
Composition and Dominance of Edible and Inedible Phytoplankton Predict Responses of Baltic Sea Summer Communities to Elevated Temperature and CO2



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sampling and Measurements
2.3. Data Analysis
2.4. Statistical Analyses
3. Results
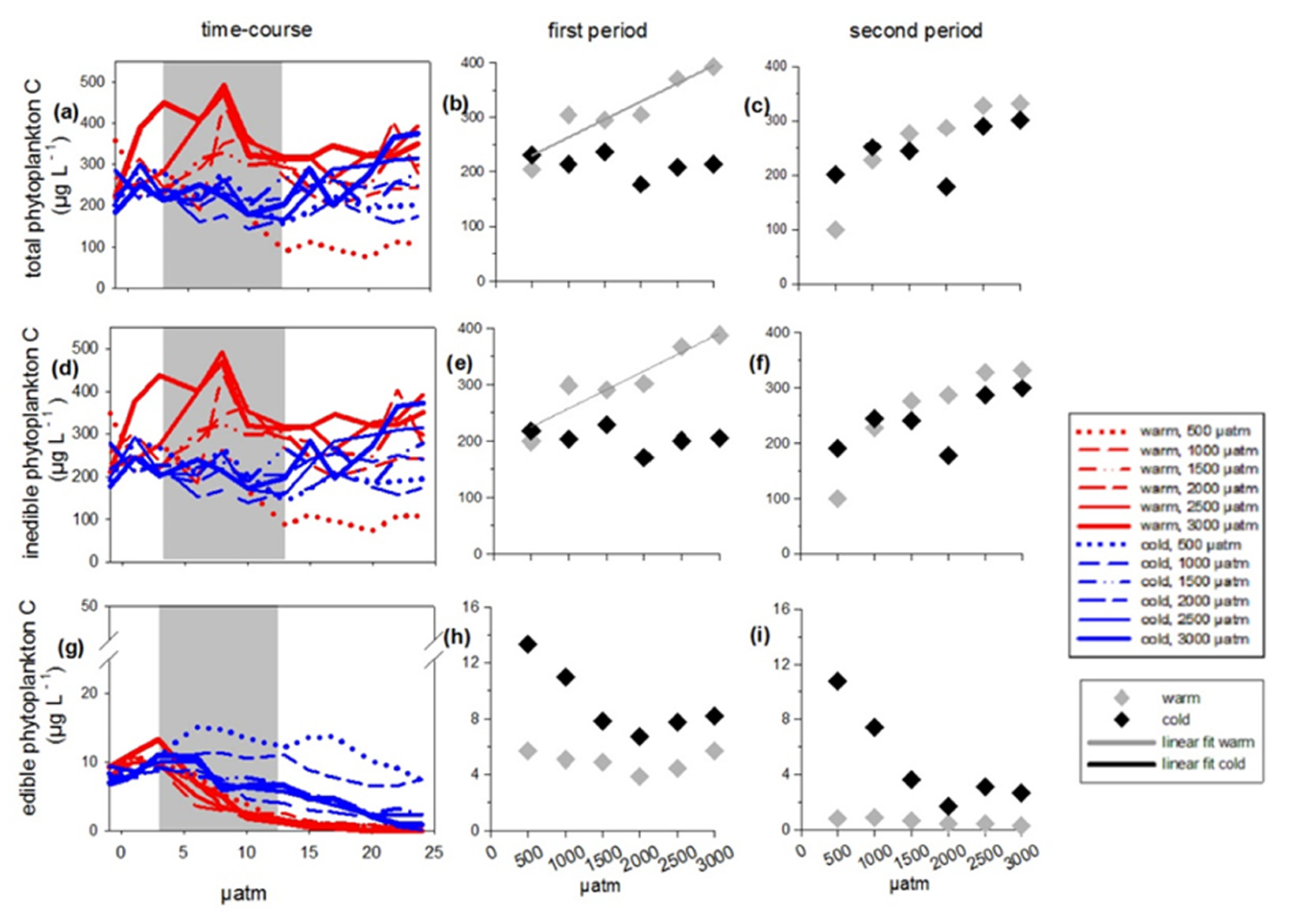
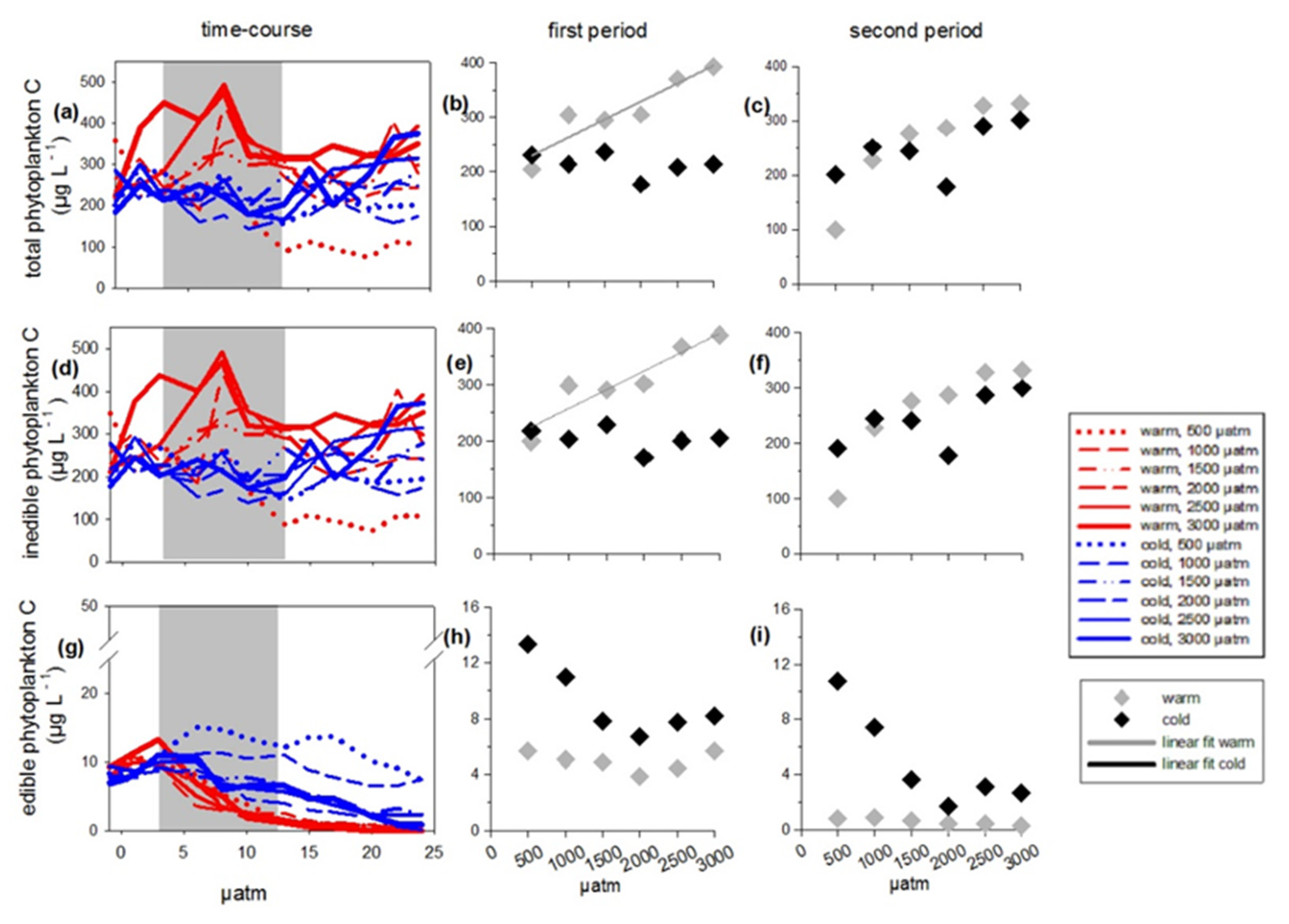
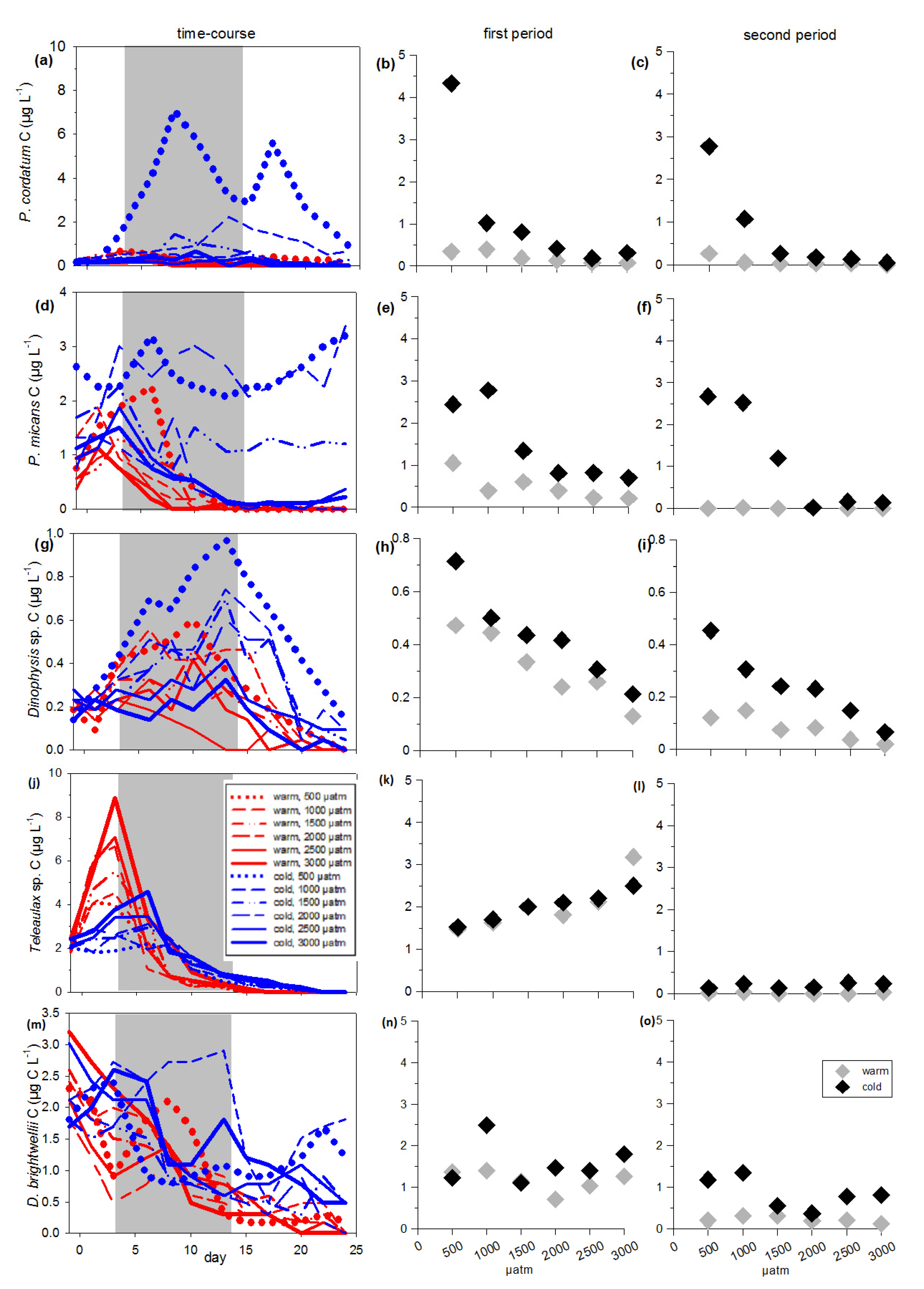
3.1. Total Phytoplankton Carbon
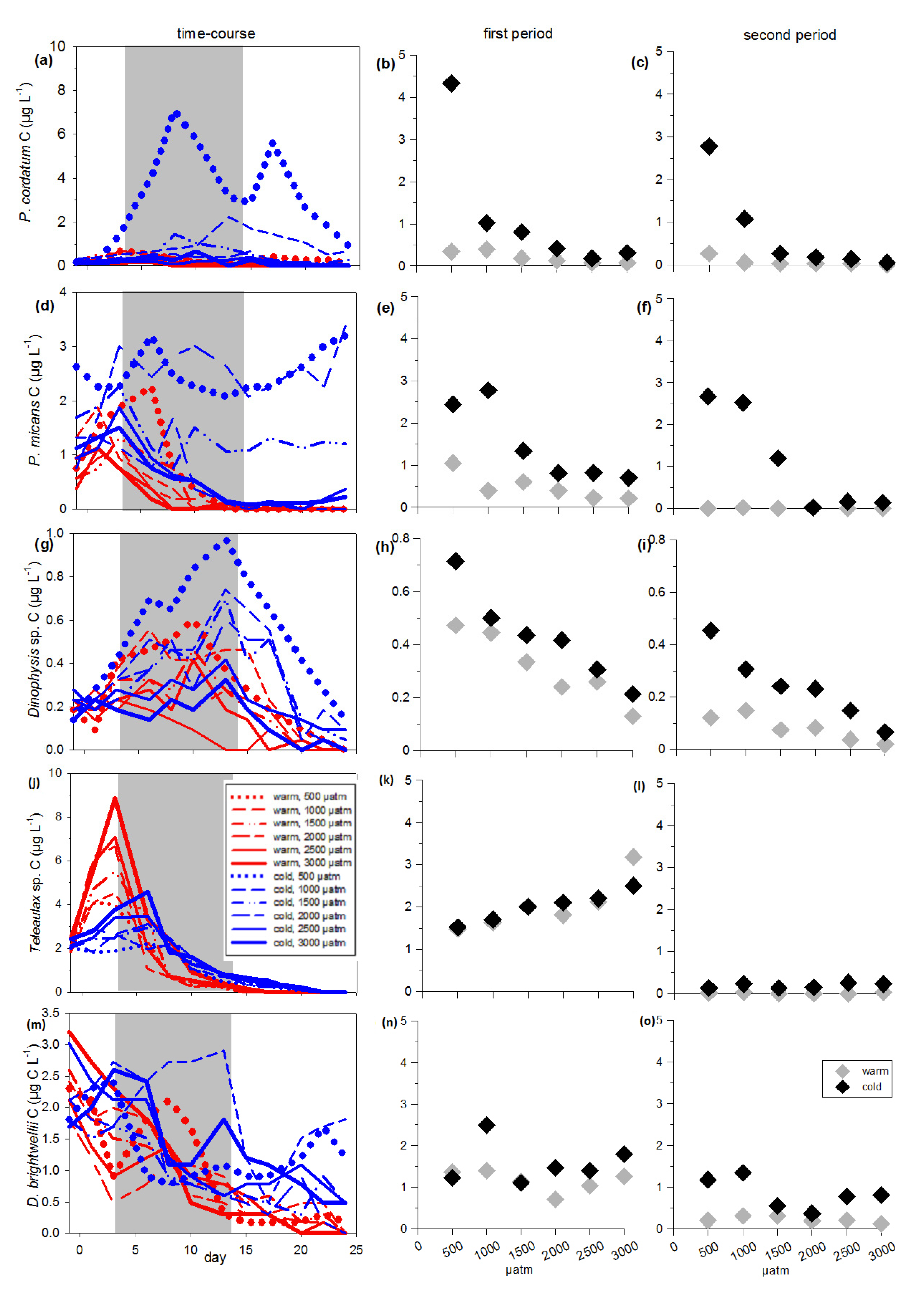
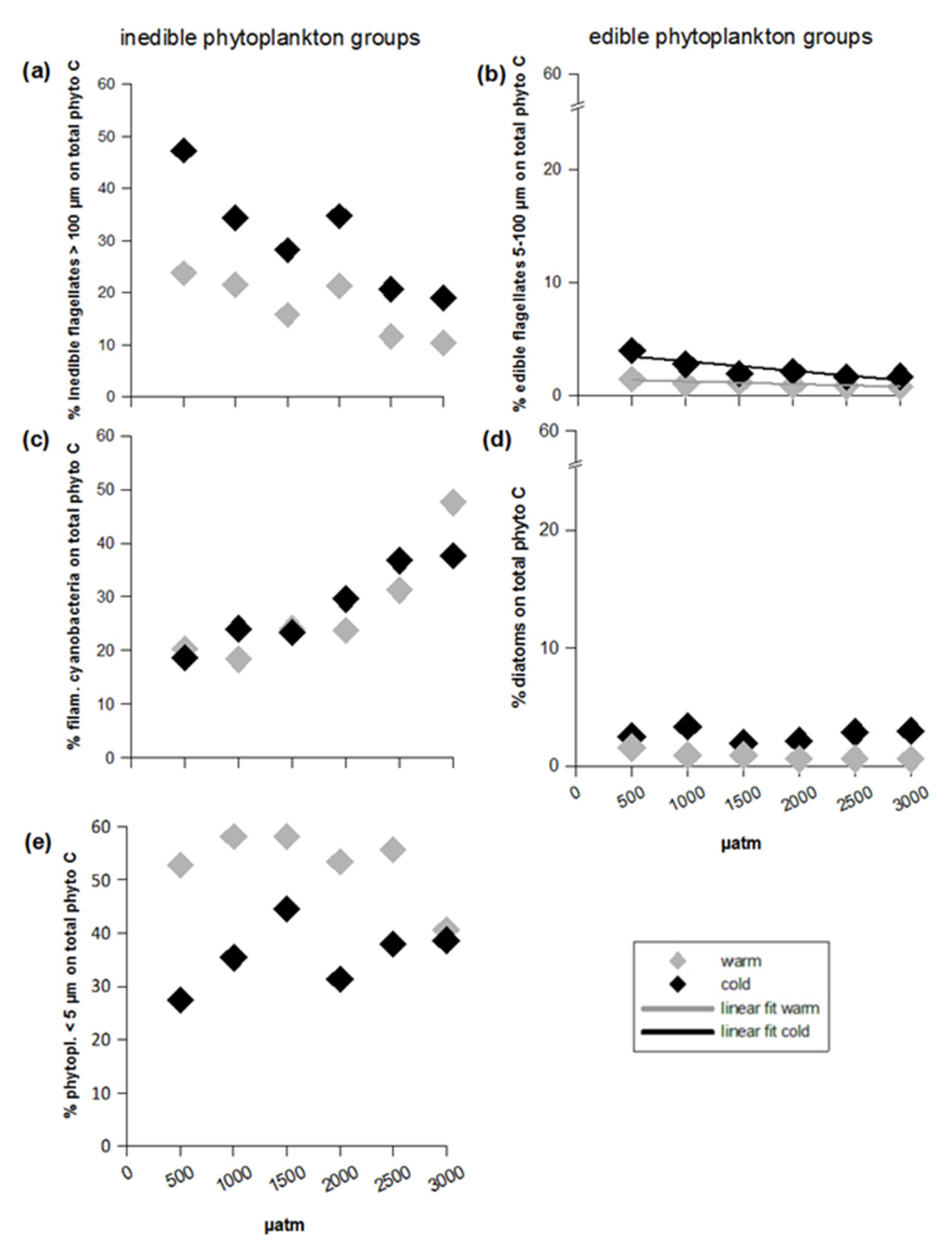
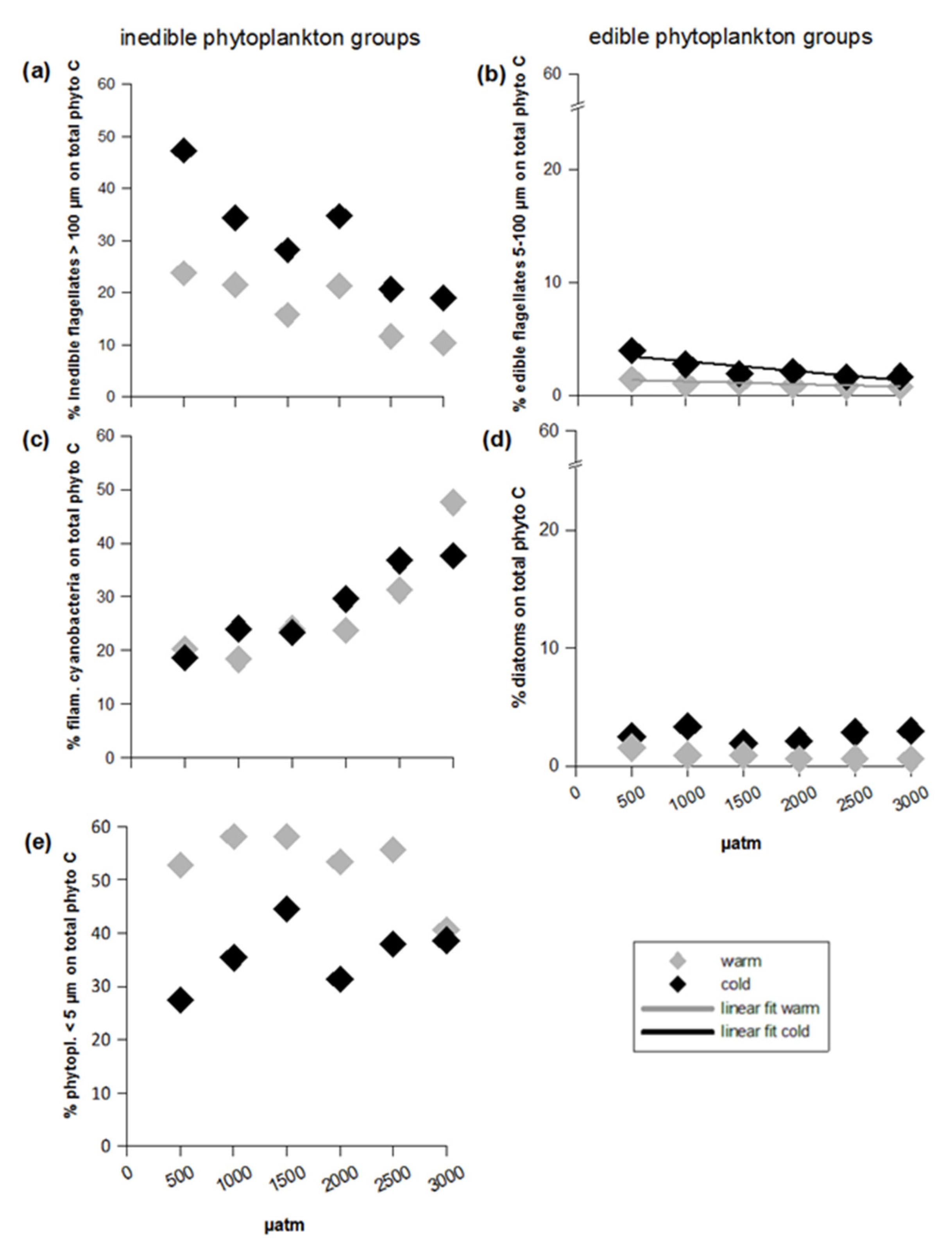
3.2. Inedible Phytoplankton Groups
3.3. Edible Phytoplankton Groups
3.4. Dissolved Inorganic Nutrients
3.5. Particulate Organic Matter Stoichiometry
4. Discussion
4.1. Effects of Temperature and CO2 on the Inedible Fraction
4.2. Effects of Temperature and CO2 on the Edible Fraction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable | Factor | df Residual | t-Value | p |
|---|---|---|---|---|
| Entire course of experimental time | ||||
| (log) total phytoplankton C (µg L−1) | time × T | 136 | −2.44663 | 0.0157 * |
| time × CO2 | 136 | 2.27327 | 0.0246 * | |
| (log) inedible phytoplankton C (µg L−1) | time × T | 136 | −2.23612 | 0.0270 * |
| time × CO2 | 136 | 2.44929 | 0.0156 * | |
| Edible phytoplankton C (µg L−1) | time × T | 136 | −4.37454 | <0.001 *** |
| time × CO2 | 136 | −4.00765 | <0.001 *** | |
| Inedible flagellates > 100 µm C (µg L−1) | time × T | 136 | −2.15323 | 0.0331 * |
| (log) filamentous cyanobacteria (µg L−1) | T × CO2 | 136 | 2.231858 | 0.0274 * |
| time × CO2 | 136 | 2.758938 | 0.0067 ** | |
| Edible flagellates 5–100 µm C (µg L−1) | T | 136 | −2.13354 | 0.0347 * |
| CO2 | 136 | −2.30590 | 0.0226 * | |
| T × CO2 | 136 | 2.024677 | 0.0449 * | |
| time × T | 136 | −2.84060 | 0.0052 ** | |
| time x CO2 | 136 | −2.86083 | 0.0049 ** | |
| Diatom C (µg L−1) | time × T | 136 | −4.12406 | <0.001 *** |
| time x CO2 | 136 | −3.57416 | <0.001 *** | |
| time × T × CO2 | 136 | 2.750142 | 0.0068 ** | |
| (log) C:N | CO2 | 136 | −2.32444 | 0.0216 * |
| PO43− (µg L−1) | T | 136 | 2.785997 | 0.0061 ** |
| First period | ||||
| % inedible flagellates > 100 µm | T | 8 | −3.93871 | 0.0043 ** |
| on total phytopl. C | CO2 | 8 | −4.95259 | 0.0011 ** |
| % edible flagellates 5–100 µm on total phytopl. C | T | 8 | −5.18685 | <0.001 *** |
| CO2 | 8 | −5.02835 | 0.0010 ** | |
| T × CO2 | 8 | 2.562598 | 0.0335 * | |
| % filam. cyanobacteria on total phytopl. C | CO2 | 8 | 3.644613 | 0.0065 ** |
| % phytopl. <5 µm on total phytopl. C | T | 8 | 3.831281 | 0.0050 ** |
| (log) inedible flagellates >100 µm C | T | 8 | −3.25951 | 0.0115 * |
| (µg L−1) | CO2 | 8 | −5.12617 | <0.001 ** |
| T × CO2 | 8 | 2.35489 | 0.0463 * | |
| (log) filamentous cyanobacteria C (µg L−1) | CO2 | 8 | 3.320493 | 0.0105 * |
| T × CO2 | 8 | 2.891799 | 0.0201 * | |
| (log) C:N | CO2 | 8 | 3.127289 | 0.0141 * |
| C:P | T × CO2 | 8 | 2.586698 | 0.0323 * |
| N:P | CO2 | 8 | 2.491033 | 0.0375 * |
| Second period | ||||
| (log) edible phytoplankton C (µg L−1) | T | 8 | −5.00670 | 0.0010 ** |
| CO2 | 8 | −3.67592 | 0.0063 ** | |
| (log) filamentous cyanobacteria C (µg L−1) | CO2 | 8 | 6.531437 | <0.001 **** |
| (log) edible flagellates 5–100 µm C (µg L−1) | T | 8 | −6.69378 | <0.001 *** |
