
Bacterial Morphotypes as Important Trait for Uropathogenic E. coli Diagnostic; a Virulence-Phenotype-Phylogeny Study

 ,
,  , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Urine Samples Collection
2.2. Urinalysis and Detection of UPEC Morphotypes in Urine Sediment
2.3. Urine Cultures and Biochemical Identification of Obtained Bacterial Isolates
2.4. DNA Extraction
2.5. Molecular Identification of E. coli
2.6. Identification of Phylogenetic Groups
2.7. Genotypic Characterization of UPEC Isolates
2.8. Phenotypic Characterization of UPEC Isolates
2.8.1. Motility Test
2.8.2. α-Hemolysin Production
2.8.3. Biofilm Formation Assay
2.8.4. Capsule Production
2.8.5. Adherence Assay
2.8.6. Antibiotic Resistance
2.9. Statistical Analysis
2.10. Ethic Statements
3. Results
3.1. Clinical Characteristics of Adult Women with UTI
3.2. Urine Cultures
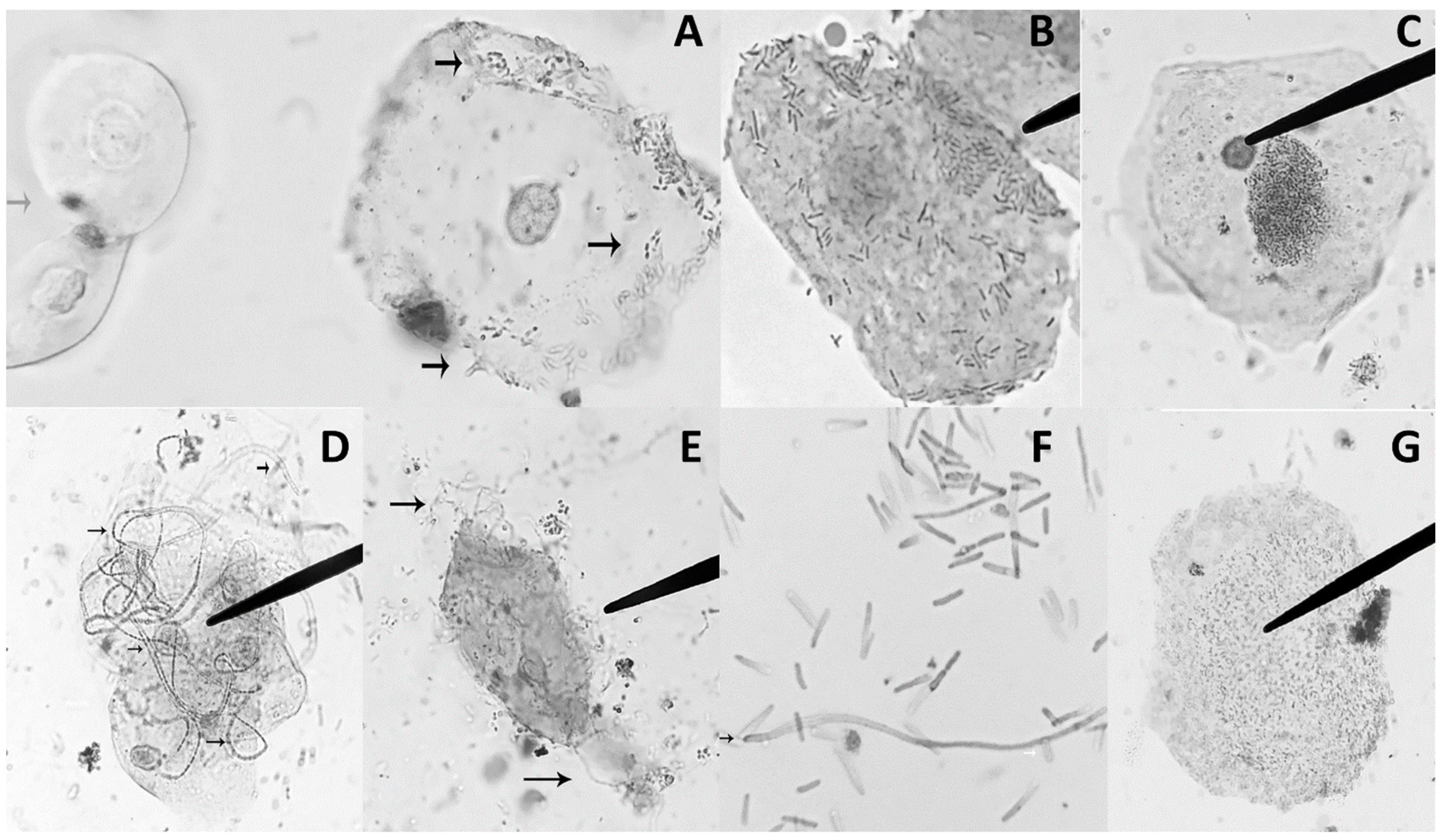
3.3. UPEC Morphotypes in Urine Sediment of Patients with UTI
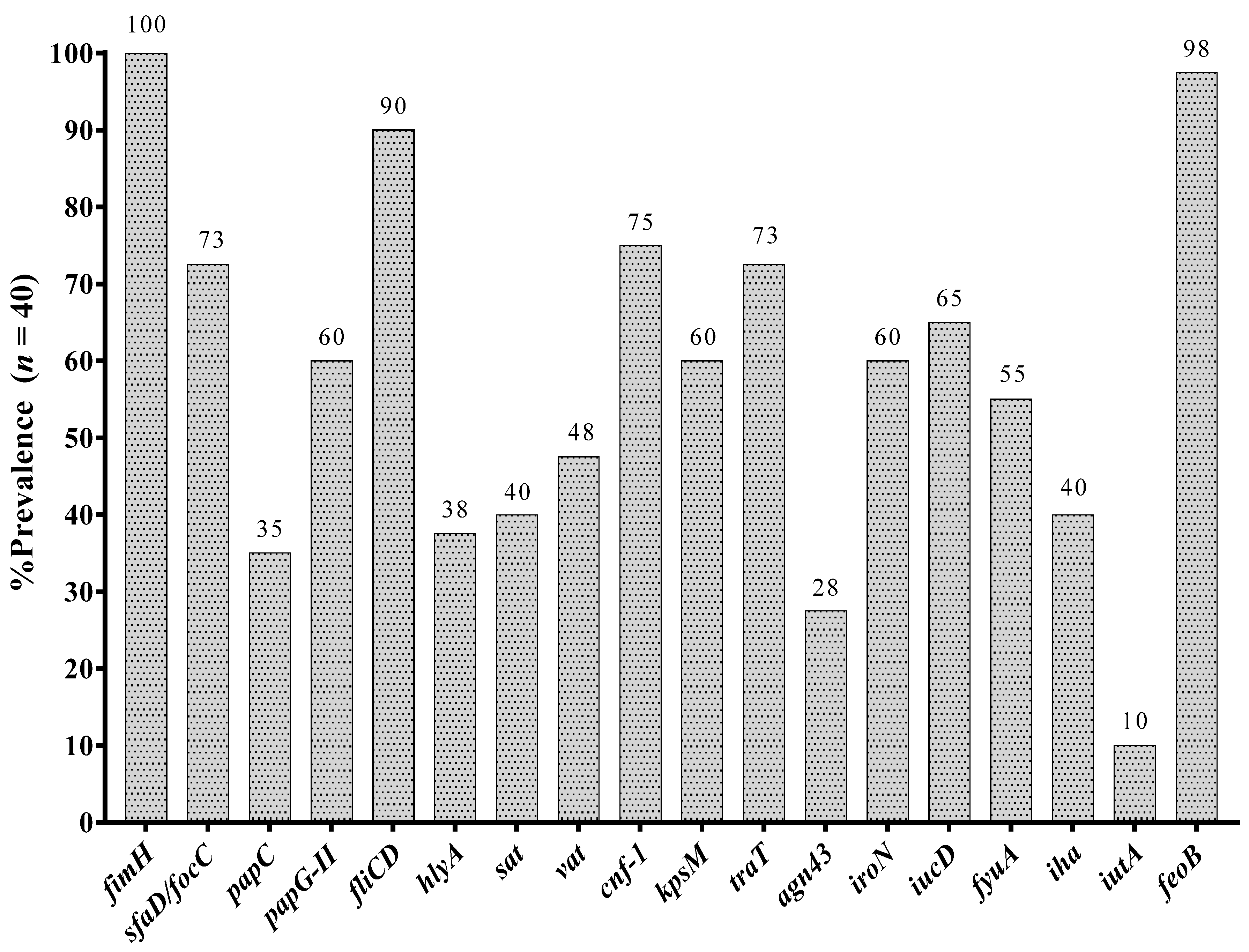
3.4. Prevalence of Virulence Associated Genes
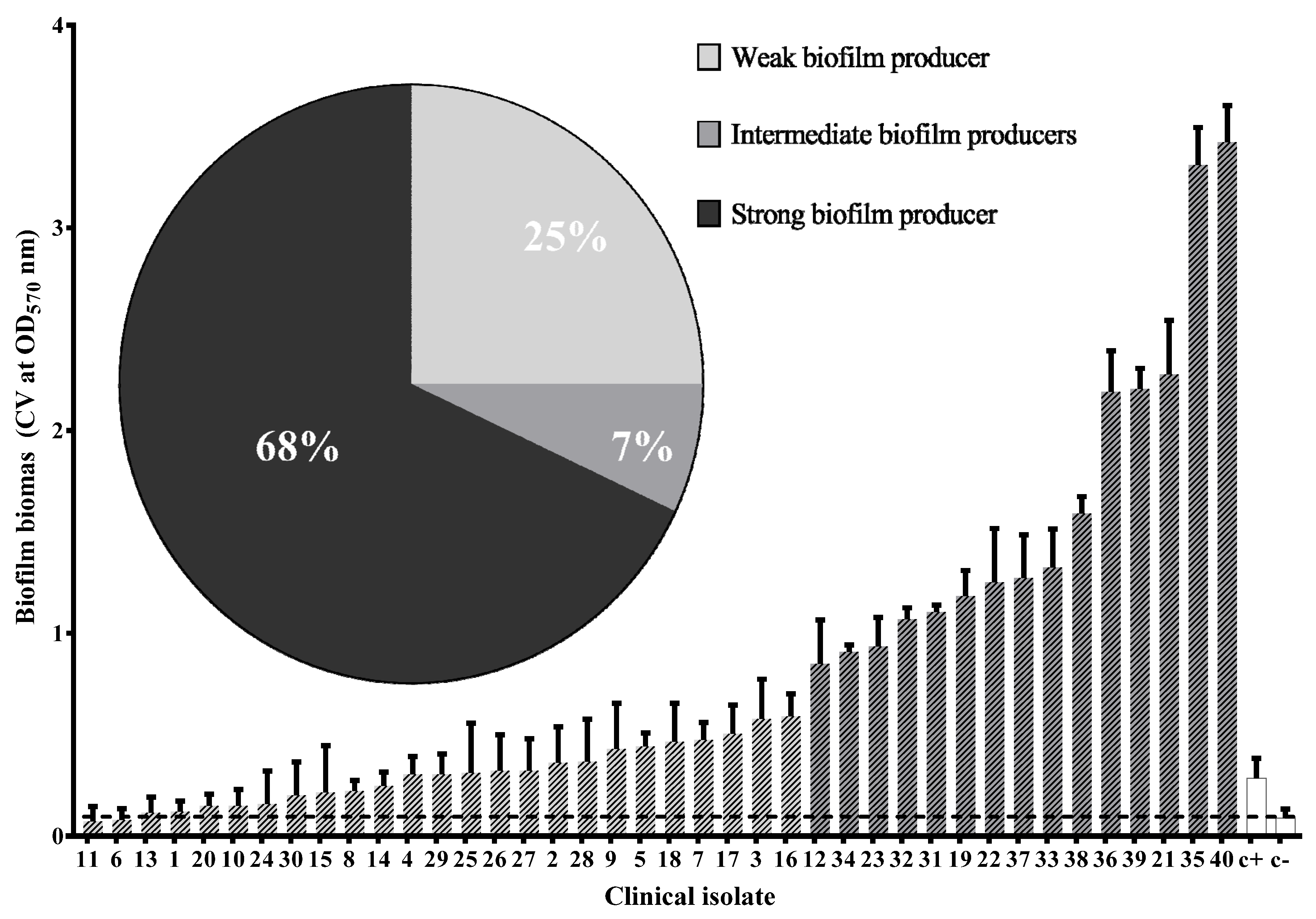
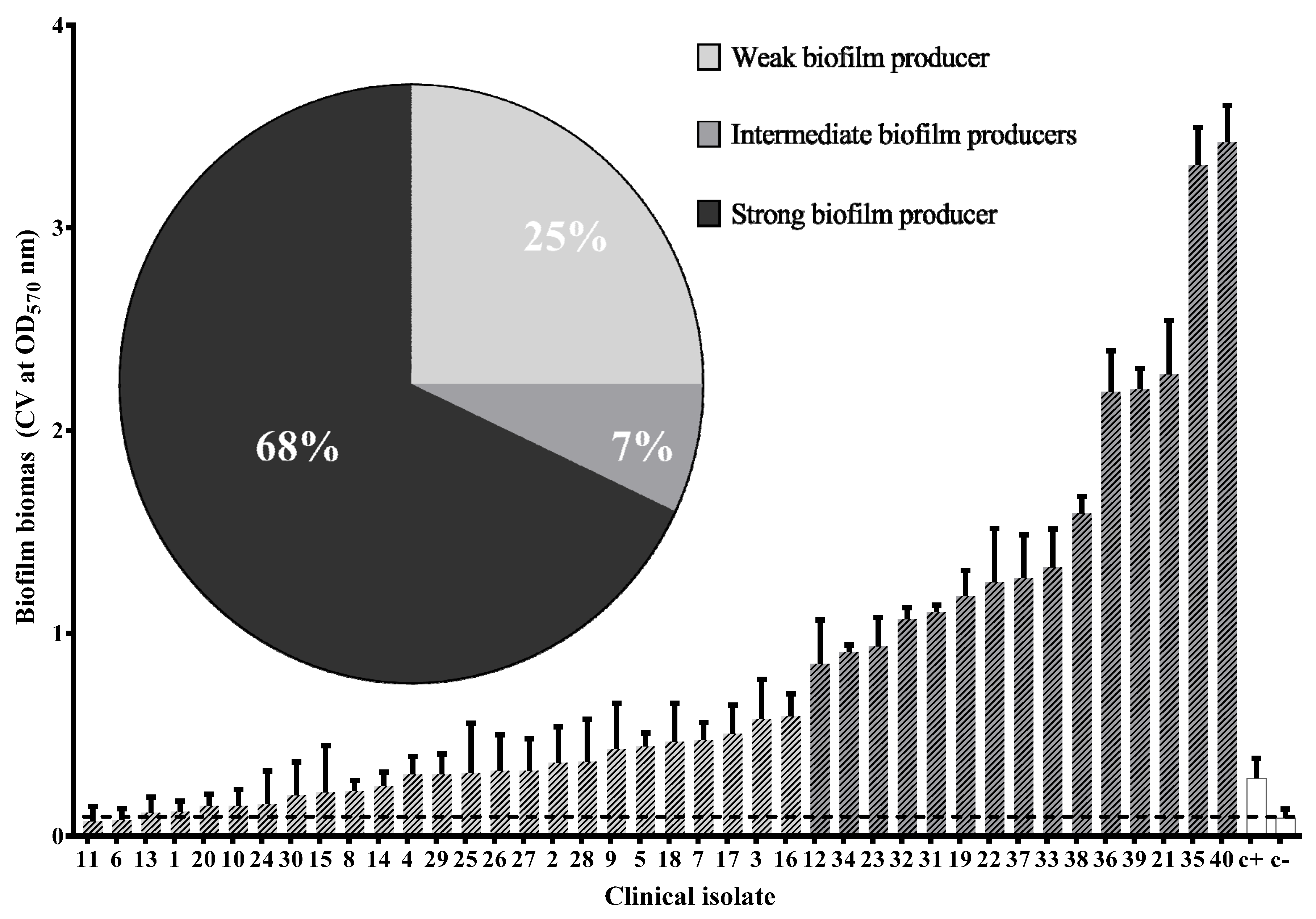
3.5. Virulence Phenotypes of Clinical Isolates of UPEC
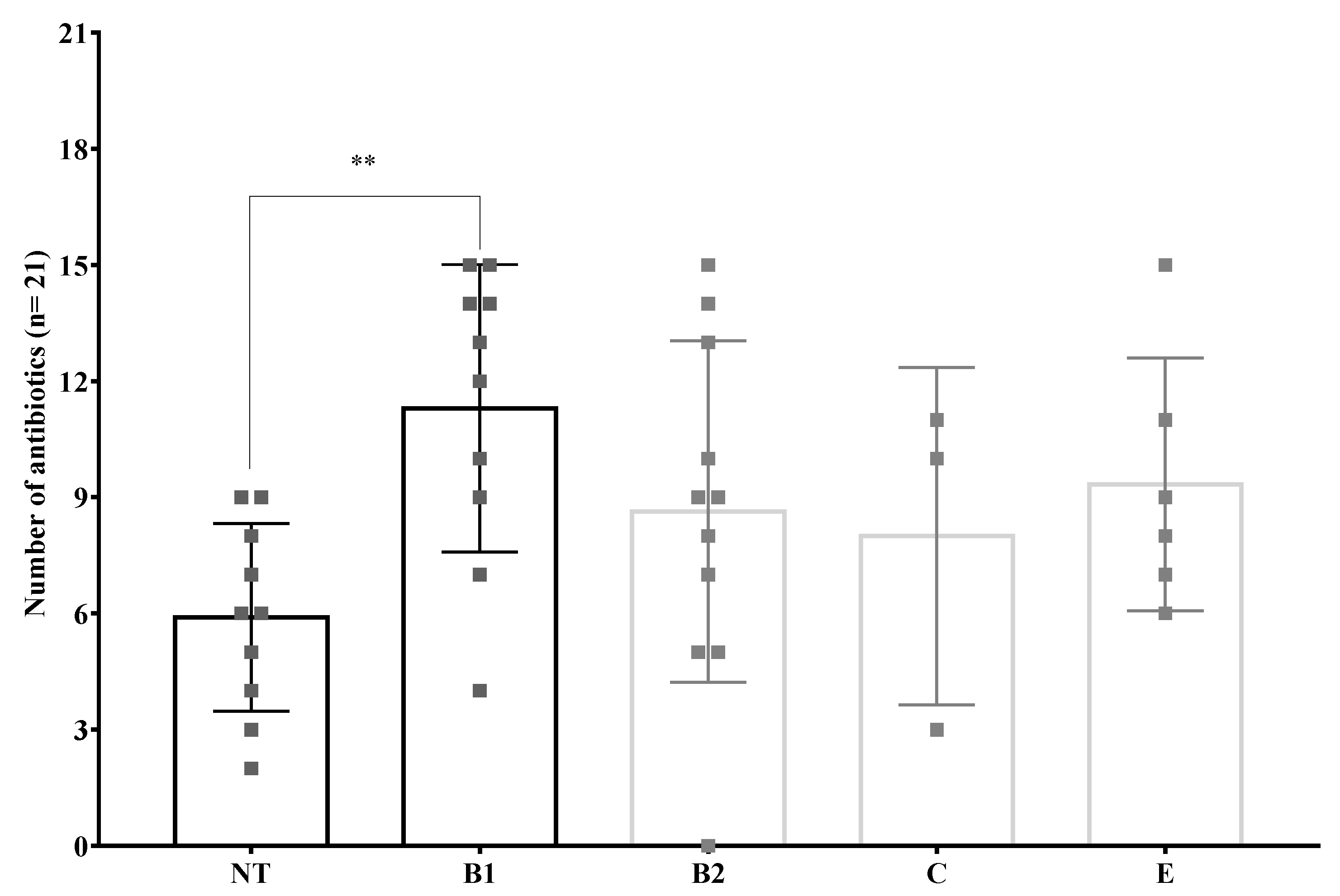
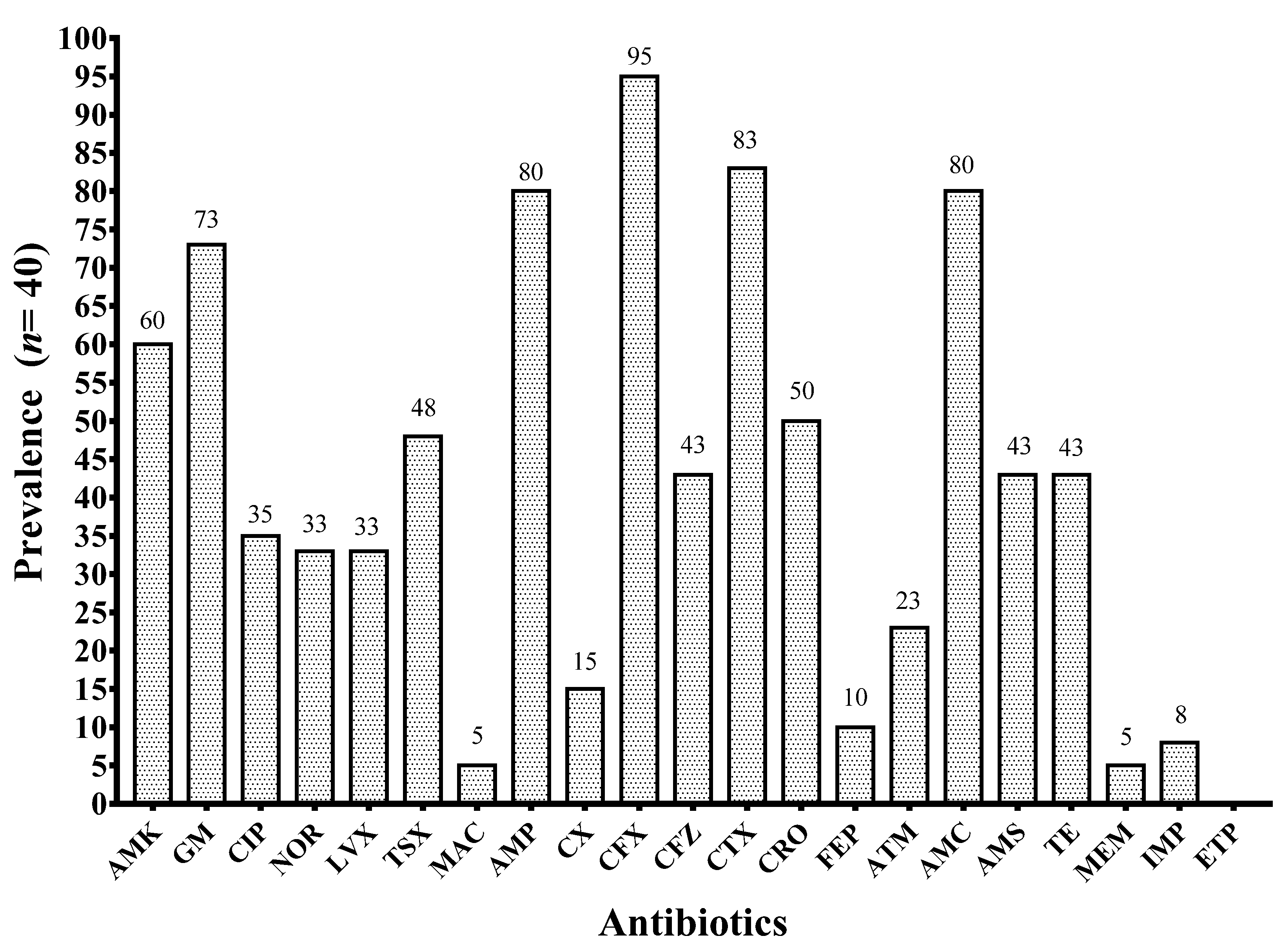
