Moving from qPCR to Chip Digital PCR Assays for Tracking of some Fusarium Species Causing Fusarium Head Blight in Cereals

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Samples
2.2. Plant Samples
2.3. Design of Primers and Probes
2.4. qPCR
2.5. Chip Digital PCR
3. Results
3.1. Fungal Samples
3.2. Plant Samples
- Naturally contaminated samples, belonging to classes ii) and iii);
- Synthetic samples created by spiking plant DNA (extracted from samples found not contaminated) with fungal DNA at different concentrations.
4. Discussion
- ➢
- As the most advantageous feature, dPCR relies on absolute quantification of the target operated by dPCR; on the contrary, “results generated from qPCR were relative to calibration curve and were not the actual number of copies in a sample itself” [23].
- ➢
- Secondarily, the high sample partitioning ensures accurate results even at very low target copy numbers as well as detection of rare targets even in a high background of non-target DNA [24],
- ➢
- Lastly, dPCR is less sensitive to contaminants eventually present in the samples; complex biomolecules such as humic acid can, in fact, significantly inhibit qPCR reactions, but dPCR can overcome this lack thanks to its endpoint quantification [23].
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bertero, A.; Moretti, A.; Spicer, L.J.; Caloni, F. Fusarium molds and mycotoxins: Potential species-specific effects. Toxins 2018, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- Schaarschmidt, S.; Fauhl-Hassek, C. The fate of mycotoxins during the processing of wheat for human consumption. Compr. Rev. Food Sci. Food Saf. 2018, 17, 556–593. [Google Scholar] [CrossRef] [Green Version]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegulo, S.N. Factors influencing deoxynivalenol accumulation in small grain cereals. Toxins 2012, 4, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.G.; Godley, N.P. Reduction of Fusarium head blight and deoxynivalenol in wheat with early fungicide applications of prothioconazole. FAD Part. A 2010, 27, 629–635. [Google Scholar]