| CO2 | 8 | −6.92440 | <0.001 *** | |
| (log) diatom C (µg L−1) | T | 8 | −3.15491 | 0.0135 * |
| Response Variable | Factor | df Residual | t-Value | p |
|---|---|---|---|---|
| Total phytoplankton C first period (µg L−1) | CO2 warm | 4 | 5.086691 | 0.0070 ** |
| CO2 cold | 4 | −0.928886 | 0.4055 | |
| Inedible phytoplankton C first period (µg L−1) | CO2 warm | 4 | 5.107762 | 0.0069 ** |
| CO2 cold | 4 | −0.743826 | 0.4983 | |
| Inedible flagellates > 100 µm C first period (µg L−1) | CO2 warm | 4 | −1.571086 | 0.1913 |
| CO2 cold | 4 | 5.005789 | 0.0075 ** | |
| Filamentous cyanobacteria C first period (µg L−1) | CO2 warm | 4 | 3.982850 | 0.0164 * |
| CO2 cold | 4 | 4.209925 | 0.0136 * | |
| Edible flagellates 5–100 µm C first period (µg L−1) | CO2 warm | 4 | 0.053824 | 0.9597 |
| CO2 cold | 4 | −3.590508 | 0.0230 * | |
| % edible flagellates 5–100 µm on | CO2 warm | 4 | −3.685677 | <0.001 *** |
| total phytoplankton C (µg L−1) | CO2 cold | 4 | −3.692101 | 0.0211 * |
| C:N second period | CO2 warm | 4 | 1.29419 | 0.2653 |
| CO2 cold | 4 | −10.16259 | <0.001 *** | |
| C:P first period | CO2 warm | 4 | 2.761417 | 0.0508 * |
| CO2 cold | 4 | 0.042495 | 0.9681 |
References
- IPCC. Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2019. [Google Scholar]
- Beardall, J.; Raven, J.A. The potential effects of global climate change on microalgal photosynthesis, growth and ecology. Phycologia 2004, 43, 26–40. [Google Scholar] [CrossRef]
- Caldeira, K.; Wickett, M.E. Anthropogenic carbon and ocean pH. Nature 2003, 425, 365. [Google Scholar] [CrossRef] [PubMed]
- IPPC. Climate Change 2014: Impacts, Adaptation and Vulnerability; IPCC Working Group II Contribution to the Fifth Assessement Report of the International Panel on Climate Change. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, J.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1–32. [Google Scholar]
- IPPC. Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emision Pathways in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2018. [Google Scholar]
- Siegel, H.; Gerth, M. Development of Sea Surface Temperature (SST) in the Baltic Sea 2015. HELCOM Baltic Sea Environment Fact Sheets. 2015. Available online: http://www.helcom.fi/baltic-sea-trend/environment-fact-sheets/ (accessed on 24 August 2021).
- Hoegh-Guldberg, O.; Bruno, J.F. The Impact of Climate Change on the World’s Marine Ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef]
- Guinder, V.A.; Molinero, J.C. Climate change effects on marine phytoplankton. In Marine Ecology in a Changing World; Hugo, A.A., Menendez, M.C., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2013; pp. 68–90. [Google Scholar]
- Lewandowska, A.; Sommer, U. Climate change and the spring bloom: A mesocosm study on the influence of light and temperature on phytoplankton and mesozooplankton. Mar. Ecol. Prog. Ser. 2010, 405, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Keller, A.A.; Oviatt, C.A.; Walker, H.A.; Hawk, J.D. Predicted impacts of elevated temperature on the magnitude of the winter-spring phytoplankton bloom in temperate coastal waters: A mesocosm study. Limnol. Oceanogr. 1999, 44, 344–356. [Google Scholar] [CrossRef] [Green Version]
