
Identification of Bull Semen Microbiome by 16S Sequencing and Possible Relationships with Fertility

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Semen Collection
2.2. DNA Extraction
2.3. 16S rRNA Amplification and Sequencing
2.4. 16S Profiling
2.5. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Foote, H.R.; Siddhartha, L.S.; Parks, E.J. Artificial insemination. In Reference Module in Food Science; Cornell University: Ithaca, NY, USA; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Al-Kass, Z.; Eriksson, E.; Bagge, E.; Wallgren, M.; Morrell, J.M. Microbiota of semen from stallions in Sweden identified by MALDI-TOF. Vet. Anim. Sci. 2020, 10, 100143. [Google Scholar] [CrossRef]
- Thibier, M.; Guerin, B. Hygienic aspects of storage and use of semen for artificial insemination. Anim. Reprod. Sci. 2000, 62, 233–251. [Google Scholar] [CrossRef]
- Bielanski, A. Disinfection procedures for controlling microorganisms in the semen and embryos of humans and farm animals. Theriogenology 2007, 68, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Sannat, C.; Nair, A.; Sahu, S.B.; Sahasrabudhe, S.A.; Kumar, A.; Gupta, A.K.; Shende, R.K. Critical sources of bacterial contamination and adoption of standard sanitary protocol during semen collection and processing in Semen Station. Vet. World 2015, 8, 631–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Council Directive 88/407/EEC of 18 June 1988 Laying down the Animal Health Requirements Applicable to Intra-Community Trade in and Imports of Deep-Frozen Semen of Domestic Animals of the Bovine Species Annex C2. Available online: https://www.eumonitor.eu/9353000/1/j9vvik7m1c3gyxp/vhckkm8vsrz6 (accessed on 23 November 2021).
- Zampieri, D.; Santos, V.G.; Braga, P.A.C.; Ferreira, R.C.; Ballottin, D.; Tasic, L.; Basso, A.C.; Sanches, B.V.; Pontes, J.H.F.; Pereira da Silva, B.; et al. Microorganisms in cryopreserved semen and culture media used in the in vitro production (IVP) of bovine embryos identified by matrix-assistedlaser desorption ionization mass spectrometry (MALDI-MS). Theriogenology 2013, 80, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Catry, B.; Van Duijkeren, E.; Pomba, M.C.; Greko, C.; Moreno, M.A.; Pyörälä, S.; Ruzauskas, M.; Sanders, P.; Threlfall, E.J.; Ungemach, F.; et al. Reflection paper on MRSA in food-producing and companion animals: Epidemiology and control options for human and animal health. Epidemiol. Infect. 2010, 138, 626–644. [Google Scholar] [CrossRef] [PubMed]
- Baud, D.; Pattaroni, C.; Vulliemoz, N.; Castella, V.; Marsland, J.B.; Stojanov, M. Sperm microbiota and its impact on semen parameters. Front. Microbial. 2019, 10, 234. [Google Scholar] [CrossRef] [Green Version]
- Del Porto, G.B.; Derrick, F.C.; Bannister, E.R. Bacterial effect on sperm motility. Urology 1975, 5, 638–639. [Google Scholar] [CrossRef]
- Cagnoli, C.I.; Chiapparrone, M.L.; Cacciato, C.S.; Rodríguez, M.G.; Aller, J.F.; Catena, M.D.C. Effects of Campylobacter fetus on bull sperm quality. Microb. Pathog. 2020, 149, 104486. [Google Scholar] [CrossRef] [PubMed]
- González-Marín, C.; Roy, R.; López-Fernández, C.; Diez, B.M.; Carabaño, M.J.; Fernández, J.L.; Kjelland, M.E.; Moreno, J.F.; Gosálvez, J. Bacteria in bovine semen can increase sperm DNA fragmentation rates: A kinetic experimental approach. Anim. Reprod. Sci. 2011, 123, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Marchiani, S.; Baccani, I.; Tamburrino, L.; Mattiuz, G.; Nicolò, S.; Bonaiuto, C.; Panico, C.; Vignozzi, L.; Antonelli, A.; Rossolini, G.; et al. Effects of common gram-negative pathogens causing male genitourinary-tract infections on human sperm functions. Sci. Rep. 2021, 11, 19177. [Google Scholar] [CrossRef] [PubMed]
- Oghbaei, H.; Rastgar Rezaei, Y.; Nikanfar, S.; Zarezadeh, R.; Sadegi, M.; Latifi, Z.; Nouri, M.; Fattahi, A.; Ahmadi, Y.; Bleisinger, N. Effects of bacteria on male fertility: Spermatogenesis and sperm function. Life Sci. 2020, 256, 117891. [Google Scholar] [CrossRef] [PubMed]
