Differential Bacterial Predation by Free-Living Amoebae May Result in Blooms of Legionella in Drinking Water Systems


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. L. pneumophila Culture
2.2. E. coli Culture
2.3. Amoebae Culture
2.4. Amoeba–Bacteria Co-Culture
2.5. Fluorescent Microscopy and Image Processing
2.6. Determining the Intracellular Bacteria
2.7. Statistical Analysis
3. Results
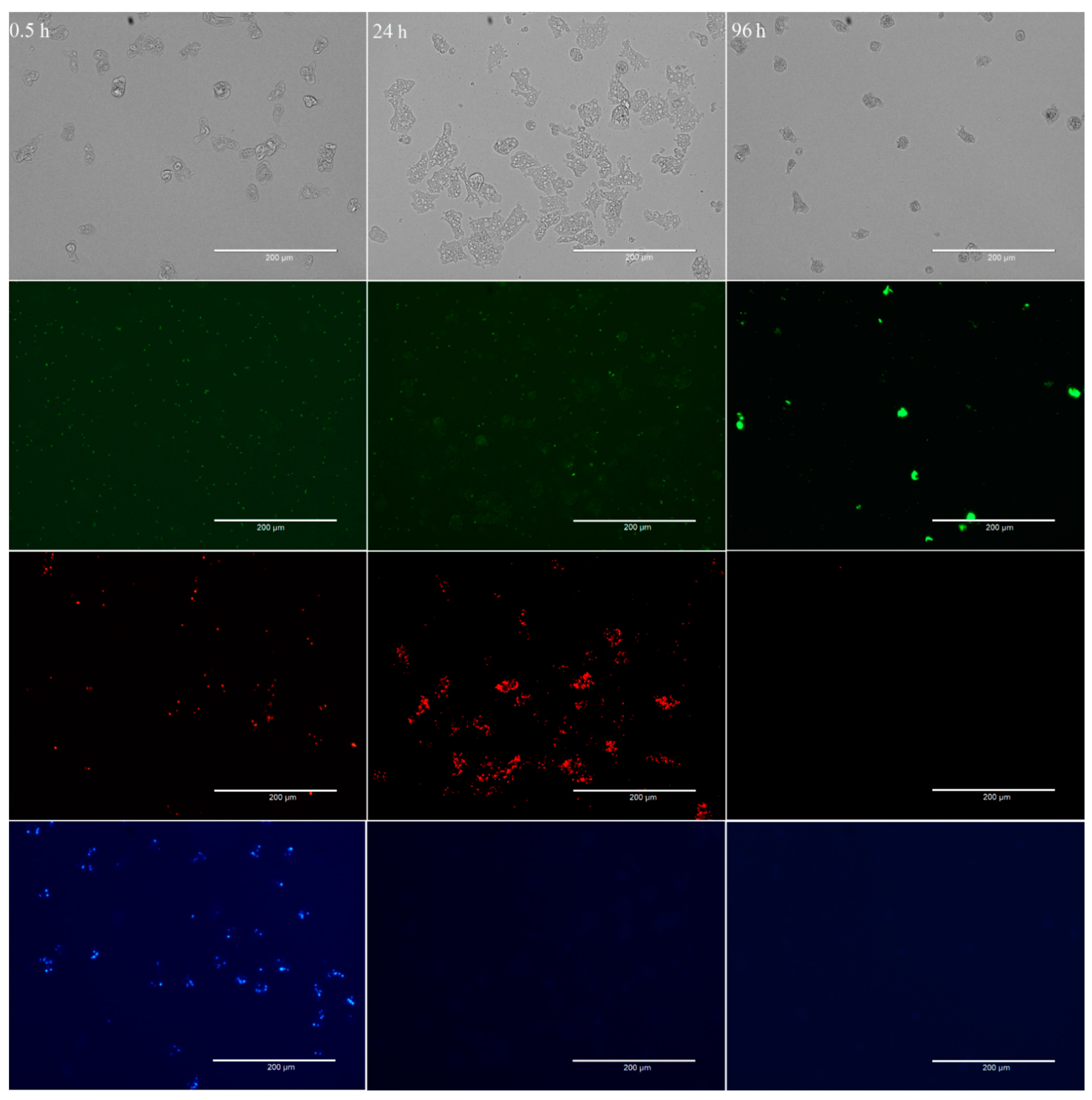
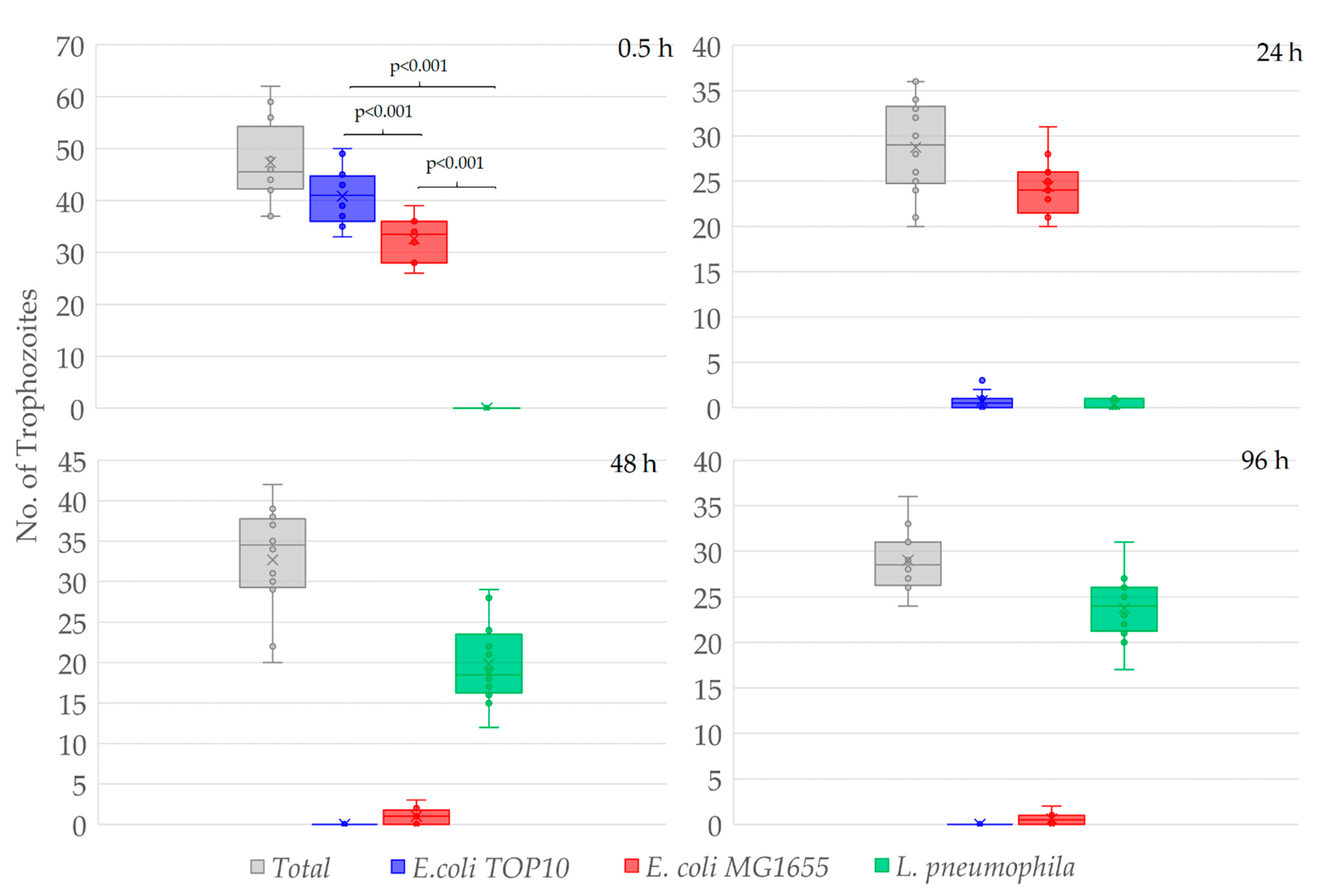
3.1. Differential Feeding Preference of W. magna
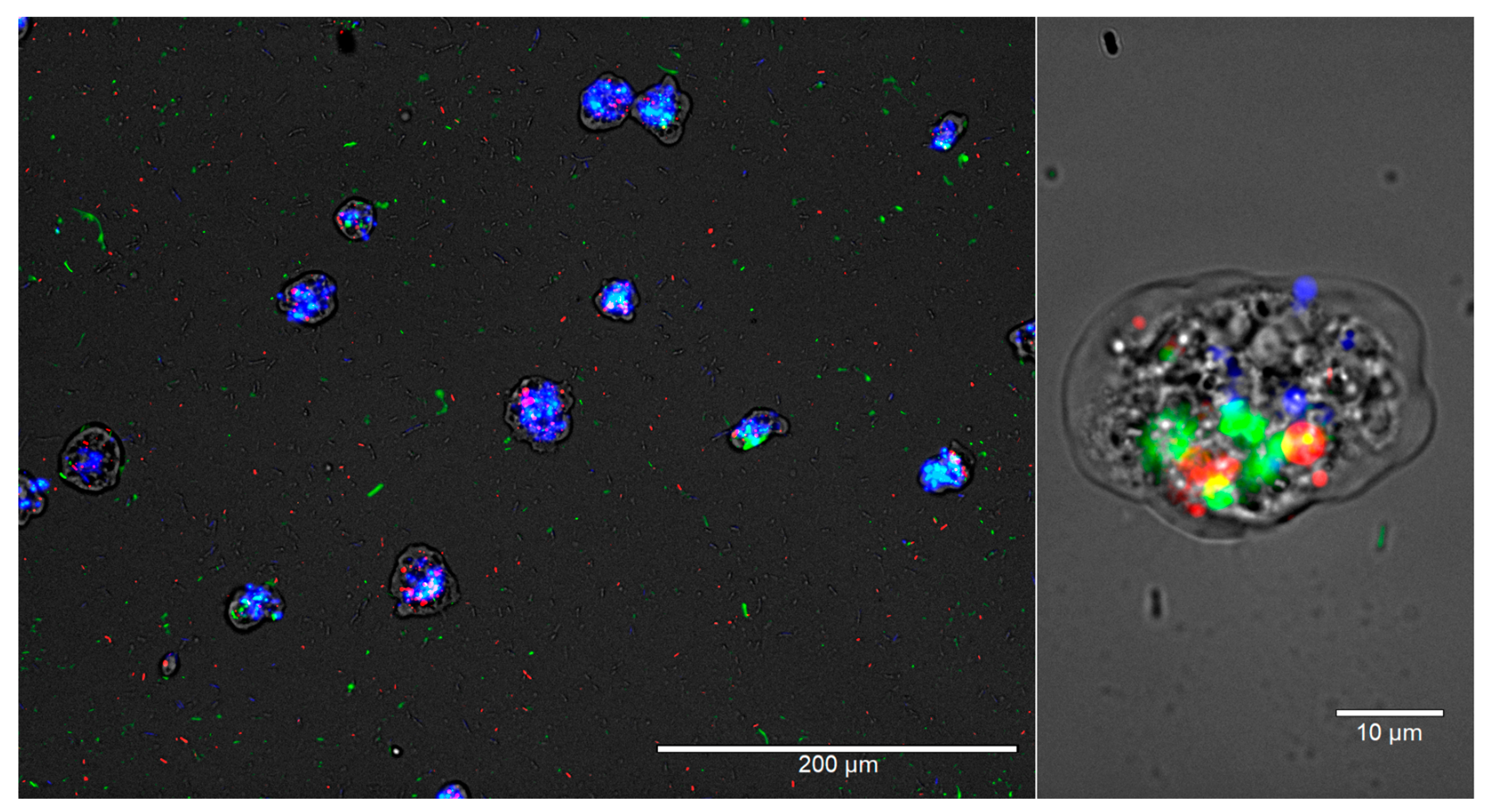
3.2. Interactions of A. polyphaga and V. vermiformis with Bacteria
3.3. Amoebae–Bacteria Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atlas, R. Legionella: From environmental habitats to disease pathology, detection and control. Environ. Microbiol. 1999, 1, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Orrison, L.H.; Cherry, W.B.; Fliermans, C.B.; Dees, S.B.; McDougal, L.K.; Dodd, D.J. Characteristics of environmental isolates of Legionella pneumophila. Appl. Environ. Microbiol. 1981, 42, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaute, J.; Zucs, P.; de Jong, B. On behalf of the European Legionnaires’ Disease Surveillance Network (2013) Legionnaires’ disease in Europe, 2009–2010. Euro Surveill. 2013, 18, 6–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beer, K.D.; Gargano, J.W.; Roberts, V.A.; Hill, V.R.; Garrison, L.E.; Kutty, P.K.; Hilborn, E.D.; Wade, T.J.; Fullerton, K.E.; Yoder, J.S. Surveillance for Waterborne Disease Outbreaks Associated with Drinking Water—United States, 2011–2012. Morb. Mortal. Wkly. Rep. 2015, 64, 842–848. [Google Scholar] [CrossRef]
- Gargano, J.; Adam, E.; Collier, S.; Fullerton, K.; Feinman, S.; Beach, M. Mortality from selected diseases that can be transmitted by water–United States, 2003–2009. J. Water Health 2017, 15, 438–450. [Google Scholar] [CrossRef] [Green Version]
- Craun, G.F.; Brunkard, J.M.; Yoder, J.S.; Roberts, V.A.; Carpenter, J.; Wade, T.; Calderon, R.L.; Roberts, J.M.; Beach, M.J.; Roy, S.L. Causes of outbreaks associated with drinking water in the United States from 1971 to 2006. Clin. Microbiol. Rev. 2010, 23, 507–528. [Google Scholar] [CrossRef] [Green Version]
