | 0.422 * | 0.263 | 0.267 | 0.052 | 0.271 | 0.207 | 0.368 | 0.688 *** | — | |
| PD_tree | −0.167 | 0.358 * | 0.509 * | −0.094 | 0.552 ** | 0.691 *** | 0.848 *** | 0.677 *** | 0.391 | — |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alekseev, I.; Zverev, A.; Abakumov, E. Microbial Communities in Permafrost Soils of Larsemann Hills, Eastern Antarctica: Environmental Controls and Effect of Human Impact. Microorganisms 2020, 8, 1202. https://doi.org/10.3390/microorganisms8081202
Alekseev I, Zverev A, Abakumov E. Microbial Communities in Permafrost Soils of Larsemann Hills, Eastern Antarctica: Environmental Controls and Effect of Human Impact. Microorganisms. 2020; 8(8):1202. https://doi.org/10.3390/microorganisms8081202
Chicago/Turabian StyleAlekseev, Ivan, Aleksei Zverev, and Evgeny Abakumov. 2020. "Microbial Communities in Permafrost Soils of Larsemann Hills, Eastern Antarctica: Environmental Controls and Effect of Human Impact" Microorganisms 8, no. 8: 1202. https://doi.org/10.3390/microorganisms8081202
APA StyleAlekseev, I., Zverev, A., & Abakumov, E. (2020). Microbial Communities in Permafrost Soils of Larsemann Hills, Eastern Antarctica: Environmental Controls and Effect of Human Impact. Microorganisms, 8(8), 1202. https://doi.org/10.3390/microorganisms8081202