- Prandini, A.; Sigolo, S.; Filippi, L.; Battilani, P.; Piva, G. Review of predictive models for Fusarium head blight and related mycotoxin contamination in wheat. Food Chem Toxicol. 2009, 47, 27–931. [Google Scholar] [CrossRef]
- Morcia, C.; Rattotti, E.; Stanca, A.M.; Tumino, G.; Rossi, V.; Ravaglia, S.; Germeier, C.U.; Herrmann, M.; Polisenska, I.; Terzi, V. Fusarium genetic traceability: Role for mycotoxin control in small grain cereals agro-food chains. J. Cereal Sci. 2013, 57, 175–182. [Google Scholar] [CrossRef]
- Mulè, G.; González-Jaén, M.T.; Hornok, L.; Nicholson, P.; Waalwijk, C. Advances in molecular diagnosis of toxigenic Fusarium species: A review. FAD 2005, 22, 316–323. [Google Scholar]
- Ceresoli, C. Metodiche Molecolari Per L’identificazione e Quantificazione di Funghi Produttori di Micotossine in Cereali. Master’s Thesis, University of Modena and Reggio Emilia, Reggio Emilia, Italy, July 2019. [Google Scholar]
- Pecoraro, S.; Berben, G.; Burns, M.; Corbisier, P.; De Giacomo, M.; De Loose, M.; Dagand, E.; Dobnik, D.; Eriksson, R.; Holst-Jensen, A. Overview and Recommendations for the Application of Digital PCR. EUR 29673 EN; Publications Office of the European Union: Luxembourg, 2019; JRC115736; ISBN 978-92-76-00180-5. [Google Scholar] [CrossRef]
- Taylor, S.C.; Laperriere, G.; Germain, H. Droplet digital PCR versus qPCR for gene expression analysis with low abundant targets: From variable nonsense to publication quality data. Sci. Rep. 2017, 7, 2409. [Google Scholar] [CrossRef] [Green Version]
- Quan, P.-L.; Sauzade, M.; Brouzes, E. dPCR: A technology review. Sensors 2018, 18, 1271. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Aguirre, I.; Rački, N.; Dreo, T.; Ravnikar, M. Droplet digital PCR for absolute quantification of pathogens. In Plant. Pathology. Techniques and Protocols, 2nd ed.; Lacomme, C., Ed.; Humana Press: New York, NY, USA, 2015; Volume 1302, pp. 331–347. [Google Scholar]