- Fields, B.; Benson, R.F.; Besser, R.E. Legionella and Legionnaires’ disease: 25 years of investigation. Clin. Microbiol. Rev. 2002, 15, 506–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Declerck, P.; Behets, J.; van Hoef, V.; Ollevier, F. Detection of Legionella spp. and some of their amoeba hosts in floating biofilms from anthropogenic and natural aquatic environments. Water Res. 2007, 41, 3159–3167. [Google Scholar] [CrossRef] [PubMed]
- Koubar, M.; Rodier, M.H.; Garduno, R.A.; Frere, J. Passage through Tetrahymena tropicalis enhances the resistance to stress and the infectivity of Legionella pneumophila. FEMS Microbiol. Lett. 2011, 325, 10–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, M.; Ross, K.; Bentham, R. Legionella, protozoa, and biofilms: Interactions within complex microbial systems. Microb. Ecol. 2009, 58, 538–547. [Google Scholar] [CrossRef] [PubMed]
- van Heijnsbergen, E.; Schalk, J.A.; Euser, S.; Brandsema, P.S.; den Boer, J.W.; De Roda Husman, A.M. Confirmed and potential sources of Legionella reviewed. Environ. Sci. Technol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Rasch, J.; Krüger, S.; Fontvieille, D.; Ünal, C.M.; Michel, R.; Labrosse, A.; Steinert, M. Legionella-protozoa-nematode interactions in aquatic biofilms and influence of Mip on Caenorhabditis elegans colonization. Int. J. Med. Microbiol. 2016, 306, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Parry, J.D. Protozoan grazing of freshwater biofilms. Adv. Appl. Microbiol. 2004, 54, 167–196. [Google Scholar] [PubMed]
- Sherr, B.F.; Sherr, E.B.; Berman, T. Grazing, growth, and ammonium excretion rates of a heterotrophic microflagellate fed with four species of bacteria. Appl. Environ. Microbiol. 1983, 45, 1196–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coşkun, K.A.; Özçelik, S.; Tutar, L.; Elaldı, N.; Tutar, Y. Isolation and identification of free-living amoebae from tap water in Sivas, Turkey. Biomed Res. Int. 2013, 2013, 675145. [Google Scholar] [CrossRef] [Green Version]
- Delafont, V.; Brouke, A.; Bouchon, D.; Moulin, L.; Hechard, Y. Microbiome of free-living amoebae isolated from drinking water. Water Res. 2013, 47, 6958–6965. [Google Scholar] [CrossRef]
- Declerck, P.; Behets, J.; Margineanu, A.; van Hoef, V.; De Keersmaecker, B.; Ollevier, F. Replication of Legionella pneumophila in biofilms of water distribution pipes. Microbiol. Res. 2009, 164, 593–603. [Google Scholar] [CrossRef]