- Eini, F.; Kutenaei, M.A.; Zareei, F.; Dastjerdi, Z.S.; Shirzeyli, M.H.; Salehi, E. Effect of bacterial infection on sperm quality and DNA fragmentation in subfertile men with Leukocytospermia. BMC Mol. Cell Biol. 2021, 22, 42. [Google Scholar] [CrossRef]
- Givens, M.D.; Marley, M.S.D. Pathogens that cause infertility of bulls or transmission via semen. Theriogenology 2008, 70, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Appiah, M.O.; Wang, J.; Lu, W. Microflora in the reproductive tract of cattle: A Review. Agriculture 2020, 10, 232. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Owens, S.E. Postpartum uterine infection and endometritis in dairy cattle. Anim. Reprod. 2018, 14, 622–629. [Google Scholar] [CrossRef]
- Fourichon, C.; Seegers, H.; Malher, X. Effect of disease on reproduction in the dairy cow: A metaanalysis. Theriogenology 2000, 53, 1729–1759. [Google Scholar] [CrossRef]
- Borsberry, S.; Dobson, H. Periparturient diseases and their effect on reproductive performance in five dairy herds. Vet. Rec. 1989, 124, 217–219. [Google Scholar] [CrossRef]
- LeBlanc, S.J.; Duffield, T.F.; Leslie, K.E.; Bateman, K.G.; Keefe, G.P.; Walton, J.S.; Johnson, W.H. Defining and diagnosing postpartum clinical endometritis and its impact on reproductive performance in dairy cows. J. Dairy Sci. 2002, 85, 2223–2236. [Google Scholar] [CrossRef]
- Marey, M.A.; Yousef, M.S.; Kowsar, R.; Hambruch, N.; Shimizu, T.; Pfarrer, C.; Miyamoto, A. Local immune system in oviduct physiology and pathophysiology: Attack or tolerance? Domest. Anim. Endocrinol. 2016, 56, 204–211. [Google Scholar] [CrossRef]
- Karsch, F.J.; Battaglia, D.F.; Breen, K.M.; Debus, N.; Harris, T.G. Mechanisms for ovarian cycle disruption by immune/inflammatory stress. Stress 2002, 5, 101–112. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Noakes, D.E.; Rycroft, A.N.; Pfeiffer, D.U.; Dobson, H. Influence of uterine bacterial contamination after parturition on ovarian dominant follicle selection and follicle growth and function in cattle. Reproduction 2002, 123, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Bromfield, J.J.; Sheldon, I.M. Lipopolysaccharide initiates inflammation in bovine granulosa cells via the TLR4 pathway and perturbs oocyte meiotic progression in vitro. Endocrinology 2011, 152, 5029–5040. [Google Scholar] [CrossRef] [Green Version]
- Al-Kass, Z.; Guo, Y.; Vinnere Pettersson, O.; Niazi, A.; Morrell, J.M. Metagenomic analysis of bacteria in stallion semen. Anim. Reprod. Sci. 2020, 221, 106568. [Google Scholar] [CrossRef] [PubMed]
- Althouse, G.C.; Lu, K.G. Bacteriospermia in extended porcine semen. Theriogenology 2005, 63, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Yániz, J.L.; Marco-Aguado, M.A.; Mateos, J.A.; Santolaria, P. Bacterial contamination of ram semen, antibiotic sensitivities, and effects on sperm quality during storage at 15°C. Anim. Reprod. Sci. 2010, 122, 142–1499. [Google Scholar] [CrossRef] [PubMed]
- Sanocka-Maciejewska, D.; Ciupińska, M.; Kurpisz, M. Bacterial infection and semen quality. J. Reprod. Immunol. 2005, 67, 51–56. [Google Scholar] [CrossRef]
- Enwuru, C.A.; Iwalokun, B.; Enwuru, V.N.; Ezechi, O.; Oluwadun, A. The effect of presence of facultative bacteria species on semen and sperm quality of men seeking fertility care. Afr. J. Urol. 2016, 22, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Prince, P.W.; Almquist, J.O.; Reid, J.J. Bacteriological studies of bovine semen. II. The incidence of specific types of bacteria and the relation to fertility. J. Dairy Sci. 1949, 32, 849–855. [Google Scholar] [CrossRef]
- Riigi Teataja. Loomakaitseseadus—Riigi Teataja. Available online: https://www.riigiteataja.ee/akt/LoKS (accessed on 24 September 2021).