- Liu, J.; Li, C.; Muhae-Ud-Din, G.; Liu, T.; Chen, W.; Zhang, J.; Gao, L. Development of the droplet digital PCR to detect the teliospores of Tilletia controversa Kühn in the soil with greatly enhanced sensitivity. Front. Microbiol. 2020, 11, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santander, R.D.; Meredith, C.L.; Aćimović, S.G. Development of a viability digital PCR protocol for the selective detection and quantification of live Erwinia amylovora cells in cankers. Sci. Rep. 2019, 9, 11530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Samarrai, T.H.; Schmid, J. A simple method for extraction of fungal genomic DNA. Lett. Appl. Microbiol. 2000, 30, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Kulik, T.; Jestoi, M.; Okorski, A. Development of TaqMan assays for the quantitative detection of Fusarium avenaceum/Fusarium tricinctum and Fusarium poae esyn1 genotypes from cereal grain. FEMS Microbiol. Lett. 2011, 314, 49–56. [Google Scholar] [CrossRef]
- Morcia, C.; Bergami, R.; Scaramagli, S.; Ghizzoni, R.; Carnevali, P.; Terzi, V. A chip digital PCR assay for quantification of common wheat contamination in pasta production chain. Foods 2020, 9, 911. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, M.; Beyer, M.; Logrieco, A.; Audenaert, K.; Balmas, V.; Basler, R.; Boutigny, A.L.; Chrpová, J.; Czembor, E.; Gagkaeva, T.; et al. A European database of Fusarium graminearum and F. culmorum trichothecene genotypes. Front. Microbiol. 2016, 7, 406. [Google Scholar]
- Vanheule, A.; De Boevre, M.; Moretti, A.; Scauflaire, J.; Munaut, F.; De Saeger, S.; Bekaert, B.; Haesaert, G.; Waalwijk, C.; Van der Lee, T. Genetic divergence and chemotype diversity in the fusarium head blight pathogen Fusarium poae. Toxins 2017, 9, 255. [Google Scholar] [CrossRef]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in cereal grains – An update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [Green Version]