- Murga, R.; Forster, T.S.; Brown, E.; Pruckler, J.M.; Fields, B.S.; Donlan, R.M. Role of biofilms in the survival of Legionella pneumophila in a model potable-water system. Microbiology 2001, 147, 3121–3126. [Google Scholar] [CrossRef] [Green Version]
- Shaheen, M.; Scott, C.; Ashbolt, N.J. Long-term persistence of infectious Legionella with free-living amoebae in drinking water biofilms. Int. J. Hyg. Environ. Health 2019, 222, 678–686. [Google Scholar] [CrossRef]
- Schoen, M.E.; Ashbolt, N.J. An in-premise model for Legionella exposure during showering events. Water Res. 2011, 45, 5826–5836. [Google Scholar] [CrossRef]
- König, L.; Siegl, A.; Penz, T.; Haider, S.; Wentrup, C.; Polzin, J.; Mann, E.; Schmitz-Esser, S.; Domman, D.; Horn, M. Biphasic metabolism and host interaction of a chlamydial symbiont. mSystems 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mou, Q.; Leung, P.H. Differential expression of virulence genes in Legionella pneumophila growing in Acanthamoeba and human monocytes. Virulence 2018, 9, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Okubo, T.; Matsushita, M.; Nakamura, S.; Matsuo, J.; Nagai, H.; Yamaguchi, H. Acanthamoeba S13WT relies on its bacterial endosymbiont to backpack human pathogenic bacteria and resist Legionella infection on solid media. Environ. Microbiol. Rep. 2018, 10, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Schmitz-Esser, S.; Tischler, P.; Arnold, R.; Montanaro, J.; Wagner, M.; Rattei, T.; Horn, M. The genome of the amoeba symbiont “Candidatus Amoebophilus asiaticus” reveals common mechanisms for host cell interaction among amoeba-associated bacteria. J. Bacteriol. 2010, 192, 1045–1057. [Google Scholar] [CrossRef] [Green Version]
- Matz, C.; Kjelleberg, S. Off the hook–how bacteria survive protozoan grazing. Trends Microbiol. 2005, 13, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Jousset, A. Ecological and evolutive implications of bacterial defences against predators. Environ. Microbiol. 2012, 14, 1830–1843. [Google Scholar] [CrossRef]
- Erken, M.; Weitere, M.; Kjelleberg, S.; McDougald, D. In situ grazing resistance of Vibrio cholerae in the marine environment. FEMS Microbiol. Ecol. 2011, 76, 504–512. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa. Int. J. Syst. Evol. Microbiol. 2002, 52, 297–354. [Google Scholar] [CrossRef]
- Hahn, M.W.; Höfle, M.G. Grazing of protozoa and its effect on populations of aquatic bacteria. FEMS Microbiol. Ecol. 2001, 35, 113–121. [Google Scholar] [CrossRef]
