
YgfY Contributes to Stress Tolerance in Shewanella oneidensis Neither as an Antitoxin Nor as a Flavinylation Factor of Succinate Dehydrogenase

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria and Growth Conditions
2.2. Construction of Strains and Plasmids
2.3. Overexpression of YgfY and YgfX in E. coli
2.4. Bacterial Two-Hybrid Assay
2.5. RNA Isolation and Sequencing
2.6. qRT-PCR Assay
2.7. Assay of Transcription Capability Using Click Chemistry
2.8. Assay of Nitrite Tolerance
2.9. Assay of Tolerance to Heat Shock
2.10. Quantification of Succinate
3. Results
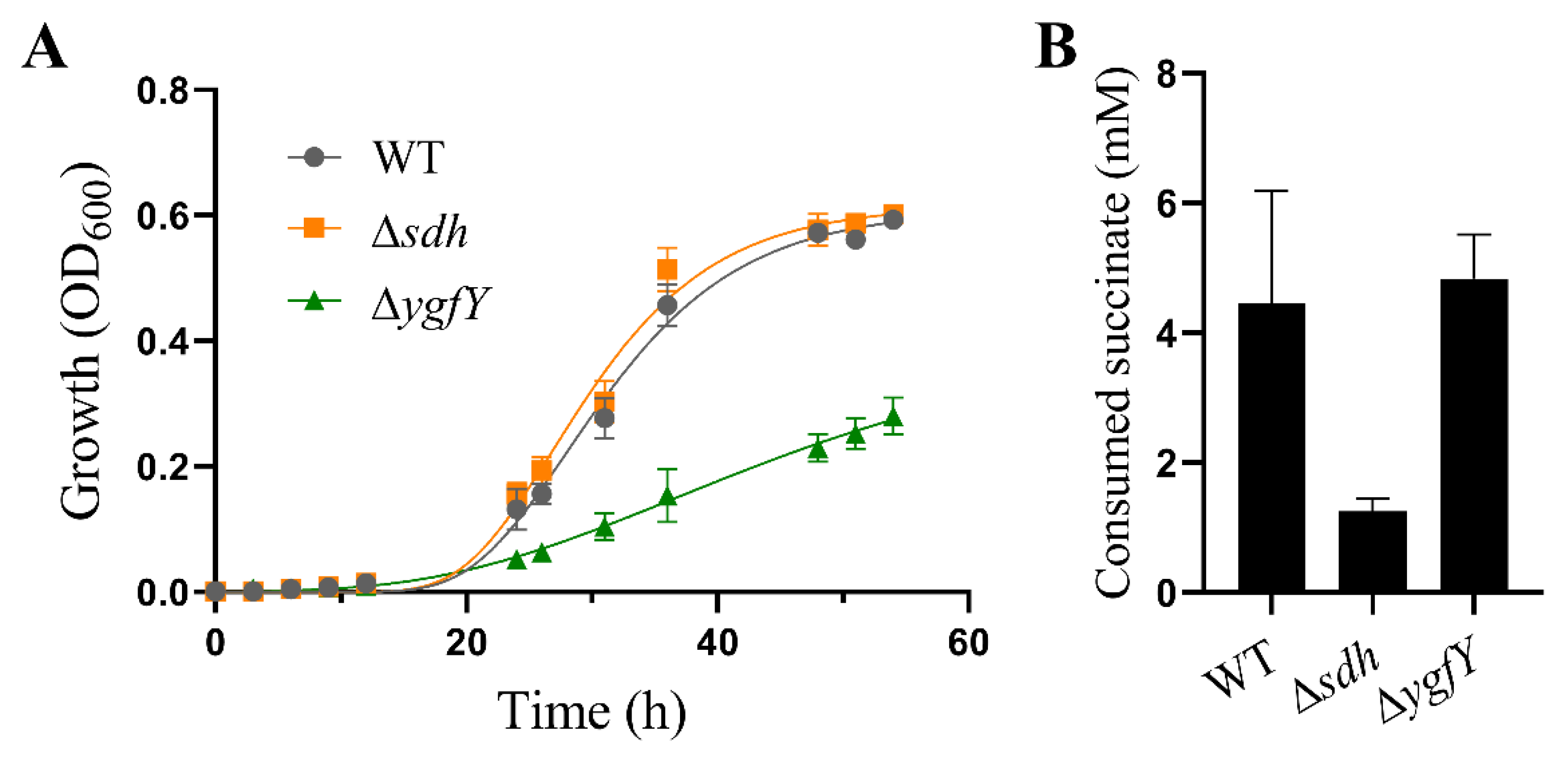
3.1. YgfY Is Required for Normal Growth of S. oneidensis MR-1
3.2. YgfY and YgfX Do Not Fulfil the Criteria of Type IV TA System
3.3. YgfY Does Not Attribute to Succinate Catabolism
3.4. YgfY Contributes to Capability of Transcription and Translation in S. oneidensis MR-1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Heidelberg, J.F.; Paulsen, I.T.; Nelson, K.E.; Gaidos, E.J.; Nelson, W.C.; Read, T.D.; Eisen, J.A.; Seshadri, R.; Ward, N.; Methe, B.; et al. Genome sequence of the dissimilatory metal ion-reducing bacterium Shewanella oneidensis. Nat. Biotechnol. 2002, 20, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Tiedje, J.M. Shewanella—The environmentally versatile genome. Nat. Biotechnol. 2002, 20, 1093–1094. [Google Scholar] [CrossRef]
- Li, B.B.; Cheng, Y.Y.; Fan, Y.Y.; Liu, D.F.; Fang, C.Y.; Wu, C.; Li, W.W.; Yang, Z.C.; Yu, H.Q. Estimates of abundance and diversity of Shewanella genus in natural and engineered aqueous environments with newly designed primers. Sci. Total Environ. 2018, 637–638, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Zhen, X.; Tang, K.; Liu, T.; Xu, X.; Chen, Z.; Guo, Y.; Liu, X.; Wood, T.K.; Ouyang, S.; et al. Novel polyadenylylation-dependent neutralization mechanism of the HEPN/MNT toxin/antitoxin system. Nucleic Acids Res. 2020, 48, 11054–11067. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, Targets, and Triggers: An Overview of Toxin-Antitoxin Biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; Tan, Q.; Awano, N.; Wu, K.P.; Inouye, M. YeeU enhances the bundling of cytoskeletal polymers of MreB and FtsZ, antagonizing the CbtA (YeeV) toxicity in Escherichia coli. Mol. Microbiol. 2012, 84, 979–989. [Google Scholar] [CrossRef]