- Jestoi, M. Emerging Fusarium -mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin—A review. Critical Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef]
- Cao, Y.; Yu, M.; Dong, G.; Chen, B.; Zhang, B. Digital PCR as an emerging tool for monitoring of microbial biodegradation. Molecules 2020, 25, 706. [Google Scholar] [CrossRef] [Green Version]
- Demeke, T.; Dobnik, D. Critical assessment of digital PCR for the detection and quantification of genetically modified organisms. Anal. Bioanal Chem 2018, 410, 4039–4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehle, N.; Gregur, L.; Bogožalec Košir, A.; Dobnik, D. One-step reverse-transcription digital PCR for reliable quantification of different Pepino mosaic virus genotypes. Plants 2020, 9, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay Code | Probe and Primers | Biological Target | Target Gene | Reference |
|---|---|---|---|---|
| F. spo | Pr: FAM-CTGCATCACAACCC-MGB F: GCAAGTCGACCACTGTGAGTACA R: TGAAACTACCCCGCCAAGTC | F. sporotrichioides | tef1 GenBank: MN120771.1 | This work |
| F. gram/culm | Pr: FAM-ATCAGTGCTTAAATGCA-MGB F: CAGTAGAGTCGACAAGATCTGCAATC R: TGAAAGTCGCGTAGCTGGAA | F. graminearum F. culmorum | Tri GenBank: MH514957.1 | This work |
| F. poae | Pr: FAM-AAAGCGGTCGAGTCTG-MGB F: GCGGCCGCTTTTGTCA R: GCCTTTCCAGCAAGAGATGGT | F. poae | esyn1 | [17] |
| F. avetric | Pr: FAM- CCGTCGAGTCCTCT -MGB F: AGCAGTCGAGTTCGTCAACAGA R: GGCYTTTCCTGCGAACTTG | F. avenaceum, F.tricinctum | esyn1 | [17] |
| Grano CO2 | Pr: VIC- CATGAGCGTGTGCGTG -MGB F: TGCTAACCGTGTGGCATCAC R: GGTACATAGTGCTGCTGCATCTG | Triticum genus | Triticum TaHd1 | [18] |
| Avena dig | Pr: VIC- ACAATCTTTGCTTGTTCTT-MGB F: TCGTTGATTTTTGGTTGCTTTG R: AGCCTTTGCAATCCACATCTG | Avena | actin 1, GenBank: AF234528.1 | This work |
| Assay Code | Initial Activation Step | Denaturation/Annealing/Amplification Step | N. of Cycles |
|---|---|---|---|
| F. sporo | 95 °C, 10 min | 95 °C, 30 s; 58 °C, 2 min | 47 |
| F. gram/culm | 95 °C, 10 min | 95 °C, 30 s; 60 °C, 2 min | 45 |