- Miglior, F.; Pizzi, F.; Guaita, N. Effect of environmental factors on non return rate in Italian Holstein-Friesians. Interbull. Bull. 1997, 27, 106–108. [Google Scholar]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T. Development of a prokaryotic universal primer for simultaneous analysis of bacteria and archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. A quality control tool for high throughput sequence data. FASTQC 2010, 532, 1. [Google Scholar]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tse, H.; Yuen, K.Y. Then and now: Use of 16S rDNA gene sequencing for bacterial identification and discovery of novel bacteria in clinical microbiology laboratories. Clin. Microbiol. Infect. 2008, 14, 908–934. [Google Scholar] [CrossRef] [Green Version]
- Gunsalus, I.C.; Salisbury, G.W.; Willett, E.L. The bacteriology of bull semen. J. Dairy Sci. 1941, 24, 911–919. [Google Scholar] [CrossRef]
- Bush, L.J.; Ludwick, T.M.; Ferguson, L.C.; Ely, F. The effect of bacteria on the fertility of bovine semen. J. Dairy Sci. 1950, 33, 633–638. [Google Scholar] [CrossRef]
- Mitra, J.; Chowdhury, S.; Panda, S.; Chakraborty, M.; Singha, A. Microbiological evaluation of bovine frozen semen samples in West Bengal, India. Explor. Anim. Med. Res. 2016, 6, 185–191. [Google Scholar]
- Balqis, U.; Hambal, M.; Admi, M.; Safika, S.; Meutia, N.; Sugit, S.; Dasrul; Abdullah, M.A.N.; Ferasyi, T.R.; Lubis, T.M.; et al. Escherichia fergusonii identified in preputial swabs from healthy Aceh cattle by phylogenetic 16S rRNA analysis. Malays. J. Microbiol. 2018, 14, 229–235. [Google Scholar] [CrossRef]
- Hambal, M.; Admi, M.; Safika, S.; Sari, W.E.; Ferasyi, T.R.; Dasrul, D.; Balqis, U.; Darmawi, D. Identification of Staphylococcus species isolated from preputium of Aceh cattle based on 16S rRNA gene sequences analysis. Vet. World 2019, 12, 1540–1545. [Google Scholar] [CrossRef] [PubMed]
- Haapala, V.; Pohjanvirta, T.; Vähänikkilä, N.; Halkilahti, J.; Simonen, H.; Pelkonen, S.; Soveri, T.; Simojoki, H.; Autio, T. Semen as a source of Mycoplasma bovis mastitis in dairy herds. Vet. Microbiol. 2018, 216, 60–66. [Google Scholar] [CrossRef]
- Wickware, C.L.; Johnson, T.A.; Koziol, J.H. Composition and diversity of the preputial microbiota in healthy bulls. Theriogenology 2020, 145, 231–237. [Google Scholar] [CrossRef]