- Hoffmann, C.; Harrison, C.F.; Hilbi, H. The natural alternative: Protozoa as cellular models for Legionella infection. Cell. Microbiol. 2014, 16, 15–26. [Google Scholar] [CrossRef]
- Segal, G.; Shuman, H.A. Legionella pneumophila utilizes the same genes to multiply within Acanthamoeba castellanii and human macrophages. Infect. Immun. 1999, 67, 2117–2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Valero, L.; Rusniok, C.; Carson, D.; Mondino, S.; Pérez-Cobas, A.E.; Rolando, M.; Pasricha, S.; Reuter, S.; Demirtas, J.; Crumbach, J. More than 18,000 effectors in the Legionella genus genome provide multiple, independent combinations for replication in human cells. Proc. Natl. Acad. Sci. USA 2019, 116, 2265–2273. [Google Scholar] [CrossRef] [Green Version]
- Al-Quadan, T.; Price, C.T.; Kwaik, Y.A. Exploitation of evolutionarily conserved amoeba and mammalian processes by Legionella. Trends Microbiol. 2012, 20, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.-Q. Striking a balance: Modulation of host cell death pathways by Legionella pneumophila. Front. Microbiol. 2011, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segal, G.; Purcell, M.; Shuman, H.A. Host cell killing and bacterial conjugation require overlapping sets of genes within a 22-kb region of the Legionella pneumophila genome. Proc. Natl. Acad. Sci. USA 1998, 95, 1669–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J.P.; Andrews, H.L.; Wong, S.K.; Isberg, R.R. Conjugative transfer by the virulence system of Legionella pneumophila. Science 1998, 279, 873–876. [Google Scholar] [CrossRef] [PubMed]
- Dopheide, A.; Lear, G.; Stott, R.; Lewis, G. Preferential feeding by the ciliates Chilodonella and Tetrahymena spp. and effects of these protozoa on bacterial biofilm structure and composition. Appl. Environ. Microbiol. 2011, 77, 4564–4572. [Google Scholar] [CrossRef] [Green Version]
- Raghupathi, P.K.; Liu, W.; Sabbe, K.; Houf, K.; Burmølle, M.; Sørensen, S.J. Synergistic Interactions within a Multispecies Biofilm Enhance Individual Species Protection against Grazing by a Pelagic Protozoan. Front. Microbiol. 2018, 8, 2649. [Google Scholar] [CrossRef] [Green Version]
- Amaro, F.; Wang, W.; Gilbert, J.A.; Anderson, O.R.; Shuman, H.A. Diverse protist grazers select for virulence-related traits in Legionella. ISME J. 2015, 9, 1607. [Google Scholar] [CrossRef]