| F. poae | 95 °C, 10 min | 95 °C, 30 s; 60 °C, 2 min | 45 |
| F. avetric | 95 °C, 10 min | 95 °C, 30 s; 59 °C, 2 min | 47 |
| Grano CO2 | 96 °C, 10 min | 98 °C, 30 s; 58–60 °C, 2 min | 45–47 |
| Avenadig | 96 °C, 10 min | 95 °C, 30 s; 58–60 °C, 2 min | 45–47 |
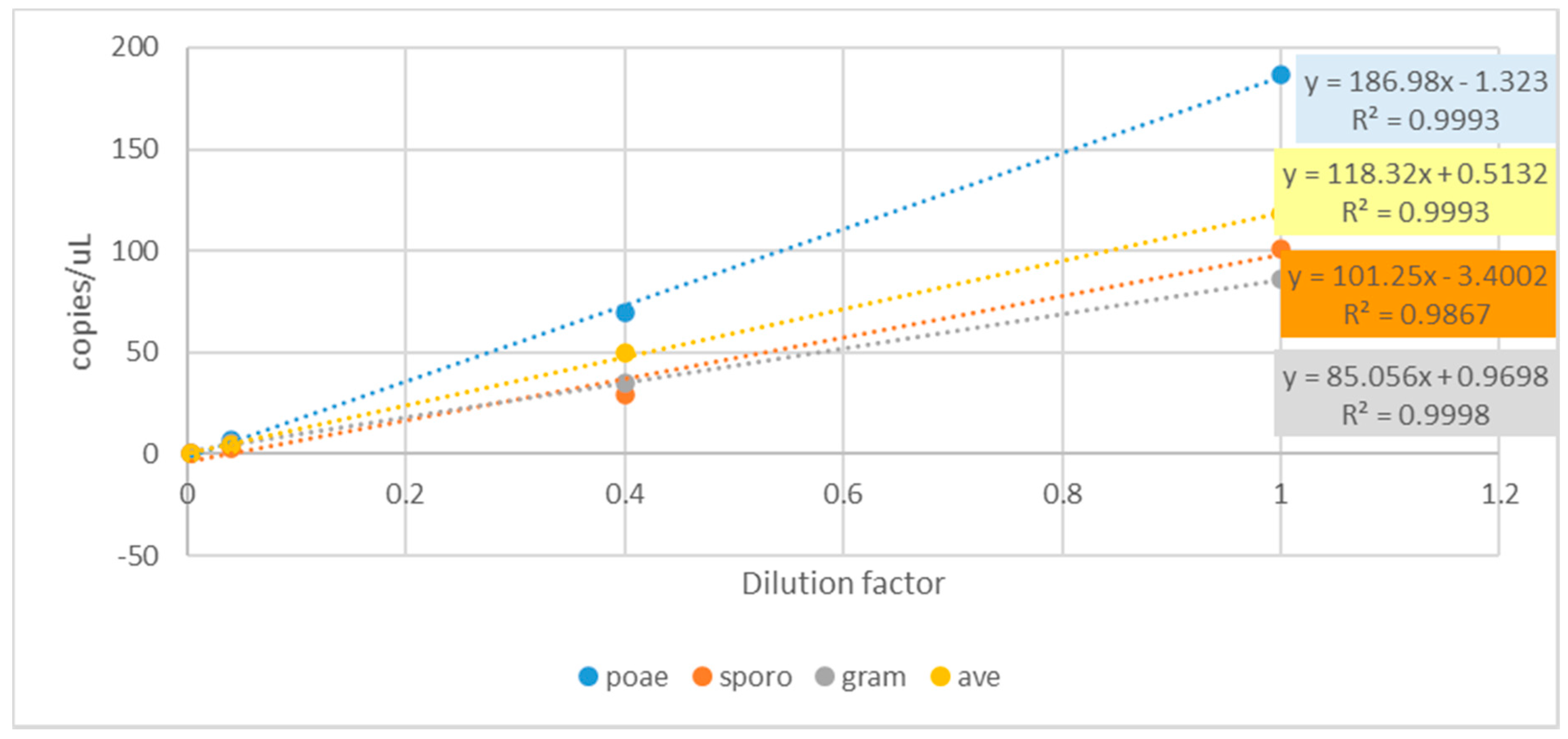
| qPCR Assay Name | R2 Coefficient | Amplification Efficiency |
|---|---|---|
| F. sporo | 0.984 | 104% |
| F. gram/culm | 0.969 | 124% |
| F. poae | 0.997 | 100% |
| F. avetric | 0.991 | 106% |
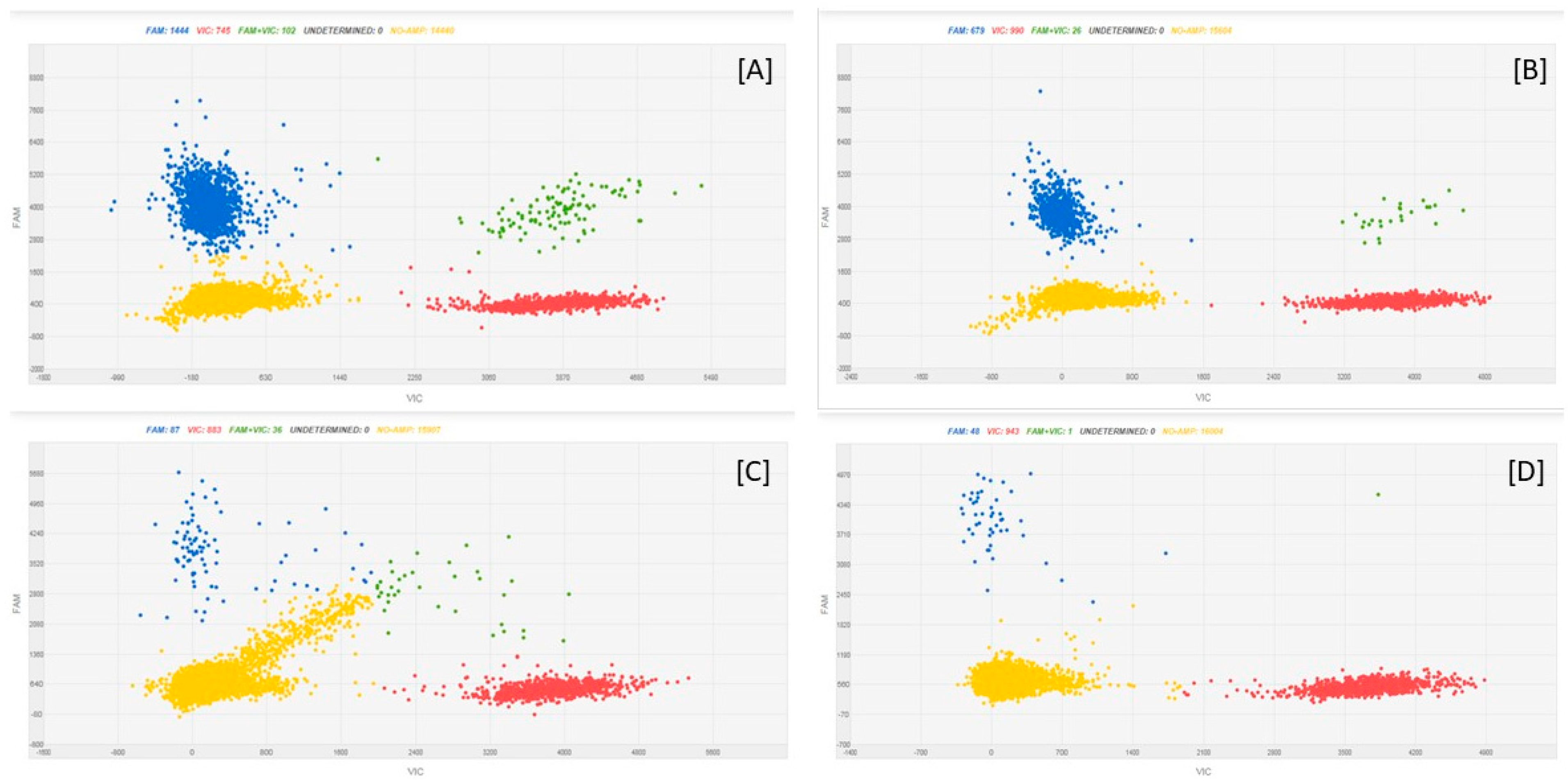
| Target | Limit of Detection | Sensitivity |
|---|---|---|
| F. sporotrichioides | 10 | 0.466% |
| F. graminearum | 13 | 0.6% |
| F. poae | 2 | 0.134% |
| F. avenaceum | 8 | 0.636% |
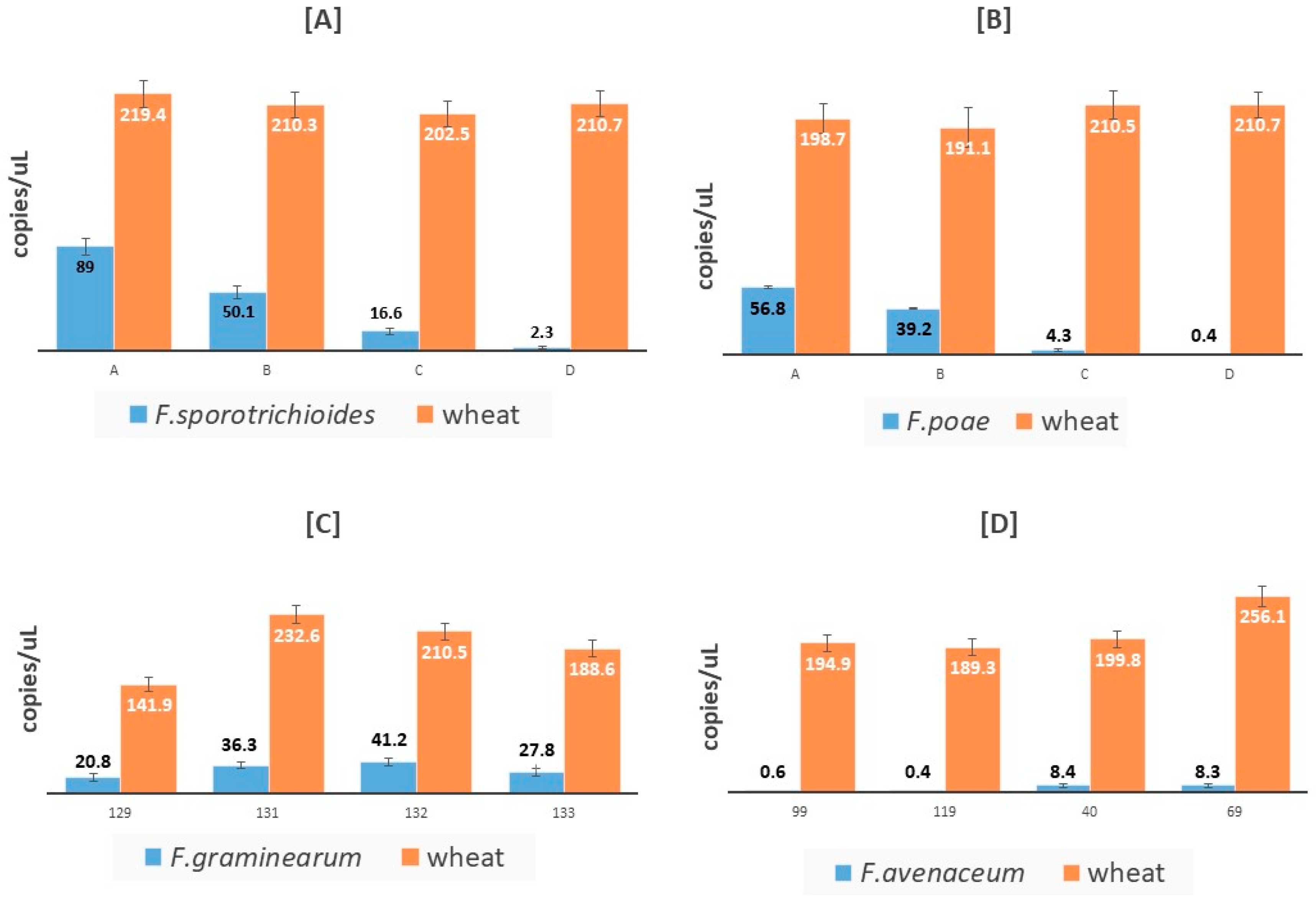
| Fusarium Strain | Plant Genus | Naturally Contaminated Sample | Spiked Sample | pg of Fungal DNA/50 ng plant DNA (qPCR) | Fungal copies/μL (cdPCR) |
|---|---|---|---|---|---|
| F. sporotrichioides | Triticum durum | X | 360 ± 12 | 89 ± 7.2 | |
| F. sporotrichioides | Triticum durum | X | 120 ± 9 | 50.1 ± 5.2 | |
| F. sporotrichioides | Triticum durum | X | 61 ± 3 | 16.65 ± 3 | |
| F. sporotrichioides | Triticum durum | X | 4.1 ± 0.9 | 2.35 ± 1.1 | |
| F. graminearum/culmorum | Triticum durum | X | 34 ± 5 | 20.8 ± 3.4 | |
| F. graminearum/culmorum | Triticum durum | X | 60 ± 2 | 36.35 ± 4.7 | |
| F. graminearum/culmorum | Triticum durum | X | 66 ± 9 | 41.2 ± 4.9 | |
| F. graminearum/culmorum | Triticum durum | X | 57 ± 6 | 27.8 ± 4.2 | |
| F. poae | Triticum durum | X | 1.3 ± 0.2 | 0.08 ± 0.04 | |
| F. poae | Triticum durum | X | 2.1 ± 0.5 | 0.87 ± 0.1 | |
| F. poae | Triticum durum | X | 1.6 ± 0.6 | 0.91 ± 0.5 | |
| F. poae | Triticum durum | X | 126 ± 1.5 | 56.7 ± 6.4 | |
| F. poae | Triticum durum | X | 62 ± 0.9 | 39.2 ± 7.3 | |
| F. poae | Triticum durum | X | 6.2 ± 0.4 | 4.35 ± 1.6 | |
| F. poae | Triticum durum | X | 0.15 ± 0.1 | 0.40 ± 0.2 | |
| F. avenaceum | Triticum durum | X | 1.1 ± 2.1 | 0.6 ± 0.46 | |