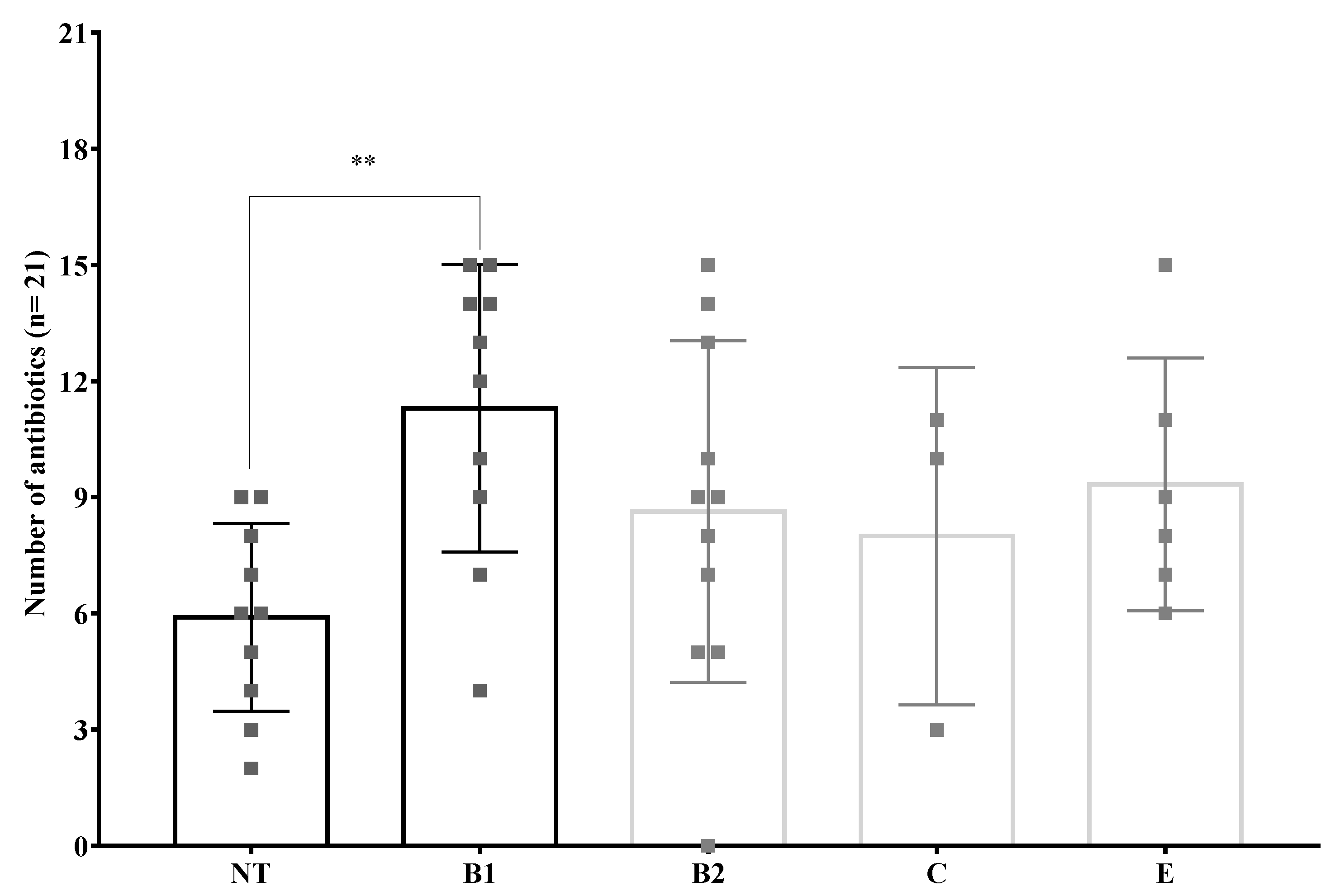
3.6. Antibiotic Resistance Phenotypes
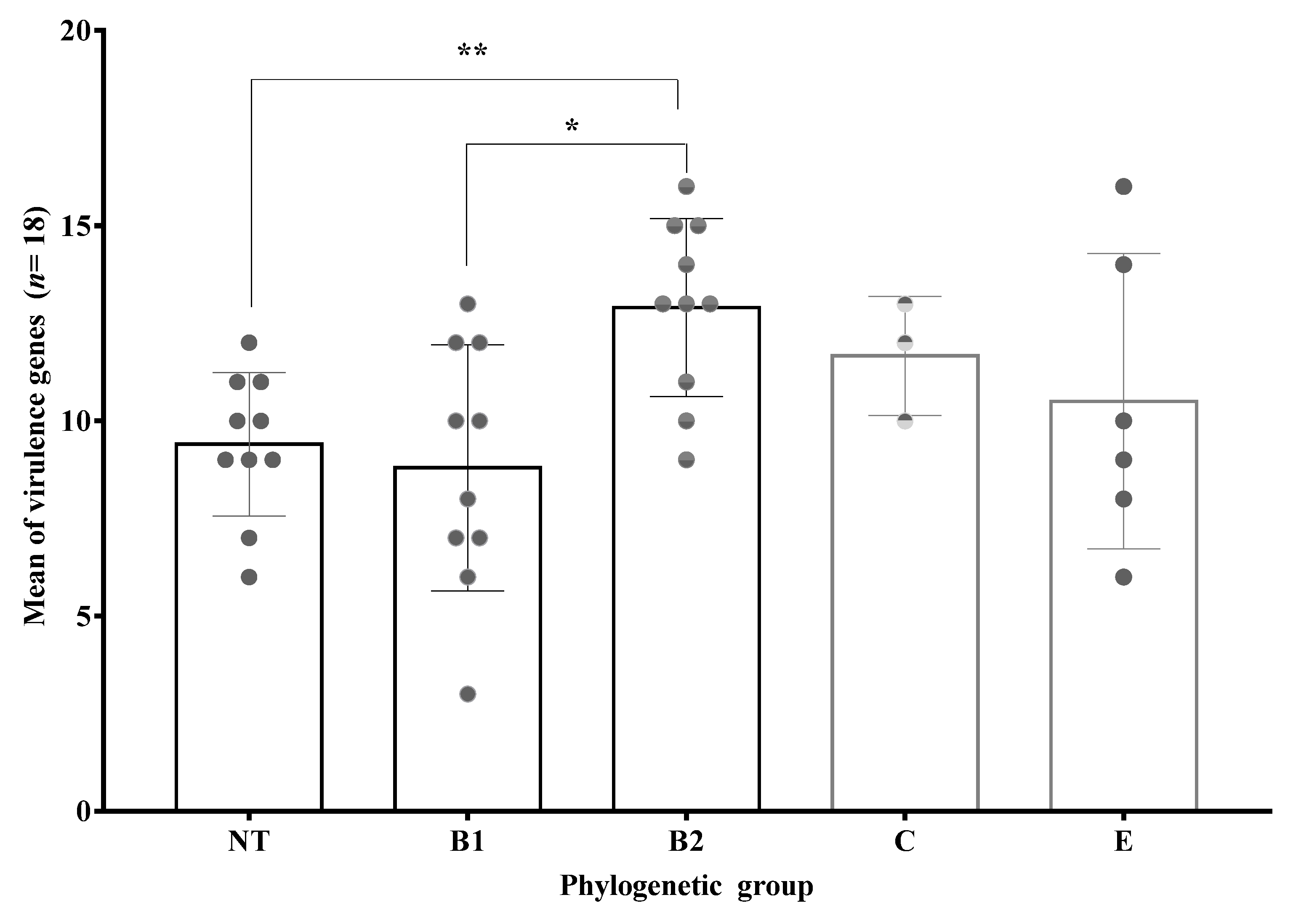
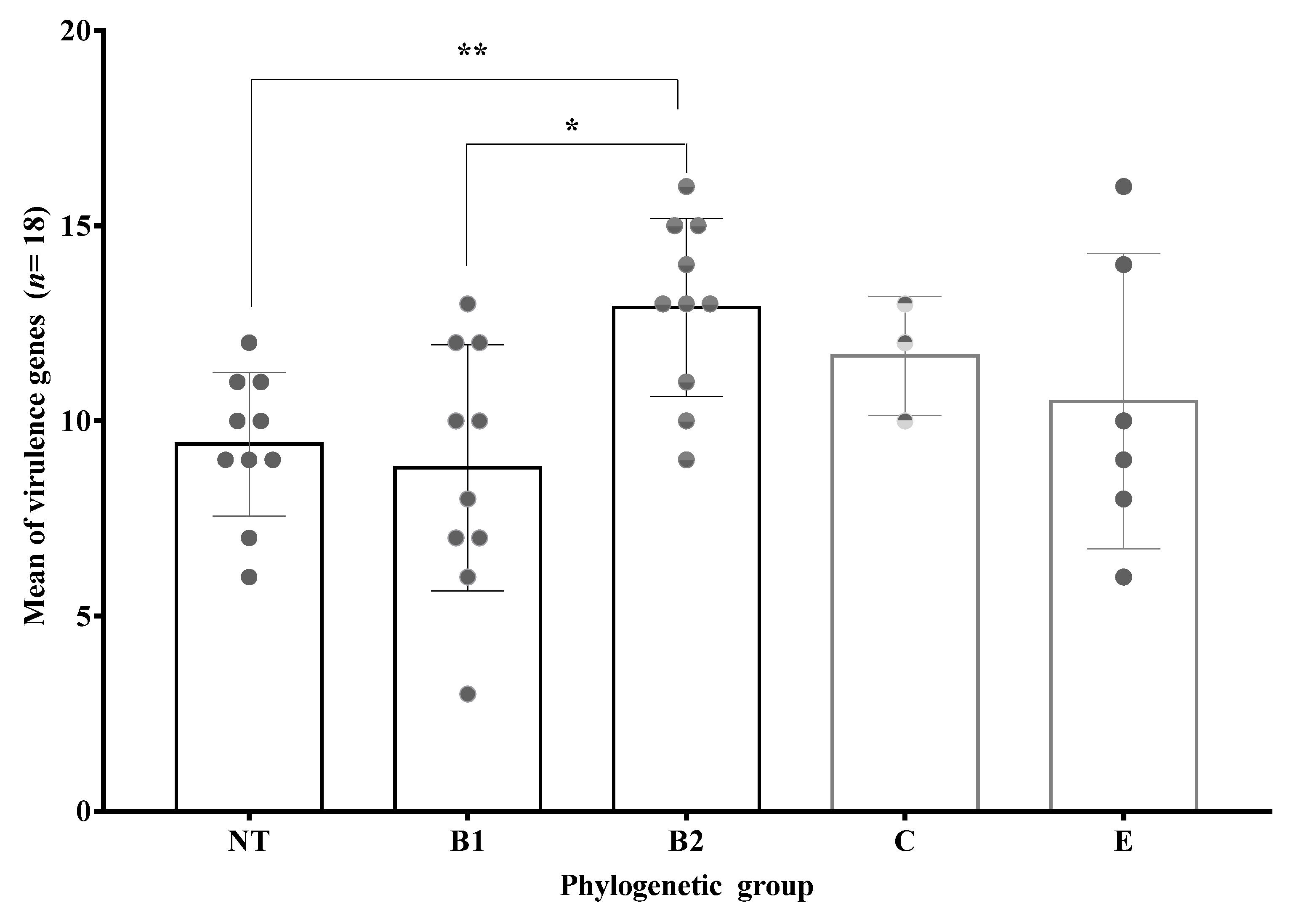
3.7. Phylogenetic Groups of Obtained Clinical Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Secretaría de Salud Boletin de Semana Epidemiológica 53. Available online: https://www.gob.mx/salud/acciones-y-programas/historico-boletin-epidemiologico (accessed on 12 January 2021).