- Huws, S.A.; McBain, A.J.; Gilbert, P. Protozoan grazing and its impact upon population dynamics in biofilm communities. J. Appl. Microbiol. 2005, 98, 238–244. [Google Scholar] [CrossRef]
- Declerck, P.; Behets, J.; Delaedt, Y.; Margineanu, A.; Lammertyn, E.; Ollevier, F. Impact of non-Legionella bacteria on the uptake and intracellular replication of Legionella pneumophila in Acanthamoeba castellanii and Naegleria lovaniensis. Microb. Ecol. 2005, 50, 536–549. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.R.; Muthye, V.; Cianciotto, N.P. Legionella pneumophila persists within biofilms formed by Klebsiella pneumoniae, Flavobacterium sp., and Pseudomonas fluorescens under dynamic flow conditions. PLoS ONE 2012, 7, e50560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, S.; Tateda, K.; Ishii, Y.; Horikawa, M.; Miyairi, S.; Gotoh, N.; Ishiguro, M.; Yamaguchi, K. Pseudomonas aeruginosa Las quorum sensing autoinducer suppresses growth and biofilm production in Legionella species. Microbiology 2009, 155, 1934–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, J.M.; Ashbolt, N.J. Do free-living amoebae in treated drinking water systems present an emerging health risk? Environ. Sci. Technol. 2011, 45, 860–869. [Google Scholar] [CrossRef]
- Falkinham, J.O., 3rd; Hilborn, E.D.; Arduino, M.J.; Pruden, A.; Edwards, M.A. Epidemiology and Ecology of Opportunistic Premise Plumbing Pathogens: Legionella pneumophila, Mycobacterium avium, and Pseudomonas aeruginosa. Environ. Health Perspect. 2015. [Google Scholar] [CrossRef] [Green Version]
- Morash, M.G.; Brassinga, A.K.C.; Warthan, M.; Gourabathini, P.; Garduno, R.A.; Goodman, S.D.; Hoffman, P.S. Reciprocal expression of integration host factor and HU in the developmental cycle and infectivity of Legionella pneumophila. Appl. Environ. Microbiol. 2009, 75, 1826–1837. [Google Scholar] [CrossRef] [Green Version]
- Piao, Z.; Sze, C.C.; Barysheva, O.; Iida, K.; Yoshida, S. Temperature-regulated formation of mycelial mat-like biofilms by Legionella pneumophila. Appl. Environ. Microbiol. 2006, 72, 1613–1622. [Google Scholar] [CrossRef] [Green Version]
- Shaheen, M.; Ashbolt, N.J. Free-living amoebae supporting intracellular growth may produce vesicle-bound respirable doses of Legionella within drinking water systems. Expos. Health 2018, 10, 201–209. [Google Scholar] [CrossRef]