- Paul, C.; Matthiessen, B.; Sommer, U. Warming, but not enhanced CO2 concentration, quantitatively and qualitatively affects phytoplankton biomass. Mar. Ecol. Prog. Ser. 2015, 528, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Sommer, U.; Lewandowska, A. Climate change and the phytoplankton spring bloom: Warming and overwintering zooplankton have similar effects on phytoplankton. Glob. Chang. Biol. 2011, 17, 154–162. [Google Scholar] [CrossRef]
- Sommer, U.; Lengfellner, K. Climate change and the timing, magnitude, and composition of the phytoplankton spring bloom. Glob. Chang. Biol. 2008, 14, 1199–1208. [Google Scholar] [CrossRef]
- Lewandowska, A.M.; Boyce, D.G.; Hofmann, M.; Matthiessen, B.; Sommer, U.; Worm, B. Effects of sea surface warming on marine plankton. Ecol. Lett. 2014, 17, 614–623. [Google Scholar] [CrossRef]
- Suikkanen, S.; Pulina, S.; Engstrom-Ost, J.; Lehtiniemi, M.; Lehtinen, S.; Brutemark, A. Climate Change and Eutrophication Induced Shifts in Northern Summer Plankton Communities. PLoS ONE 2013, 8, e66475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, C.; Sommer, U.; Garzke, J.; Moustaka-Gouni, M.; Paul, A.; Matthiessen, B. Effects of increased CO2 concentration on nutrient limited coastal summer plankton depend on temperature. Limnol. Oceanogr. 2016, 61, 853–868. [Google Scholar] [CrossRef] [Green Version]
- Seifert, M.; Rost, B.; Trimborn, S.; Hauck, J. Meta-analysis of multiple driver effects on marine phytoplankton highlights modulating role of pCO(2). Glob. Chang. Biol. 2020, 26, 6787–6804. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.J.; Bach, L.T. Universal response pattern of phytoplankton growth rates to increasing CO2. New Phytol. 2020, 228, 1710–1716. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Kordas, R.L.; Crim, R.; Hendriks, I.E.; Ramajo, L.; Singh, G.S.; Duarte, C.M.; Gattuso, J.P. Impacts of ocean acidification on marine organisms: Quantifying sensitivities and interaction with warming. Glob. Chang. Biol. 2013, 19, 1884–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raven, J.A. Physiology of inorganic C aquisition and implications for resource use efficiency by marine phytoplankton—Relation to increased CO2 and temperature. Plant Cell Environ. 1991, 14, 779–794. [Google Scholar] [CrossRef]
- Hopkinson, B.M.; Dupont, C.L.; Allen, A.E.; Morel, F.M.M. Efficiency of the CO2-concentrating mechanism of diatoms. Proc. Natl. Acad. Sci. USA 2011, 108, 3830–3837. [Google Scholar] [CrossRef] [Green Version]
- Rost, B.; Zondervan, I.; Wolf-Gladrow, D. Sensitivity of phytoplankton to future changes in ocean carbonate chemistry: Current knowledge, contradictions and research directions. Mar. Ecol. Prog. Ser. 2008, 373, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Burkhardt, S.; Amoroso, G.; Riebesell, U.; Sultemeyer, D. CO2 and HCO3- uptake in marine diatoms acclimated to different CO2 concentrations. Limnol. Oceanogr. 2001, 46, 1378–1391. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A.; Beardall, J. CO2 concentrating mechanisms and environmental change. Aquat. Bot. 2014, 118, 24–37. [Google Scholar] [CrossRef]
- Reinfelder, J.R. Carbon Concentrating Mechanisms in Eukaryotic Marine Phytoplankton. Annu. Rev. Mar. Sci. 2011, 3, 291–315. [Google Scholar] [CrossRef] [Green Version]