- Secretaría de Salud Boletin de Semana Epidemiológica 52. Available online: https://www.gob.mx/salud/acciones-y-programas/historico-boletin-epidemiologico (accessed on 12 January 2021).
- Foxman, B. The epidemiology of urinary tract infection. Nat. Rev. Urol. 2010, 7, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, Y.; Minamino, T. Structure and Function of the Bi-Directional Bacterial Flagellar Motor. Biomolecules 2014, 4, 217–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, J.A.; Marklund, B.I.; Ilver, D.; Haslam, D.; Kaack, M.B.; Baskin, G.; Louis, M.; Mollby, R.; Winberg, J.; Normark, S. The Gal(alpha 1-4)Gal-specific tip adhesin of Escherichia coli P-fimbriae is needed for pyelonephritis to occur in the normal urinary tract. Proc. Natl. Acad. Sci. USA 1994, 91, 11889–11893. [Google Scholar] [CrossRef] [Green Version]
- Livorsi, D.J.; Stenehjem, E.; Stephens, D.S. Virulence Factors of Gram- Negative Bacteria in Sepsis with a Focus on Neisseria meningitidis. Contrib Microbiol 2011, 17, 31–47. [Google Scholar]
- González, M.J.; Da Cunda, P.; Notejane, M.; Zunino, P.; Scavone, P.; Robino, L. Fosfomycin tromethamine activity on biofilm and intracellular bacterial communities produced by uropathogenic Escherichia coli isolated from patients with urinary tract infection. Pathog. Dis. 2019, 77, ftz022. [Google Scholar] [CrossRef]
- Olson, P.; Hunstad, D. Subversion of Host Innate Immunity by Uropathogenic Escherichia coli. Pathogens 2016, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Robino, L.; Scavone, P.; Araujo, L.; Algorta, G.; Zunino, P.; Pírez, M.C.; Vignoli, R. Intracellular bacteria in the pathogenesis of escherichia coli urinary tract infection in children. Clin. Infect. Dis. 2014, 59, e158–e164. [Google Scholar] [CrossRef] [Green Version]
- Rosen, D.A.; Hooton, T.M.; Stamm, W.E.; Humphrey, P.A.; Hultgren, S.J. Detection of intracellular bacterial communities in human urinary tract infection. PLoS Med. 2007, 4, 1949–1958. [Google Scholar] [CrossRef]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Monrreal, M.G.; Arenas-Hernández, M.M.P.; Enciso-Martínez, Y.; Martinez de la Peña, C.F.; Rocha-Gracia, R.d.C.; Lozano-Zarain, P.; Navarro-Ocaña, A.; Martínez-Laguna, Y.; de la Rosa-López, R. Virulence and Resistance Determinants of Uropathogenic Escherichia coli Strains Isolated from Pregnant and Non-Pregnant Women from Two States in Mexico. Infect. Drug Resist. 2020, 13, 295–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda-Estrada, L.I.; Ruíz-Rosas, M.; Molina-López, J.; Parra-Rojas, I.; González-Villalobos, E.; Castro-Alarcón, N. Relación entre factores de virulencia, resistencia a antibióticos y los grupos filogenéticos de Escherichia coli uropatógena en dos localidades de México. Enferm. Infecc. Microbiol. Clin. 2017, 35, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Iranpour, D.; Hassanpour, M.; Ansari, H.; Tajbakhsh, S.; Khamisipour, G.; Najafi, A. Phylogenetic Groups of Escherichia coli Strains from Patients with Urinary Tract Infection in Iran Based on the New Clermont Phylotyping Method. Biomed Res. Int. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Figueroa, C.; Cortés-Sarabia, K.; Del Carmen Alarcón-Romero, L.; Catalán-Nájera, H.G.; Martínez-Alarcón, M.; Vences-Velázquez, A. Observation of intracellular bacterial communities in urinary sediment using brightfield microscopy; A case report. BMC Urol. 2020. [Google Scholar] [CrossRef]
- KASS, E.H. Pyelonephritis and Bacteriuria. Ann. Intern. Med. 1962, 56, 46. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor: New York, NY, USA, 2012; Volume 33, ISBN 9781936113415. [Google Scholar]
- Walker, D.I.; McQuillan, J.; Taiwo, M.; Parks, R.; Stenton, C.A.; Morgan, H.; Mowlem, M.C.; Lees, D.N. A highly specific Escherichia coli qPCR and its comparison with existing methods for environmental waters. Water Res. 2017, 126, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: A quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Luna, V.A.; Peak, K.K.; Veguilla, W.O.; Reeves, F.; Heberlein-Larson, L.; Cannons, A.C.; Amuso, P.; Cattani, J. Use of Two Selective Media and a Broth Motility Test Can Aid in Identification or Exclusion of Bacillus anthracis. J. Clin. Microbiol. 2005, 43, 4336–4341. [Google Scholar] [CrossRef] [Green Version]
- Anthony, E.E. a Note on Capsule Staining. Science 1931, 73, 319–320. [Google Scholar] [CrossRef]
- Barrios-Villa, E.; Cortés-Cortés, G.; Lozano-Zaraín, P.; de la Paz Arenas-Hernández, M.M.; Martínez de la Peña, C.F.; Martínez-Laguna, Y.; Torres, C.; Rocha-Gracia, R.D.C. Adherent/invasive Escherichia coli (AIEC) isolates from asymptomatic people: New E. coli ST131 O25:H4/H30-Rx virotypes. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 42. [Google Scholar] [CrossRef] [Green Version]
- Vollmerhausen, T.L.; Woods, J.L.; Faoagali, J.; Katouli, M. Interactions of uroseptic Escherichia coli with renal (A-498) and gastrointestinal (HT-29) cell lines. J. Med. Microbiol. 2014, 63, 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F. Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Microbiology 2011, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Patra, N.; Prakash, M.R.; Patil, S.; Rao, M.R. First Case Report of Surgical Site Infection Due to Buttiauxella agrestis in a Neurocare Center in India. Arch. Med. Health Sci. 2018, 6, 117–119. [Google Scholar] [CrossRef]
- Antonello, V.S.; Dallé, J.; Domingues, G.C.; Ferreira, J.A.S.; Fontoura, M.d.C.Q.; Knapp, F.B. Post-cesarean surgical site infection due to Buttiauxella agrestis. Int. J. Infect. Dis. 2014, 22, 65–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardentey-Reyes, A.; Jacobs, F.; Struelens, M.J.; Rodriguez-Villalobos, H. First case of bacteremia caused by moellerella wisconsensis: Case report and a review of the literature. Infection 2009, 37, 544–546. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.R.; Hux, J.E. Quantifying the risk of infectious diseases for people with diabetes. Diabetes Care 2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, L.M.A.J.; Gorter, K.J.; Hak, E.; Goudzwaard, W.L.; Schellevis, F.G.; Hoepelman, A.I.M.; Rutten, G.E.H.M. Increased Risk of Common Infections in Patients with Type 1 and Type 2 Diabetes Mellitus. Clin. Infect. Dis. 2005, 26, 510–513. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.H.; Cho, J.H.; Lee, J.H.; Park, Y.J.; Jin, S.; Park, G.Y.; Kim, J.S.; Kang, Y.J.; Kwon, O.; Choi, J.Y.; et al. Risk factors for recurrent urinary tract infection in kidney transplant recipients. Transplant. Pro. 2013, 45, 1584–1589. [Google Scholar]