- Lima, S.F.; Teixeira, A.G.V.; Higgins, C.H.; Lima, F.S.; Bicalho, R.C. The upper respiratory tract microbiome and its potential role in bovine respiratory disease and otitis media. Sci. Rep. 2016, 6, 29050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein-Jöbstl, D.; Quijada, N.M.; Dzieciol, M.; Feldbacher, B.; Wagner, M.; Drillich, M.; Schmitz-Esser, S.; Mann, E. Microbiota of newborn calves and their mothers reveals possible transfer routes for newborn calves’ gastrointestinal microbiota. PLoS ONE 2019, 14, e0220554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bay, V.; Griffiths, B.; Carter, S.; Evans, N.J.; Lenzi, L.; Bicalho, R.C.; Oikonomou, G. 16S rRNA amplicon sequencing reveals a polymicrobial nature of complicated claw horn disruption lesions and interdigital phlegmon in dairy cattle. Sci. Rep. 2018, 8, 15529. [Google Scholar] [CrossRef]
- Kim, R.; Reboli, C.A. Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 8th ed.; ESEVIRA Saunders: Philadelphia, PA, USA, 2015; Volume 2, pp. 2373–2382. [Google Scholar]
- Foster, A.P. Staphylococcal skin disease in livestock. Vet. Dermatol. 2012, 23, 342–351. [Google Scholar] [CrossRef]
- Duus, L.M.; Høiby, N.; Wang, M.; Schiøtz, O.; Nørskov-Lauritsen, N. Bacteria of the genus Dyella can chronically colonise the airways of patients with cystic fibrosis and elicit a pronounced antibody response. Int. J. Med. Microbiol. 2013, 303, 267–269. [Google Scholar] [CrossRef]
- Piau, C.; Arvieux, C.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Capnocytophaga spp. involvement in bone infections: A review. Int. J. Antimicrob. Agents 2013, 41, 509–515. [Google Scholar] [CrossRef]
- Vergne, A.; Guérin, F.; Lienhard, R.; Le Coustumier, A.; Daurel, C.; Isnard, C.; Marty, N.; Poyart, C.; Cattoir, V. Identification and Clinical Significance of Helcococcus kunzii in Human Samples. J. Clin. Microbiol. 2015, 53, 2703–2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Headley, S.A.; Voltarelli, D.; de Oliveira, V.H.; Bronkhorst, D.E.; Alfieri, A.F.; Filho, L.C.; Okano, W.; Alfieri, A.A. Association of Histophilus somni with spontaneous abortions in dairy cattle herds from Brazil. Trop. Anim. Health Prod. 2015, 47, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Michi, A.N.; Favetto, P.H.; Kastelic, J.; Cobo, E.R. A review of sexually transmitted bovine trichomoniasis and campylobacteriosis affecting cattle reproductive health. Theriogenology 2016, 85, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Farahani, L.; Tharakan, T.; Yap, T.; Ramsay, W.J.; Jayasena, N.C.; Minhas, S. The semen microbiome and its impact on sperm function and male fertility: A systematic review and meta-analysis. Andrology 2021, 9, 115–144. [Google Scholar] [CrossRef]
- Varela, E.; Rey, J.; Plaza, E.; Muñoz de Propios, P.; Ortiz-Rodríguez, J.M.; Álvarez, M.; Anel-López, L.; Anel, L.; De Paz, P.; Gil, M.C.; et al. How does the microbial load affect the quality of equine cool-stored semen? Theriogenology 2018, 114, 212–220. [Google Scholar] [CrossRef]