- Wannicke, N.; Endres, S.; Engel, A.; Grossart, H.P.; Nausch, M.; Unger, J.; Voss, M. Response of Nodularia spumigena to pCO(2)—Part 1: Growth, production and nitrogen cycling. Biogeosciences 2012, 9, 2973–2988. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Campbell, D.A.; Irwin, A.J.; Suggett, D.J.; Finkel, Z.V. Ocean acidification enhances the growth rate of larger diatoms. Limnol. Oceanogr. 2014, 59, 1027–1034. [Google Scholar] [CrossRef]
- Bach, L.T.; Taucher, J. CO2 effects on diatoms: A synthesis of more than a decade of ocean acidification experiments with natural communities. Ocean Sci. 2019, 15, 1159–1175. [Google Scholar] [CrossRef] [Green Version]
- Eggers, S.L.; Lewandowska, A.M.; Ramos, J.B.E.; Blanco-Ameijeiras, S.; Gallo, F.; Matthiessen, B. Community composition has greater impact on the functioning of marine phytoplankton communities than ocean acidification. Glob. Chang. Biol. 2014, 20, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Tortell, P.D.; Payne, C.; Gueguen, C.; Strzepek, R.F.; Boyd, P.W.; Rost, B. Inorganic carbon uptake by Southern Ocean phytoplankton. Limnol. Oceanogr. 2008, 53, 1266–1278. [Google Scholar] [CrossRef]
- Bach, L.T.; Alvarez-Fernandez, S.; Hornick, T.; Stuhr, A.; Riebesell, U. Simulated ocean acidification reveals winners and losers in coastal phytoplankton. PLoS ONE 2017, 12, e0188198. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.J.; Achterberg, E.P.; Bach, L.T.; Boxhammer, T.; Czerny, J.; Haunost, M.; Schulz, K.G.; Stuhr, A.; Riebesell, U. No observed effect of ocean acidification on nitrogen biogeochemistry in a summer Baltic Sea plankton community. Biogeosciences 2016, 13, 3901–3913. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.J.; Sommer, U.; Paul, C.; Riebesell, U. Baltic Sea diazotrophic cyanobacterium is negatively affected by acidification and warming. Mar. Ecol. Prog. Ser. 2018, 598, 49–60. [Google Scholar] [CrossRef]
- Keys, M.; Tilstone, G.; Findlay, H.S.; Widdicombe, C.E.; Lawson, T. Effects of elevated CO2 and temperature on phytoplankton community biomass, species composition and photosynthesis during an experimentally induced autumn bloom in the western English Channel. Biogeosciences 2018, 15, 3203–3222. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.J.; Bach, L.T.; Schulz, K.G.; Boxhammer, T.; Czerny, J.; Achterberg, E.P.; Hellemann, D.; Trense, Y.; Nausch, M.; Sswat, M.; et al. Effect of elevated CO2 on organic matter pools and fluxes in a summer Baltic Sea plankton community. Biogeosciences 2015, 12, 6181–6203. [Google Scholar] [CrossRef] [Green Version]
- Wohlers-Zollner, J.; Biermann, A.; Engel, A.; Dorge, P.; Lewandowska, A.M.; von Scheibner, M.; Riebesell, U. Effects of rising temperature on pelagic biogeochemistry in mesocosm systems: A comparative analysis of the AQUASHIFT Kiel experiments. Mar. Biol. 2012, 159, 2503–2518. [Google Scholar] [CrossRef] [Green Version]
- Schulz, K.G.; Bellerby, R.G.J.; Brussaard, C.P.D.; Budenbender, J.; Czerny, J.; Engel, A.; Fischer, M.; Koch-Klavsen, S.; Krug, S.A.; Lischka, S.; et al. Temporal biomass dynamics of an Arctic plankton bloom in response to increasing levels of atmospheric carbon dioxide. Biogeosciences 2013, 10, 161–180. [Google Scholar] [CrossRef] [Green Version]