- Rogers, G.B.; Hoffman, L.R.; Whiteley, M.; Daniels, T.W.V.; Carroll, M.P.; Bruce, K.D. Revealing the dynamics of polymicrobial infections: Implications for antibiotic therapy. Trends Microbiol. 2010, 18, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Croxall, G.; Weston, V.; Joseph, S.; Manning, G.; Cheetham, P.; McNally, A. Increased human pathogenic potential of Escherichia coli from polymicrobial urinary tract infections in comparison to isolates from monomicrobial culture samples. J. Med. Microbiol. 2011, 60, 102–109. [Google Scholar] [CrossRef]
- Ranjan, K.P.; Ranjan, N. Citrobacter: An emerging health care associated urinary pathogen. Urol. Ann. 2013, 5, 313–314. [Google Scholar] [PubMed]
- Sami, H.; Sultan, A.; Rizvi, M.; Khan, F.; Ahmad, S.; Shukla, I.; Khan, H. Citrobacter as a uropathogen, its prevalence and antibiotics susceptibility pattern. CHRISMED J. Heal. Res. 2017, 4, 23. [Google Scholar] [CrossRef]
- Aller, A.I.; Castro, C.; Medina, M.J.; González, M.T.; Sevilla, P.; Morilla, M.D.; Corzo, J.E.; Martín-Mazuelos, E. Isolation of Moellerella wisconsensis from blood culture from a patient with acute cholecystitis. Clin. Microbiol. Infect. 2009, 15, 1193–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, S.; Bezdan, D.; Oberhettinger, P.; Vogel, W.; Dörfel, D.; Dick, J.; Marschal, M.; Liese, J.; Weidenmaier, C.; Autenrieth, I.; et al. Whole-genome sequencing enabling the detection of a colistin-resistant hypermutating Citrobacter werkmanii strain harbouring a novel metallo-β-lactamase VIM-48. Int. J. Antimicrob. Agents 2018, 51, 867–874. [Google Scholar] [CrossRef]
- Duman, M.; Saticioglu, I.B.; Buyukekiz, A.G.; Balta, F.; Altun, S. Molecular characterization and antimicrobial resistance profile of atypical Citrobacter gillenii and Citrobacter sp. isolated from diseased rainbow trout (Oncorhynchus mykiss). J. Glob. Antimicrob. Resist. 2017, 10, 136–142. [Google Scholar] [CrossRef]
- Robino, L.; Scavone, P.; Araujo, L.; Algorta, G.; Zunino, P.; Vignoli, R. Detection of intracellular bacterial communities in a child with Escherichia coli recurrent urinary tract infections. Pathog. Dis. 2013, 68, 78–81. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Castillo, F.Y.; Moreno-Flores, A.C.; Avelar-González, F.J.; Márquez-Díaz, F.; Harel, J.; Guerrero-Barrera, A.L. An evaluation of multidrug-resistant Escherichia coli isolates in urinary tract infections from Aguascalientes, Mexico: Cross-sectional study. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 34. [Google Scholar] [CrossRef]
- Paniagua-Contreras, G.L.; Monroy-Pérez, E.; Díaz-Velásquez, C.E.; Uribe-García, A.; Labastida, A.; Peñaloza-Figueroa, F.; Domínguez-Trejo, P.; García, L.R.; Vaca-Paniagua, F.; Vaca, S. Whole-genome sequence analysis of multidrug-resistant uropathogenic strains of Escherichia coli from Mexico. Infect. Drug Resist. 2019, 12, 2363–2377. [Google Scholar] [CrossRef] [Green Version]
- Chávez-Jacobo, V.; Ramírez-Díaz, M.; Silva-Sánchez, J.; Cervantes, C. Resistencia Bacteriana a Quinolonas: Determinantes Codificados en Plásmidos. REB. Rev. Educ. bioquímica 2015. [Google Scholar]
- Guajardo-Lara, C.E.; González-Martínez, P.M.; Ayala-Gaytán, J.J. Resistencia antimicrobiana en la infección urinaria por Escherichia coli adquirida en la comunidad: ¿Cuál antibiótico voy a usar? Salud Publica Mex. 2009, 51, 157–161. [Google Scholar] [CrossRef]
- Garza-González, E.; Bocanegra-Ibarias, P.; Bobadilla-del-Valle, M.; Ponce-de-León-Garduño, L.A.; Esteban-Kenel, V.; Silva-Sánchez, J.; Garza-Ramos, U.; Barrios-Camacho, H.; López-Jácome, L.E.; Colin-Castro, C.A.; et al. Drug resistance phenotypes and genotypes in Mexico in representative gram-negative species: Results from the infivar network. PLoS One 2021, 16, e0248614. [Google Scholar] [CrossRef] [PubMed]
- Matta-Chuquisapon, J.; Valencia-Bazalar, E.; Marocho-Chahuayo, L.; Gonzales-Escalante, E.; Sevilla-Andrade, C.R. Presencia de genes fimH y afa en aislamientos urinarios de Escherichia coli productora de betalactamasas de espectro extendido en Lima, Perú. Rev. Peru. Med. Exp. Salud Publica 2020, 37, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Dadi, B.R.; Abebe, T.; Zhang, L.; Mihret, A.; Abebe, W.; Amogne, W. Distribution of virulence genes and phylogenetics of uropathogenic Escherichia coli among urinary tract infection patients in Addis Ababa, Ethiopia. BMC Infect. Dis. 2020, 20, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Morales-Espinosa, R.; Hernandez-Castro, R.; Delgado, G.; Mendez, J.L.; Navarro, A.; Manjarrez, A.; Cravioto, A. UPEC strain characterization isolated from Mexican patients with recurrent urinary infections. J. Infect. Dev. Ctries. 2016, 10, 317–328. [Google Scholar] [CrossRef] [Green Version]
- López-Banda, D.A.; Carrillo-Casas, E.M.; Leyva-Leyva, M.; Orozco-Hoyuela, G.; Manjarrez-Hernández, Á.H.; Arroyo-Escalante, S.; Moncada-Barrón, D.; Villanueva-Recillas, S.; Xicohtencatl-Cortes, J.; Hernández-Castro, R. Identification of Virulence Factors Genes in Escherichia coli Isolates from Women with Urinary Tract Infection in Mexico. Biomed Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tabasi, M. Genotypic Characterization of Virulence Factors in Escherichia coli Isolated from Patients with Acute Cystitis, Pyelonephritis and Asymptomatic Bacteriuria. J. Clin. DIAGNOSTIC Res. 2016, 12, DC01–DC07. [Google Scholar] [CrossRef]
- Gao, Q.; Zhang, D.; Ye, Z.; Zhu, X.; Yang, W.; Dong, L.; Gao, S.; Liu, X. Virulence traits and pathogenicity of uropathogenic Escherichia coli isolates with common and uncommon O serotypes. Microb. Pathog. 2017, 104, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Ordaz-López, V.I.; Manzo-Banales, H.M.; García-Herrera, H.; Cerda-Rivera, P.E.; Ochoa, M.C.; Ramírez-Leyva, D.H. Urinary Tract Infection in Pregnancy: A Study of Pathogen and Bacterial Resistance in Mexico. J. Fam. Med.. 2016, 3, 1–4. [Google Scholar]