- Reda, A.A.; Almaw, G.; Abreha, S.; Tadeg, W.; Tadesse, B. Bacteriospermia and sperm quality of cryopreserved bull semen used in artificial insemination of cows in South Wollo zone, Ethiopia. Vet. Med. Int. 2020, 2020, 2098315. [Google Scholar] [CrossRef] [Green Version]
- Gączarzewicz, D.; Udała, J.; Piasecka, M.; Błaszczyk, B.; Stankiewicz, T. Bacterial Contamination of boar semen and its relationship to sperm quality preserved in commercial extender containing gentamicin sulfate. Pol. J. Vet. Sci. 2016, 19, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.E.; Bernard, K.A.; Harrington, S.M.; Patel, N.B.; Tucker, T.A.; Metcalfe, M.G.; McQuiston, J.R. Lawsonella clevelandensis gen. nov., sp. nov., a new member of the suborder Corynebacterineae isolated from human abscesses. Int. J. Sys.t Evol. Microbiol. 2016, 66, 2929–2935. [Google Scholar] [CrossRef]
- Flygel, T.T.; Sovershaeva, E.; Claassen-Weitz, S.; Hjerde, E.; Mwaikono, K.S.; Odland, J.Ø.; Ferrand, R.A.; Mchugh, G.; Gutteberg, T.J.; Nicol, M.P.; et al. Composition of gut microbiota of children and adolescents with perinatal human immunodeficiency virus infection taking antiretroviral therapy in Zimbabwe. J. Infect. Dis. 2020, 221, 483–492. [Google Scholar]
- Braga, R.M.; Dourado, M.N.; Araujo, W.L. Microbial interactions: Ecology in a molecular perspective. Braz. J. Microbiol. 2016, 47, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Niehaus, L.; Boland, I.; Liuz, M.; Chen, K.; Fu, D.; Henckel, C.; Chaung, K.; Miranda, S.E.; Dyckman, S.; Crum, M.; et al. Microbial coexistence through chemical-mediated interactions. Nat. Commun. 2019, 10, 2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deines, P.; Hammerschmidt, K.; Bosch, T. Microbial species coexistence depends on the host environment. mBio 2020, 11, e00807-20. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Yokota, A. Proposals of Curvibacter gracilis gen. nov., sp. nov. and Herbaspirillum putei sp. nov. for bacterial strains isolated from well water and reclassification of [Pseudomonas] huttiensis, [Pseudomonas] lanceolata, [Aquaspirillum] delicatum and [Aquaspirillum] autotrophicum as Herbaspirillum huttiense comb. nov., Curvibacter lanceolatus comb. nov., Curvibacter delicatus comb. nov. and Herbaspirillum autotrophicum comb. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 2223–2230. [Google Scholar] [PubMed]
- Graf, J. The Family Rikenellaceae. In The Prokaryotes: Other Major Lineages of Bacteria and The Archaea; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 68, pp. 857–859. [Google Scholar]
- Cai, B.; Pan, J.; Chen, H.; Chen, X.; Ye, Z.; Yuan, H.; Sun, H.; Wan, P. Oyster polysaccharides ameliorate intestinal mucositis and improve metabolism in 5-fluorouracil-treated S180 tumour-bearing mice. Carbohydr. Polym. 2021, 256, 117545. [Google Scholar] [CrossRef] [PubMed]