- Tortell, P.D.; DiTullio, G.R.; Sigman, D.M.; Morel, F.M.M. CO2 effects on taxonomic composition and nutrient utilization in an Equatorial Pacific phytoplankton assemblage. Mar. Ecol. Prog. Ser. 2002, 236, 37–43. [Google Scholar] [CrossRef]
- Tortell, P.D.; Rau, G.H.; Morel, F.M.M. Inorganic carbon acquisition in coastal Pacific phytoplankton communities. Limnol. Oceanogr. 2000, 45, 1485–1500. [Google Scholar] [CrossRef]
- Sett, S.; Schulz, K.G.; Bach, L.T.; Riebesell, U. Shift towards larger diatoms in a natural phytoplankton assemblage under combined high-CO2 and warming conditions. J. Plankton Res. 2018, 40, 391–406. [Google Scholar] [CrossRef] [Green Version]
- Hare, C.E.; Leblanc, K.; DiTullio, G.R.; Kudela, R.M.; Zhang, Y.; Lee, P.A.; Riseman, S.; Hutchins, D.A. Consequences of increased temperature and CO2 for phytoplankton community structure in the Bering Sea. Mar. Ecol. Prog. Ser. 2007, 352, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.Y.; Hare, C.E.; Leblanc, K.; Rose, J.M.; Zhang, Y.H.; DiTullio, G.R.; Lee, P.A.; Wilhelm, S.W.; Rowe, J.M.; Sun, J.; et al. Effects of increased pCO(2) and temperature on the North Atlantic spring bloom. I. The phytoplankton community and biogeochemical response. Mar. Ecol. Prog. Ser. 2009, 388, 13–25. [Google Scholar] [CrossRef]
- Torstensson, A.; Chierici, M.; Wulff, A. The influence of increased temperature and carbon dioxide levels on the benthic/sea ice diatom Navicula directa. Polar Biol. 2012, 35, 205–214. [Google Scholar] [CrossRef]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Lurling, M.; Van Oosterhout, F.; Faassen, E. Eutrophication and Warming Boost Cyanobacterial Biomass and Microcystins. Toxins 2017, 9, 64. [Google Scholar] [CrossRef] [Green Version]
- Litchman, E.; Pinto, P.D.; Edwards, K.F.; Klausmeier, C.A.; Kremer, C.T.; Thomas, M.K. Global biogeochemical impacts of phytoplankton: A trait-based perspective. J. Ecol. 2015, 103, 1384–1396. [Google Scholar] [CrossRef] [Green Version]
- Sommer, U.; Hansen, T.; Blum, O.; Holzner, N.; Vadstein, O.; Stibor, H. Copepod and microzooplankton grazing in mesocosms fertilised with different Si:N ratios: No overlap between food spectra and Si:N influence on zooplankton trophic level. Oecologia 2005, 142, 274–283. [Google Scholar] [CrossRef]
- Katechakis, A.; Stibor, H.; Sommer, U.; Hansen, T. Feeding selectivities and food niche separation of Acartia clausi, Penilia avirostris (Crustacea) and Doliolum denticulatum (Thaliacea) in Blanes Bay (Catalan Sea, NW Mediterranean). J. Plankton Res. 2004, 26, 589–603. [Google Scholar] [CrossRef] [Green Version]
- Sommer, U.; Sommer, F. Cladocerans versus copepods: The cause of contrasting top-down controls on freshwater and marine phytoplankton. Oecologia 2006, 147, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Garzke, J.; Hansen, T.; Ismar, S.M.H.; Sommer, U. Combined Effects of Ocean Warming and Acidification on Copepod Abundance, Body Size and Fatty Acid Content. PLoS ONE 2016, 11, e0155952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garzke, J.; Sommer, U.; Ismar, S.M.H. Is the chemical composition of biomass the agent by which ocean acidification influences on zooplankton ecology? Aquat. Sci. 2017, 79, 733–748. [Google Scholar] [CrossRef] [Green Version]