- Benchimol, M. The release of secretory vesicle in encysting Giardia lamblia. FEMS Microbiol. Lett. 2004, 235, 81–87. [Google Scholar] [CrossRef]
- Marti, M.; Hehl, A.B. Encystation-specific vesicles in Giardia: A primordial Golgi or just another secretory compartment? Trends Parasitol. 2003, 19, 440–446. [Google Scholar] [CrossRef]
- Greub, G.; Raoult, D. Morphology of Legionella pneumophila according to their location within Hartmanella vermiformis. Res. Microbiol. 2003, 154, 619–621. [Google Scholar] [CrossRef] [PubMed]
- Kilvington, S.; Price, J. Survival of Legionella pneumophila within cysts of Acanthamoeba polyphaga following chlorine exposure. J. Appl. Bacteriol. 1990, 68, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Stout, J.E.; Muder, R.R.; Mietzner, S.; Wagener, M.M.; Perri, M.B.; DeRoos, K.; Goodrich, D.; Arnold, W.; Williamson, T.; Ruark, O.; et al. Role of environmental surveillance in determining the risk of hospital-acquired legionellosis: A national surveillance study with clinical correlations. Infect. Control Hosp. Epidemiol. 2007, 28, 818–824. [Google Scholar] [CrossRef]
- Bernander, S.; Jacobson, K.; Helbig, J.H.; Lück, P.C.; Lundholm, M. A hospital-associated outbreak of Legionnaires’ disease caused by Legionella pneumophila serogroup 1 is characterized by stable genetic fingerprinting but variable monoclonal antibody patterns. J. Clin. Microbiol. 2003, 41, 2503–2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernander, S.; Jacobson, K.; Lundholm, M. A hospital-associated outbreak of Legionnaires′ disease caused by Legionella pneumophila serogroups 4 and 10 with a common genetic fingerprinting pattern. APMIS 2004, 112, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Oberdorfer, K.; Müssigbrodt, G.; Wendt, C. Genetic diversity of Legionella pneumophila in hospital water systems. Int. J. Hyg. Environ. Health 2008, 211, 172–178. [Google Scholar] [CrossRef]
- Phin, N.; Parry-Ford, F.; Harrison, T.; Stagg, H.R.; Zhang, N.; Kumar, K.; Lortholary, O.; Zumla, A.; Abubakar, I. Epidemiology and clinical management of Legionnaires’ disease. Lancet. Infect. Dis. 2014, 14, 1011–1021. [Google Scholar] [CrossRef]
- MacIntyre, D.L.; Miyata, S.T.; Kitaoka, M.; Pukatzki, S. The Vibrio cholerae type VI secretion system displays antimicrobial properties. Proc. Natl. Acad. Sci. USA 2010, 107, 19520–19524. [Google Scholar] [CrossRef] [Green Version]