| F. avenaceum | Triticum durum | X | 1.6 ± 0.9 | 0.4 ± 0.3 | |
| F. avenaceum | Triticum durum | X | 14 ± 2.3 | 8.4 ± 2.2 | |
| F. avenaceum | Triticum durum | X | 16 ± 1.9 | 8.3 ± 2.3 | |
| F. avenaceum | Triticum durum | X | 300 ± 9.2 | 118 ± 8.5 | |
| F. avenaceum | Triticum durum | X | 120 ± 7.5 | 50.1 ± 5.4 | |
| F. avenaceum | Triticum durum | X | 46 ± 4.6 | 5.5 ± 1.8 | |
| F. avenaceum | Triticum durum | X | 4.8 ± 1.8 | 0.89 ± 0.71 | |
| F. avenaceum | Avena sativa | X | 286 ± 8.2 | 128 ± 9 | |
| F. avenaceum | Avena sativa | X | 111 ± 5.8 | 55.3 ± 5.8 | |
| F. avenaceum | Avena sativa | X | 42 ± 4.2 | 10.1 ± 2.4 | |
| F. avenaceum | Avena sativa | X | 5.9 ± 1.2 | 3.9 ± 1.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morcia, C.; Tumino, G.; Gasparo, G.; Ceresoli, C.; Fattorini, C.; Ghizzoni, R.; Carnevali, P.; Terzi, V. Moving from qPCR to Chip Digital PCR Assays for Tracking of some Fusarium Species Causing Fusarium Head Blight in Cereals. Microorganisms 2020, 8, 1307. https://doi.org/10.3390/microorganisms8091307
Morcia C, Tumino G, Gasparo G, Ceresoli C, Fattorini C, Ghizzoni R, Carnevali P, Terzi V. Moving from qPCR to Chip Digital PCR Assays for Tracking of some Fusarium Species Causing Fusarium Head Blight in Cereals. Microorganisms. 2020; 8(9):1307. https://doi.org/10.3390/microorganisms8091307
Chicago/Turabian StyleMorcia, Caterina, Giorgio Tumino, Giulia Gasparo, Caterina Ceresoli, Chiara Fattorini, Roberta Ghizzoni, Paola Carnevali, and Valeria Terzi. 2020. "Moving from qPCR to Chip Digital PCR Assays for Tracking of some Fusarium Species Causing Fusarium Head Blight in Cereals" Microorganisms 8, no. 9: 1307. https://doi.org/10.3390/microorganisms8091307
APA StyleMorcia, C., Tumino, G., Gasparo, G., Ceresoli, C., Fattorini, C., Ghizzoni, R., Carnevali, P., & Terzi, V. (2020). Moving from qPCR to Chip Digital PCR Assays for Tracking of some Fusarium Species Causing Fusarium Head Blight in Cereals. Microorganisms, 8(9), 1307. https://doi.org/10.3390/microorganisms8091307