- Garzke, J.; Sommer, U.; Ismar-Rebitz, S.M.H. Zooplankton growth and survival differentially respond to interactive warming and acidification effects. J. Plankton Res. 2020, 42, 189–202. [Google Scholar] [CrossRef]
- Lennartz, S.T.; Lehmann, A.; Herrford, J.; Malien, F.; Hansen, H.P.; Biester, H.; Bange, H.W. Long-term trends at the Boknis Eck time series station (Baltic Sea), 1957–2013: Does climate change counteract the decline in eutrophication? Biogeosciences 2014, 11, 6323–6339. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, J.; Gutowska, M.A.; Saphorster, J.; Heinemann, A.; Trubenbach, K.; Fietzke, J.; Hiebenthal, C.; Eisenhauer, A.; Kortzinger, A.; Wahl, M.; et al. Calcifying invertebrates succeed in a naturally CO2-rich coastal habitat but are threatened by high levels of future acidification. Biogeosciences 2010, 7, 3879–3891. [Google Scholar] [CrossRef] [Green Version]
- Melzner, F.; Thomsen, J.; Koeve, W.; Oschlies, A.; Gutowska, M.A.; Bange, H.W.; Hansen, H.P.; Kortzinger, A. Future ocean acidification will be amplified by hypoxia in coastal habitats. Mar. Biol. 2013, 160, 1875–1888. [Google Scholar] [CrossRef]
- Lewis, E. Program Developed for CO2 System Calculations; Oak Ridge National Laboratory ORNL/CDIAC: Oak Ridge, TN, USA, 1998. [Google Scholar]
- Brock, T.D. Calculating solar radiation for ecological studies. Ecol. Model 1981, 14, 1–19. [Google Scholar] [CrossRef]
- Hansen, T.; Gardeler, B.; Matthiessen, B. Technical Note: Precise quantitative measurements of total dissolved inorganic carbon from small amounts of seawater using a gas chromatographic system. Biogeosciences 2013, 10, 6601–6608. [Google Scholar] [CrossRef]
- Dickson, A.G. An exact definition of total alkalinity and procedure for the estimation of alkalinity and total inorganic carbon from titration data. Deep Sea Res. 1981, 28, 609–623. [Google Scholar] [CrossRef]
- Dickson, A.G.; Afghan, J.D.; Anderson, G.C. Reference materials for oceanic CO2 analysis: A method for the certification of total alkalinity. Mar. Chem. 2003, 80, 185–197. [Google Scholar] [CrossRef]
- Pierrot, D.; Lewis, E. MS Excel Program Developed for CO2 System Calculations: ORNL/CDIAC-105a; Oak Ridge National Laboratory ORNL/CDIAC: Oak Ridge, TN, USA, 2006. [Google Scholar]
- Hansson, I. A new set of acidityconstants for carbonic acid and boric acid in seawater. Deep Seawater Res. 1973, 20, 661–678. [Google Scholar]
- Mehrbach, C.; Culberson, C.H.; Hawley, J.E.; Pytkowicz, R.M. Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnol. Oceanogr. 1973, 18, 897–907. [Google Scholar] [CrossRef]
- Dickson, A.G.; Millero, F.J. A comparison of the equilibrium constants for the dissociations of carbonic acid in seawater media. Deep Sea Res. 1987, 34, 1733–1741. [Google Scholar] [CrossRef]
- Dickson, A.G. Standard potential of the reaction: AgCl(s) + 1/2H2(s) + HCl(aqu) and the standard acidity constant of the ion HSO4− in synthetic sea water from 273.15 to 318.15 K. J. Chem. Thermodyn. 1990, 22, 113–127. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Mitt. Int. Ver. Theor. Angew. Limnol. 1958, 9, 263–272. [Google Scholar] [CrossRef]