- Miyata, S.T.; Kitaoka, M.; Brooks, T.M.; McAuley, S.B.; Pukatzki, S. Vibrio cholerae requires the type VI secretion system virulence factor VasX to kill Dictyostelium discoideum. Infect. Immun. 2011, 79, 2941–2949. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, M.; Binet, M.; Bouteleux, C.; Herbelin, P.; Soreau, S.; Hechard, Y. Permissiveness of freshly isolated environmental strains of amoebae for growth of Legionella pneumophila. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef]
- Fields, B.S.; Sanden, G.N.; Barbaree, J.M.; Morrill, W.E.; Wadowsky, R.M.; White, E.H.; Feeley, J.C. Intracellular multiplication of Legionella pneumophila in amoebae isolated from hospital hot water tanks. Curr. Microbiol. 1989, 18, 131–137. [Google Scholar] [CrossRef]
- Wang, H.; Edwards, M.A.; Falkinham, J.O., III; Pruden, A. Probiotic approach to pathogen control in premise plumbing systems? A review. Environ. Sci. Technol. 2013, 47, 10117–10128. [Google Scholar] [CrossRef]
- Lu, J.; Buse, H.; Gomez-Alvarez, V.; Struewing, I.; Santo Domingo, J.; Ashbolt, N.J. Impact of drinking water conditions and copper materials on downstream biofilm microbial communities and Legionella pneumophila colonization. J. Appl. Microbiol. 2014, 117, 905–918. [Google Scholar] [CrossRef]
- Pinto, A.J.; Xi, C.; Raskin, L. Bacterial community structure in the drinking water microbiome is governed by filtration processes. Environ. Sci. Technol. 2012, 46, 8851–8859. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaheen, M.; Ashbolt, N.J. Differential Bacterial Predation by Free-Living Amoebae May Result in Blooms of Legionella in Drinking Water Systems. Microorganisms 2021, 9, 174. https://doi.org/10.3390/microorganisms9010174
Shaheen M, Ashbolt NJ. Differential Bacterial Predation by Free-Living Amoebae May Result in Blooms of Legionella in Drinking Water Systems. Microorganisms. 2021; 9(1):174. https://doi.org/10.3390/microorganisms9010174
Chicago/Turabian StyleShaheen, Mohamed, and Nicholas J. Ashbolt. 2021. "Differential Bacterial Predation by Free-Living Amoebae May Result in Blooms of Legionella in Drinking Water Systems" Microorganisms 9, no. 1: 174. https://doi.org/10.3390/microorganisms9010174
APA StyleShaheen, M., & Ashbolt, N. J. (2021). Differential Bacterial Predation by Free-Living Amoebae May Result in Blooms of Legionella in Drinking Water Systems. Microorganisms, 9(1), 174. https://doi.org/10.3390/microorganisms9010174