- Bravata-Alcantara, J.C.; Bello-Lopez, J.M.; Cortes-Ortiz, I.A.; Mendez-Velazquez, J.J.; Aviles-Soto, B.; Quintas-Granados, L.I.; Chavez-Ocana, S.d.C.; Rosel-Pech, C.; Gonzalez-Barrios, J.A.; Sierra Martinez, M. Distribution of Virulence and Antimicrobial Resistance Genes in Phylogenetic Groups of Escherichia coli Strains Isolated from Mexican Patients with Urinary Infection. Jundishapur J. Microbiol. 2019. In Press. [Google Scholar] [CrossRef] [Green Version]
- Luna-Pineda, V.M.; Ochoa, S.A.; Cruz-Córdova, A.; Cázares-Domínguez, V.; Reyes-Grajeda, J.P.; Flores-Oropeza, M.A.; Arellano-Galindo, J.; Castro-Hernández, R.; Flores-Encarnación, M.; Ramírez-Vargas, A.; et al. Features of urinary Escherichia coli isolated from children with complicated and uncomplicated urinary tract infections in Mexico. PLoS One 2018, 13, e0204934. [Google Scholar] [CrossRef] [Green Version]
- Asadi, S.; Kargar, M.; Solhjoo, K.; Najafi, A.; Ghorbani-Dalini, S. The association of virulence determinants of uropathogenic Escherichia coli with antibiotic resistance. Jundishapur J. Microbiol. 2014, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Yun, K.W.; Kim, H.Y.; Park, H.K.; Kim, W.; Lim, I.S. Virulence factors of uropathogenic Escherichia coli of urinary tract infections and asymptomatic bacteriuria in children. J. Microbiol. Immunol. Infect. 2014, 47, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Chiñas, U.; Pérez-Ramos, A.; Belmont-Monroy, L.; Chávez-Berrocal, M.E.; González-Villalobos, E.; Navarro-Ocaña, A.; Eslava, C.A.; Molina-Lopez, J. Characterization of auto-agglutinating and non-typeable uropathogenic Escherichia coli strains. J. Infect. Dev. Ctries. 2019, 13, 465–472. [Google Scholar] [CrossRef]
- Dobrindt, U.; Blum-Oehler, G.; Nagy, G.; Schneider, G.; Johann, A.; Gottschalk, G.; Hacker, J. Genetic Structure and Distribution of Four Pathogenicity Islands (PAI I536 to PAI IV536) of Uropathogenic Escherichia coli Strain 536. Infect. Immun. 2002, 70, 6365–6372. [Google Scholar] [CrossRef] [Green Version]
- Manjarrez-Hernandez, A.; Molina-López, J.; Gavilanes-Parra, S.; Hernandez-Castro, R. Escherichia coli clonal group A among uropathogenic infections in Mexico City. J. Med. Microbiol. 2016, 65, 1438–1444. [Google Scholar] [CrossRef]
- Bien, J.; Sokolova, O.; Bozko, P. Role of Uropathogenic Escherichia coli Virulence Factors in Development of Urinary Tract Infection and Kidney Damage. Int. J. Nephrol. 2012, 2012, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyer, D.M.; Radulovic, S.; Jones, F.-E.; Mobley, H.L.T. Sat, the Secreted Autotransporter Toxin of Uropathogenic Escherichia coli, Is a Vacuolating Cytotoxin for Bladder and Kidney Epithelial Cells. Infect. Immun. 2002, 70, 4539–4546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, J. The role of RTX toxins in host specificity of animal pathogenic Pasteurellaceae. Vet. Microbiol. 2011, 153, 51–58. [Google Scholar] [CrossRef]
- Gur, C.; Coppenhagen-Glazer, S.; Rosenberg, S.; Yamin, R.; Enk, J.; Glasner, A.; Bar-On, Y.; Fleissig, O.; Naor, R.; Abed, J.; et al. Natural killer cell-mediated host defense against uropathogenic E. coli is counteracted by bacterial hemolysinA-dependent killing of NK cells. Cell Host. Microbe. 2013, 14, 664–674. [Google Scholar] [CrossRef] [Green Version]
- Diabate, M.; Munro, P.; Garcia, E.; Jacquel, A.; Michel, G.; Obba, S.; Goncalves, D.; Luci, C.; Marchetti, S.; Demon, D.; et al. Escherichia coli α-Hemolysin Counteracts the Anti-Virulence Innate Immune Response Triggered by the Rho GTPase Activating Toxin CNF1 during Bacteremia. PLOS Pathog. 2015, 11, e1004732. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | Size Product | Tm °C | Reference |
|---|---|---|---|---|
| fimH | Forward: TTATGGCGGCGTGTTATC Reverse: TCCCTACTGCTCCTAACG | 545 bp | 54 | This study |
| sfaD/focC | Forward: AGGCAAATGGACAGGTATGG Reverse: TCACCCAGAACAAACTTTCC | 412 bp | This study | |
| papG-II | Forward: ATTCACCATAGAGGCGACTG Reverse: ATCATTATGCGGCTCAGAC | 237 bp | This study | |
| papC | Forward: TTCTCTCTCCCTCAATACGG Reverse: TTATAACCTCAACGGGACGG | 926 bp | This study | |
| fliCD | Forward: CCGAATCAGAGTTAGTTCCG Reverse: CCCAGCGATGAAATACTTGC | 610 bp | 60 | This study |
| sat | Forward: GTTGGCAAACAGGTCAAAC Reverse: CTCGGAGTATTGGCTTCAG | 809 bp | This study | |
| hlyA | Forward: GATACGCTGATAGGTGAG Reverse: CCAGGTGTGACTCAATAC | 564 bp | 58 | This study |
| kpsM | Forward: CCAGAGTAGATATGACCAG Reverse: CTACGAGAAATACGAACAC | 409 bp | This study | |
| agn43 | Forward: CACACAGCCACTAATACC Reverse: CACCTGAATACCCTTACC | 488 bp | 58 | This study |
| vat | Forward: ATACAGTCTCGTCTCTGG Reverse GTGACAGTCCCTTTATCC | 670 bp | This study | |
| cnf-1 | Forward: CAGACTCATCTTCACTCG Reverse: AGACAGAGACCTTACGAC | 551 bp | This study | |
| traT | Forward: TGGTATAGTTCACATCTTCC Reverse: TAAAGCCTACTACTGGATTC | 233 bp | This study | |
| fyuA | Forward: CGCCAGTAAACAATCTTCCC Reverse: CCCAAACACCATATCAACGG | 937 bp | 60 | This study |
| iucD | Forward: CGTGAGACCCAGTTTATTTCC Reverse: GGGCTGCTGAAGATATGAATAACC | 334 bp | This study | |
| iroN | Forward: CAGAATGATGCGGTAACTCC Reverse: CGTGAGACCCAGTTTATTTCC | 435 bp | This study | |
| iutA | Forward: GTTCACGCTCTTTGTCAGG Reverse: GGGCTTAATCTCGGGAAAGG | 801 bp | This study | |
| feoB | Forward: GTCTAACCTTGAGCGTAACC Reverse: GGCGAGGAAGATAGTCAGC | 736 bp | This study | |
| iha | Forward: TGTGCTCTGGTTTGATATGG Reverse: CATTCTGGGTGCCTTATATCC | 594 bp | This study | |
| ybbW | Forward: TGATTGGCAAAATCTGGCCG Reverse: ATACTGGCAATCAGTACGCC | 667 bp | [20] |
| (a) Included Patients’ Groups (n = 85) | (b) Patients with UTI Caused by UPEC (n = 37) | |||||
|---|---|---|---|---|---|---|
| With UTI n = 40 (%) | Without UTI n = 45 (%) | p | With Morphotype n = 24 (%) | Without Morphotype n = 13 (%) | p | |
| Urinalysis | ||||||
| pH: | ||||||
| 5.0–6.5 | 33 (83) | 39 (87) | 0.55 | 22 (92) | 11 (85) | 0.6 |
| 7.0–8.0 | 7 (17) | 6 (13) | 2 (8) | 2 (15) | ||
| LE: | ||||||
| Positive | 25 (63) * | 12 (27) | 0.002 | 16 (67) | 7 (53) | 0.49 |