- Hillebrand, H.; Durselen, C.D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Menden-Deuer, S.; Lessard, E.J. Carbon to volume relationships for dinoflagellates, diatoms, and other protist plankton. Limnol. Oceanogr. 2000, 45, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Sommer, U.; Lengfellner, K.; Lewandowska, A. Experimental induction of a coastal spring bloom early in the year by intermittent high-light episodes. Mar. Ecol. Prog. Ser. 2012, 446, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Hansen, H.P.; Koroleff, F. Determination of nutrients. In Methods of Seawater Analysis, 3rd ed.; Grasshoff, K., Kremling, K., Erhardt, M., Eds.; Wiley VCH: Weinheim, Germany, 1999; pp. 159–228. [Google Scholar]
- Sommer, F.; Stibor, H.; Sommer, U.; Velimirov, B. Grazing by mesozooplankton from Kiel Bight, Baltic Sea, on different sized algae and natural seston size fractions. Mar. Ecol. Prog. Ser. 2000, 199, 43–53. [Google Scholar] [CrossRef]
- Raateoja, M.; Kuosa, H.; Hallfors, S. Fate of excess phosphorus in the Baltic Sea: A real driving force for cyanobacterial blooms? J. Sea Res. 2011, 65, 315–321. [Google Scholar] [CrossRef]
- Niemistoe, L.; Rinne, I.; Melvasalo, T.; Niemi, A. Blue-green algae and their nitrogen fixation in the Baltic Sea in 1980, 1982, 1984. Finn. Mar. Res. 1989, 17, 3–20. [Google Scholar]
- Nausch, G.; Nehring, D.; Nagel, K. Nutrient Concentrations, Trends and Their Relation to Eutrophication; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Taucher, J.; Schulz, K.G.; Dittmar, T.; Sommer, U.; Oschlies, A.; Riebesell, U. Enhanced carbon overconsumption in response to increasing temperatures during a mesocosm experiment. Biogeosciences 2012, 9, 3531–3545. [Google Scholar] [CrossRef] [Green Version]
- Van de Waal, D.B.; Brandenburg, K.M.; Keuskamp, J.; Trimborn, S.; Rokitta, S.; Kranz, S.A.; Rost, B. Highest plasticity of carbon-concentrating mechanisms in earliest evolved phytoplankton. Limnol. Oceanogr. Lett. 2019, 4, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Ramos, J.B.E.; Biswas, H.; Schulz, K.G.; LaRoche, J.; Riebesell, U. Effect of rising atmospheric carbon dioxide on the marine nitrogen fixer Trichodesmium. Glob. Biogeochem. Cycles 2007, 21, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Hutchins, D.A.; Fu, F.X.; Zhang, Y.; Warner, M.E.; Feng, Y.; Portune, K.; Bernhardt, P.W.; Mulholland, M.R. CO2 control of Trichodesmium N-2 fixation, photosynthesis, growth rates, and elemental ratios: Implications for past, present, and future ocean biogeochemistry. Limnol. Oceanogr. 2007, 52, 1293–1304. [Google Scholar] [CrossRef] [Green Version]
- Kranz, S.A.; Sultemeyer, D.; Richter, K.U.; Rost, B. Carbon acquisition by Trichodesmium: The effect of pCO(2) and diurnal changes. Limnol. Oceanogr. 2009, 54, 548–559. [Google Scholar] [CrossRef]
- Tortell, P.D. Evolutionary and ecological perspectives on carbon acquisition in phytoplankton. Limnol. Oceanogr. 2000, 45, 744–750. [Google Scholar] [CrossRef]
- Wannicke, N.; Koch, B.P.; Voss, M. Release of fixed N-2 and C as dissolved compounds by Trichodesmium erythreum and Nodularia spumigena under the influence of high light and high nutrient (P). Aquat. Microb. Ecol. 2009, 57, 175–189. [Google Scholar] [CrossRef]