- Masuda, H.; Tan, Q.; Awano, N.; Yamaguchi, Y.; Inouye, M. A novel membrane-bound toxin for cell division, CptA (YgfX), inhibits polymerization of cytoskeleton proteins, FtsZ and MreB, in Escherichia coli. FEMS Microbiol. Lett. 2012, 328, 174–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Z.; Wang, P.; Sun, C.; Guo, Y.; Wang, X. Interaction of Type IV Toxin/Antitoxin Systems in Cryptic Prophages of Escherichia coli K-12. Toxins 2017, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- McNeil, M.B.; Hampton, H.G.; Hards, K.J.; Watson, B.N.J.; Cook, G.M.; Fineran, P.C. The succinate dehydrogenase assembly factor, SdhE, is required for the flavinylation and activation of fumarate reductase in bacteria. FEBS Lett. 2014, 588, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.Y.; Wang, W.J.; Ding, S.T.; Zhang, M.X.; Tang, A.G.; Zhang, L.; Li, D.B.; Li, B.B.; Deng, G.Z.; Wu, C. Pyruvate accelerates palladium reduction by regulating catabolism and electron transfer pathway in Shewanella oneidensis. Appl. Environ. Microbiol. 2021, 87, e02716-20. [Google Scholar] [CrossRef]
- Cheng, Y.Y.; Wu, C.; Wu, J.Y.; Jia, H.L.; Wang, M.Y.; Wang, H.Y.; Zou, S.M.; Sun, R.R.; Jia, R.; Xiao, Y.Z. FlrA Represses Transcription of the Biofilm-Associated bpfA Operon in Shewanella putrefaciens. Appl. Environ. Microbiol. 2017, 83, e02410-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Cheng, Y.Y.; Yin, H.; Song, X.N.; Li, W.W.; Zhou, X.X.; Zhao, L.P.; Tian, L.J.; Han, J.C.; Yu, H.Q. Oxygen promotes biofilm formation of Shewanella putrefaciens CN32 through a diguanylate cyclase and an adhesin. Sci. Rep. 2013, 3, 1945. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Chan, D.; Guo, X.; Chang, F.; Chen, M.; Wang, Q.; Song, X.; Wu, C. Hydrogen production driven by formate oxidation in Shewanella oneidensis MR-1. Appl. Microbiol. Biotechnol. 2020, 104, 5579–5591. [Google Scholar] [CrossRef] [PubMed]
- Jao, C.Y.; Salic, A. Exploring RNA transcription and turnover in vivo by using click chemistry. Proc. Natl. Acad. Sci. USA 2008, 105, 15779–15784. [Google Scholar] [CrossRef] [Green Version]
- Edwards, R.A.; Keller, L.H.; Schifferli, D.M. Improved allelic exchange vectors and their use to analyze 987P fimbria gene expression. Gene 1998, 207, 149–157. [Google Scholar] [CrossRef]