| Negative | 15 (37) | 33 (73) | 8 (33) | 6 (46) | ||
| Pyuria: | ||||||
| Positive (>5 WBCs/HPF) | 31 (78) * | 13 (29) | <0.0001 | 14 (58) | 10 (85) | 0.72 |
| Negative (<5 WBCs/HPF) | 9 (22) | 32 (71) | 10 (42) | 4 (15) | ||
| Bacteriuria: | ||||||
| >2+ | 23 (58) | 0 (0) | <0.0001 | 7 (29) | 13 (100) | <0.0001 |
| <2+ | 17 (42) | 45 (100) * | 17 (71) * | 0 (0) | ||
| Comorbidities | ||||||
| Diabetes | 11 (28) | 12 (27) | >0.99 | 7 (29) | 4 (31) | >0.99 |
| Renal failure | 3 (8) | 3 (7) | >0.99 | 1 (4) | 2 (15) | 0.28 |
| Hypertension | 10 (25) | 12 (27) | >0.99 | 5 (20) | 3 (23) | >0.99 |
| Diagnosed ITU | 6 (15) | 2 (4) | 0.12 | 3 (13) | 3 (23) | 0.64 |
| Vaginal infection | 4 (10) | 4 (9) | >0.99 | 0 (0) * | 3 (23) | 0.037 |
| Pregnancy | 3 (8) | 1 (2) | 0.33 | 2 (8) | 1 (8) | >0.99 |
| Hypothyroidism | 1 (3) | 5 (11) | 0.2 | 0 (0) | 1 (8) | 0.35 |
| Urine culture | ||||||
| <100,000 CFU/mL | 17 (43) | 45 (100) * | <0.0001 | 17 (71) | - | <0.0001 |
| >100,000 CFU/mL | 20 (50) | 0 (0) | 7 (29) | 13 (100%) | ||
| Virulence Profile | Clinical Isolates |
|---|---|
| fimH, feoB, fliCD, cnf-1, sfaD/focC | 2,4–6,9–13,16,20,22–24,29,31,33–37,39–40 |
| fimH, feoB, fliCD, cnf-1, sfaD/focC, traT, papG-II, kpsM | 2,5,6,9,12,13,20,23,37 |
| fimH. feoB, fliCD, cnf-1, sfaD/focC, traT, papG-II, kpsM, vat, sat | 5,6,12,37 |
| Gene | % (n = 40) | r | p | Reported PAI a |
|---|---|---|---|---|
| papG-II: | PAI I, IICFT073, PAI I-V536, PAI I-IIJ96 | |||
| papC | 32.5 | 0.49 | 0.001 | PAI I-IICFT073 |
| vat | 40 | 0.47 | 0.002 | Not named PAI Ec222 |
| iroN | 45 | 0.38 | 0.017 | PAI III536 |
| fyuA | 42.5 | 0.39 | 0.012 | PAI IICFT073, PAI III, IV536 |
| sfaD/focC | 72.5 | 0.54 | <0.001 | PAI I, IICFT073, PAI I-IVJ96, PAI I-IIJ96, PAI I-III536 |
| papC: | PAI IICFT073 | |||
| iroN | 30 | 0.38 | 0.0141 | PAI III536 |
| fyuA | 30 | 0.45 | 0.003 | PAI IICFT073, PAI III, IV536 |
| sat: | PAI IICFT073 | |||
| vat | 27.5 | 0.35 | 0.028 | Not named PAI Ec222 |
| iroN | 35 | 0.46 | 0.002 | PAI III536 |
| fyuA | 35 | 0.53 | <0.001 | PAI IICFT073, PAI III, IV536 |
| kpsM | 32.5 | 0.35 | 0.02 | PAI V536 |
| iucD | 35 | 0.39 | 0.01 | PAI IICFT073, PAI III-IV536, Not named PAI UMN026 |
| hlyA: | PAI ICFT073, PAI I-II536, PAI I-IIJ96 | |||
| kpsM | 35 | 0.53 | <0.001 | PAI V536 |
| cnf-1 | 35 | 0.33 | 0.04 | PAI IIJ96 |
| vat: | Not named PAI Ec222 | |||
| cnf-1 | 42.5 | 0.32 | 0.05 | PAI IIJ96 |
| cnf-1: | PAI IIJ96 | |||
| iha | 37.5 | 0.35 | 0.025 | PAI I, IICFT073, PAI I-IVJ96, PAI I-IIJ96, PAI I-II536 |
| KpsM | 52.5 | 0.35 | 0.025 | PAI V536 |
| iroN: | PAI III536 | |||
| fyuA | 47.5 | 0.59 | <0.001 | PAI IICFT073, PAI III, IV536 |
| Gene | NT n = 10 (%) | p | B1 n = 10 (%) | p | B2 n = 11 (%) | p | C n = 3 (%) | p | E n = 6 (%) | p | Total n = 40 (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| fimH | 10 (100) | 1 | 10 (100) | 1 | 11 (100) | 1 | 3 (100) | 1 | 6 (100) | 1 | 14 (35) |
| papC | 2 (20) | 0.7 | 2 (20) | 0.7 | 7 (64) | 0.02 | 0 (0) | 0.53 | 3 (50) | 0.64 | 40 (100) |
| papG-II | 6 (60) | 1 | 4 (40) | 0.15 | 9 (82) | 0.13 | 3 (100) | 0.26 | 2 (33) | 0.19 | 29 (73) |
| sfaD/focC | 8 (80) | 0.69 | 9 (90) | 0.23 | 7 (64) | 0.45 | 3 (100) | 0.54 | 2 (33) | 0.03 | 24 (60) |
| fliCD | 10 (100) | 0.55 | 9 (90) | 1 | 10 (91) | 1 | 3 (100) | 1 | 4 (67) | 0.09 | 36 (90) |
| cnf-1 | 8 (80) | 1 | 7 (70) | 0.68 | 9 (82) | 0.69 | 2 (67) | 1 | 4 (67) | 0.62 | 16 (40) |
| vat | 4 (40) | 0.72 | 4 (40) | 0.72 | 6 (55) | 0.72 | 2 (67) | 0.59 | 3 (50) | 1 | 15 (38) |
| sat | 3 (30) | 0.71 | 1 (10) | 0.06 | 7 (64) | 0.08 | 2 (67) | 0.55 | 3 (50) | 0.66 | 24 (60) |
| hlyA | 4 (40) | 1 | 4 (40) | 1 | 6 (55) | 0.27 | 0 (0) | 0.27 | 1 (17) | 0.38 | 19 (48) |
| feoB | 10 (100) | 1 | 9 (90) | 1 | 11 (100) | 1 | 3 (100) | 1 | 6 (100) | 1 | 30 (75) |
| iucD | 4 (40) | 0.12 | 5 (50) | 0.27 | 9 (82) | 0.26 | 2 (67) | 1 | 6 (100) | 0.07 | 29 (73) |
| iroN | 4 (40) | 0.15 | 3 (30) | 0.05 | 10 (91) | 0.02 | 3 (100) | 0.26 | 4 (67) | 1 | 11 (28) |
| fyuA | 4 (40) | 0.3 | 1 (10) | 0.002 | 11 (100) | 0.0003 | 2 (67) | 0.99 | 4 (67) | 0.67 | 24 (60) |
| iha | 1 (10) | 0.03 | 4 (40) | 1 | 6 (55) | 0.29 | 2 (67) | 0.55 | 3 (50) | 0.66 | 26 (65) |
| iutA | 0 (0) | 0.55 | 3 (30) | 0.71 | 9 (82) | 1 | 0 (0) | 1 | 0 (0) | 1 | 22 (55) |
| traT | 9 (90) | 0.23 | 4 (40) | 0.01 | 8 (73) | 1 | 2 (67) | 1 | 6 (100) | 0.16 | 16 (40) |
| kpsM | 5 (50) | 0.48 | 5 (50) | 0.48 | 9 (82) | 0.14 | 1 (33) | 0.55 | 4 (67) | 0.63 | 4 (10) |
| agn43 | 2 (20) | 0.69 | 4 (40) | 0.41 | 1 (9) | 0.23 | 2 (67) | 0.17 | 2 (33) | 1 | 39 (98) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballesteros-Monrreal, M.G.; Arenas-Hernández, M.M.P.; Barrios-Villa, E.; Juarez, J.; Álvarez-Ainza, M.L.; Taboada, P.; De la Rosa-López, R.; Bolado-Martínez, E.; Valencia, D. Bacterial Morphotypes as Important Trait for Uropathogenic E. coli Diagnostic; a Virulence-Phenotype-Phylogeny Study. Microorganisms 2021, 9, 2381. https://doi.org/10.3390/microorganisms9112381
Ballesteros-Monrreal MG, Arenas-Hernández MMP, Barrios-Villa E, Juarez J, Álvarez-Ainza ML, Taboada P, De la Rosa-López R, Bolado-Martínez E, Valencia D. Bacterial Morphotypes as Important Trait for Uropathogenic E. coli Diagnostic; a Virulence-Phenotype-Phylogeny Study. Microorganisms. 2021; 9(11):2381. https://doi.org/10.3390/microorganisms9112381
Chicago/Turabian StyleBallesteros-Monrreal, Manuel G., Margarita M. P. Arenas-Hernández, Edwin Barrios-Villa, Josue Juarez, Maritza Lizeth Álvarez-Ainza, Pablo Taboada, Rafael De la Rosa-López, Enrique Bolado-Martínez, and Dora Valencia. 2021. "Bacterial Morphotypes as Important Trait for Uropathogenic E. coli Diagnostic; a Virulence-Phenotype-Phylogeny Study" Microorganisms 9, no. 11: 2381. https://doi.org/10.3390/microorganisms9112381
APA StyleBallesteros-Monrreal, M. G., Arenas-Hernández, M. M. P., Barrios-Villa, E., Juarez, J., Álvarez-Ainza, M. L., Taboada, P., De la Rosa-López, R., Bolado-Martínez, E., & Valencia, D. (2021). Bacterial Morphotypes as Important Trait for Uropathogenic E. coli Diagnostic; a Virulence-Phenotype-Phylogeny Study. Microorganisms, 9(11), 2381. https://doi.org/10.3390/microorganisms9112381