- Ploug, H.; Adam, B.; Musat, N.; Kalvelage, T.; Lavik, G.; Wolf-Gladrow, D.; Kuypers, M.M.M. Carbon, nitrogen and O-2 fluxes associated with the cyanobacterium Nodularia spumigena in the Baltic Sea. ISME J. 2011, 5, 1549–1558. [Google Scholar] [CrossRef] [Green Version]
- Litchman, E.; Klausmeier, C.A.; Schofield, O.M.; Falkowski, P.G. The role of functional traits and trade-offs in structuring phytoplankton communities: Scaling from cellular to ecosystem level. Ecol. Lett. 2007, 10, 1170–1181. [Google Scholar] [CrossRef]
- Wasmund, N.; Nausch, G.; Gerth, M.; Busch, S.; Burmeister, C.; Hansen, R.; Sadkowiak, B. Extension of the growing season of phytoplankton in the western Baltic Sea in response to climate change. Mar. Ecol. Prog. Ser. 2019, 622, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Tunin-Ley, A.; Ibanez, F.; Labat, J.P.; Zingone, A.; Lemee, R. Phytoplankton biodiversity and NW Mediterranean Sea warming: Changes in the dinoflagellate genus Ceratium in the 20th century. Mar. Ecol. Prog. Ser. 2009, 375, 85–99. [Google Scholar] [CrossRef]
- Hinder, S.L.; Hays, G.C.; Edwards, M.; Roberts, E.C.; Walne, A.W.; Gravenor, M.B. Changes in marine dinoflagellate and diatom abundance under climate change. Nat. Clim. Chang. 2012, 2, 271–275. [Google Scholar] [CrossRef]
- Dodge, J.D.; Marshall, H.G. Biogeographic analysis of the armoured planktonic dinoflagellate Ceratium in the North Atlantic and adjacent seas. J. Phycol. 1994, 30, 905–922. [Google Scholar] [CrossRef]
- O’Connor, M.I.; Piehler, M.F.; Leech, D.M.; Anton, A.; Bruno, J.F. Warming and Resource Availability Shift Food Web Structure and Metabolism. PLoS Biol. 2009, 7, 1–6. [Google Scholar] [CrossRef]
- Gaedke, U.; Ruhenstroth-Bauer, M.; Wiegand, I.; Tirok, K.; Aberle, N.; Breithaupt, P.; Lengfellner, K.; Wohlers, J.; Sommer, U. Biotic interactions may overrule direct climate effects on spring phytoplankton dynamics. Glob. Chang. Biol. 2010, 16, 1122–1136. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paul, C.; Sommer, U.; Matthiessen, B. Composition and Dominance of Edible and Inedible Phytoplankton Predict Responses of Baltic Sea Summer Communities to Elevated Temperature and CO2. Microorganisms 2021, 9, 2294. https://doi.org/10.3390/microorganisms9112294
Paul C, Sommer U, Matthiessen B. Composition and Dominance of Edible and Inedible Phytoplankton Predict Responses of Baltic Sea Summer Communities to Elevated Temperature and CO2. Microorganisms. 2021; 9(11):2294. https://doi.org/10.3390/microorganisms9112294
Chicago/Turabian StylePaul, Carolin, Ulrich Sommer, and Birte Matthiessen. 2021. "Composition and Dominance of Edible and Inedible Phytoplankton Predict Responses of Baltic Sea Summer Communities to Elevated Temperature and CO2" Microorganisms 9, no. 11: 2294. https://doi.org/10.3390/microorganisms9112294
APA StylePaul, C., Sommer, U., & Matthiessen, B. (2021). Composition and Dominance of Edible and Inedible Phytoplankton Predict Responses of Baltic Sea Summer Communities to Elevated Temperature and CO2. Microorganisms, 9(11), 2294. https://doi.org/10.3390/microorganisms9112294