- Dehio, C.; Meyer, M. Maintenance of broad-host-range incompatibility group P and group Q plasmids and transposition of Tn5 in Bartonella henselae following conjugal plasmid transfer from Escherichia coli. J. Bacteriol. 1997, 179, 538–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995, 177, 4121–4130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez-Laslop, N.; Lee, H.; Neyfakh, A.A. Increased persistence in Escherichia coli caused by controlled expression of toxins or other unrelated proteins. J. Bacteriol. 2006, 188, 3494–3497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeil, M.B.; Fineran, P.C. The Conserved RGxxE Motif of the Bacterial FAD Assembly Factor SdhE Is Required for Succinate Dehydrogenase Flavinylation and Activity. Biochemistry 2013, 52, 7628–7640. [Google Scholar] [CrossRef]
- Yang, Y.; McCue, L.A.; Parsons, A.B.; Feng, S.; Zhou, J. The tricarboxylic acid cycle in Shewanella oneidensis is independent of Fur and RyhB control. BMC Microbiol. 2010, 10, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, H.X.; Khalimonchuk, O.; Schraders, M.; Dephoure, N.; Bayley, J.P.; Kunst, H.; Devilee, P.; Cremers, C.W.; Schiffman, J.D.; Bentz, B.G.; et al. SDH5, a gene required for flavination of succinate dehydrogenase, is mutated in paraganglioma. Science 2009, 325, 1139–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, L.; Zengel, J.M. Expression of ribosomal genes in bacteria. Adv. Genet. 1982, 21, 53–121. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; Dwyer, D.J.; Wierzbowski, J.; Cottarel, G.; Collins, J.J. Mistranslation of membrane proteins and two-component system activation trigger antibiotic-mediated cell death. Cell 2008, 135, 679–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, J.; Cho, C.; Guo, L.T.; Aerni, H.R.; Rinehart, J.; Soll, D. Protein aggregation caused by aminoglycoside action is prevented by a hydrogen peroxide scavenger. Mol. Cell 2012, 48, 713–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, G.; Hong, Y.; Drlica, K.; Zhao, X. Suppression of Reactive Oxygen Species Accumulation Accounts for Paradoxical Bacterial Survival at High Quinolone Concentration. Antimicrob. Agents Chemother. 2018, 62, e01622-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response: Life on the verge of death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, B.; Kemeny, G.; Switzer, R.C.; Hamilton, T.C. Quantitative evidence for protein denaturation as the cause of thermal death. Nature 1971, 232, 471–473. [Google Scholar] [CrossRef] [PubMed]
- ElBanna, S.A.; Moneib, N.A.; Aziz, R.K.; Samir, R. Genomics-guided identification of a conserved CptBA-like toxin-antitoxin system in Acinetobacter baumannii. J. Adv. Res. 2021, 30, 159–170. [Google Scholar] [CrossRef]
- McNeil, M.B.; Iglesias-Cans, M.C.; Clulow, J.S.; Fineran, P.C. YgfX (CptA) is a multimeric membrane protein that interacts with the succinate dehydrogenase assembly factor SdhE (YgfY). Microbiology 2013, 159, 1352–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcneil, M.B.; Clulow, J.S.; Wilf, N.M.; Salmond, G.P.C.; Fineran, P.C. SdhE Is a Conserved Protein Required for Flavinylation of Succinate Dehydrogenase in Bacteria. J. Biol. Chem. 2012, 287, 18418–18428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiefler, I.; Bringer, S.; Bott, M. SdhE-dependent formation of a functional Acetobacter pasteurianus succinate dehydrogenase in Gluconobacter oxydans—A first step toward a complete tricarboxylic acid cycle. Appl. Microbiol. Biot. 2015, 99, 9147–9160. [Google Scholar] [CrossRef] [PubMed]
- Maher, M.J.; Herath, A.S.; Udagedara, S.R.; Dougan, D.A.; Truscott, K.N. Crystal structure of bacterial succinate:quinone oxidoreductase flavoprotein SdhA in complex with its assembly factor SdhE. Proc. Natl. Acad. Sci. USA 2018, 115, 2982–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Maklashina, E.; Cecchini, G.; Iverson, T.M. The roles of SDHAF2 and dicarboxylate in covalent flavinylation of SDHA, the human complex II flavoprotein. Proc. Natl. Acad. Sci. USA 2020, 117, 23548–23556. [Google Scholar] [CrossRef]
- Sharma, P.; Maklashina, E.; Cecchini, G.; Iverson, T.M. Crystal structure of an assembly intermediate of respiratory Complex II. Nat. Commun. 2018, 9, 274. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.-X.; Zheng, K.-L.; Tang, A.-G.; Hu, X.-X.; Guo, X.-X.; Wu, C.; Cheng, Y.-Y. YgfY Contributes to Stress Tolerance in Shewanella oneidensis Neither as an Antitoxin Nor as a Flavinylation Factor of Succinate Dehydrogenase. Microorganisms 2021, 9, 2316. https://doi.org/10.3390/microorganisms9112316
Zhang M-X, Zheng K-L, Tang A-G, Hu X-X, Guo X-X, Wu C, Cheng Y-Y. YgfY Contributes to Stress Tolerance in Shewanella oneidensis Neither as an Antitoxin Nor as a Flavinylation Factor of Succinate Dehydrogenase. Microorganisms. 2021; 9(11):2316. https://doi.org/10.3390/microorganisms9112316
Chicago/Turabian StyleZhang, Ming-Xing, Kai-Li Zheng, Ai-Guo Tang, Xiao-Xia Hu, Xin-Xin Guo, Chao Wu, and Yuan-Yuan Cheng. 2021. "YgfY Contributes to Stress Tolerance in Shewanella oneidensis Neither as an Antitoxin Nor as a Flavinylation Factor of Succinate Dehydrogenase" Microorganisms 9, no. 11: 2316. https://doi.org/10.3390/microorganisms9112316