- Bálint, A.; Farkas, K.; Méhi, O.; Kintses, B.; Vásárhelyi, B.M.; Ari, E.; Pál, C.; Madácsy, T.; Maléth, J.; Szántó, K.J.; et al. Functional anatomical changes in ulcerative colitis patients determine their gut microbiota composition and consequently the possible treatment outcome. Pharmaceuticals 2020, 13, 346. [Google Scholar] [CrossRef]




{kind=link}
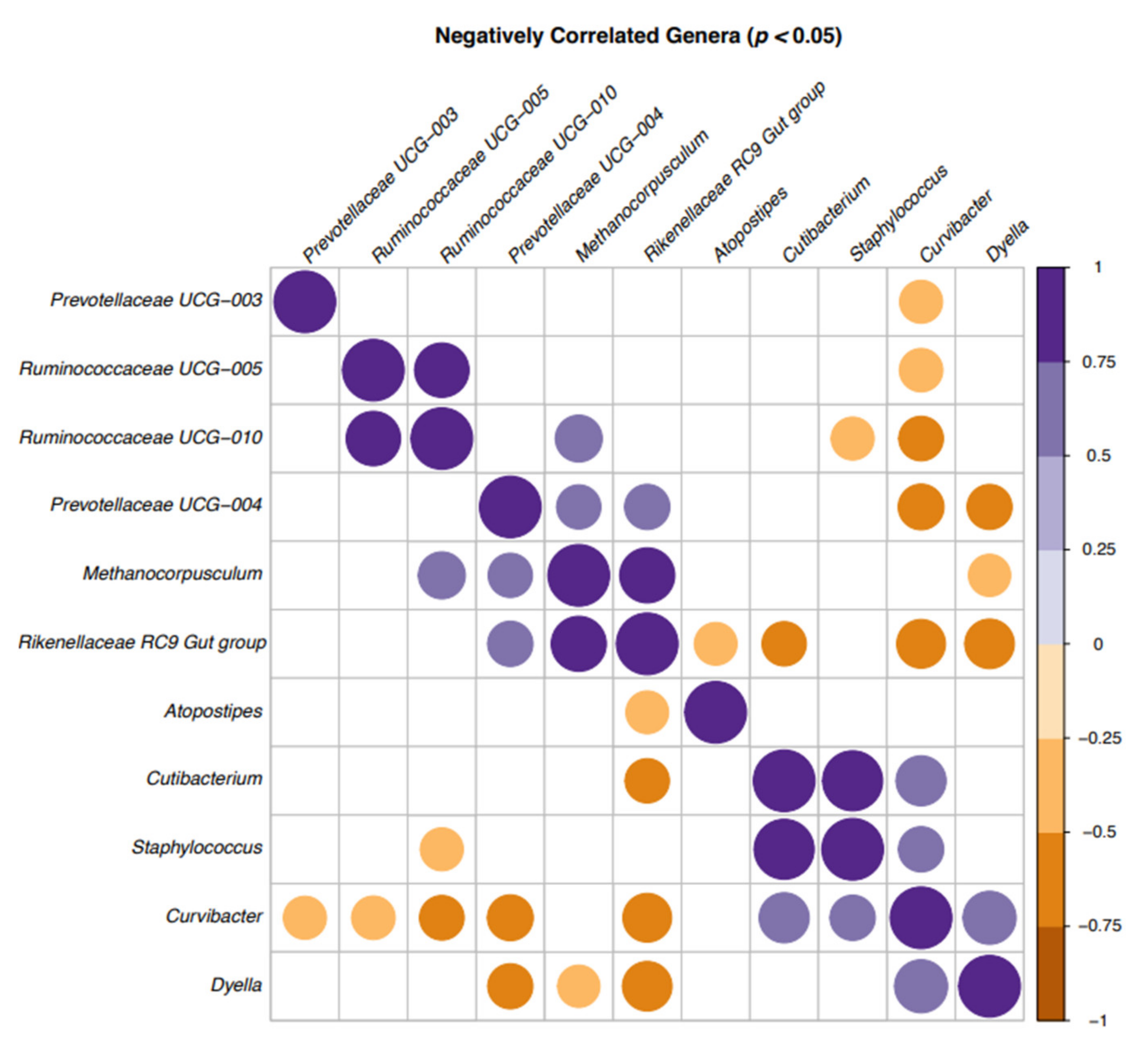{kind=link}
{kind=link}
| Primer 16S rRNA | Sequences (5′-3′) | Terminal Cycling | Reference |
|---|---|---|---|
| 341F | CCTACGGGAGGCAGCAG | (98 °C 3 min); (98 °C 30 s, 55 °C 30 s, 72 °C 40 s) × 25; 72 °C 10 min; 10 °C hold | [34] |
| 805R | GACTACNVGGGTATCTAATCC | (98 °C 3 min); (98 °C 30 s, 55 °C 30 s, 72 °C 45 s) × 8; 72 °C 5 min; 10 °C hold |
| Top 20 Genera | Counts | Total Genera | Total Counts | NRR (%) | |
|---|---|---|---|---|---|
| Bull 1 | 7 | 50,127 | 13 | 60,678 | 48.9 |
| Bull 2 | 20 | 101,453 | 69 | 149,773 | 48.3 |
| Bull 3 | 5 | 17,157 | 12 | 56,692 | 51.4 |
| Bull 4 | 17 | 53,773 | 47 | 95,193 | 55.1 |
| Bull 5 | 12 | 16,089 | 31 | 27,827 | 61.7 |
| Bull 6 | 16 | 188,695 | 47 | 247,273 | 45.9 |
| Bull 7 | 16 | 61,305 | 37 | 92,055 | 48.9 |
| Bull 8 | 17 | 74,382 | 49 | 129,282 | 52.1 |
| Bull 9 | 10 | 53,248 | 24 | 70,528 | 50.4 |
| Bull 10 | 12 | 95,853 | 26 | 114,379 | 62.1 |
| Bull 11 | 18 | 95,136 | 82 | 175,372 | 51.8 |
| Bull 12 | 19 | 70,932 | 89 | 130,601 | 55.2 |
| Bull 13 | 18 | 115,553 | 77 | 183,004 | 50.5 |
| Bull 14 | 18 | 64,435 | 76 | 118,757 | 47.6 |
| Bull 15 | 18 | 102,459 | 83 | 170,084 | 54 |
| Bull 16 | 19 | 146,754 | 53 | 198,532 | 52.6 |
| Bull 17 | 18 | 68,109 | 86 | 141,152 | 52.3 |
| Bull 18 | 9 | 116,226 | 16 | 137,258 | 37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cojkic, A.; Niazi, A.; Guo, Y.; Hallap, T.; Padrik, P.; Morrell, J.M. Identification of Bull Semen Microbiome by 16S Sequencing and Possible Relationships with Fertility. Microorganisms 2021, 9, 2431. https://doi.org/10.3390/microorganisms9122431
Cojkic A, Niazi A, Guo Y, Hallap T, Padrik P, Morrell JM. Identification of Bull Semen Microbiome by 16S Sequencing and Possible Relationships with Fertility. Microorganisms. 2021; 9(12):2431. https://doi.org/10.3390/microorganisms9122431
Chicago/Turabian StyleCojkic, Aleksandar, Adnan Niazi, Yongzhi Guo, Triin Hallap, Peeter Padrik, and Jane M. Morrell. 2021. "Identification of Bull Semen Microbiome by 16S Sequencing and Possible Relationships with Fertility" Microorganisms 9, no. 12: 2431. https://doi.org/10.3390/microorganisms9122431
APA StyleCojkic, A., Niazi, A., Guo, Y., Hallap, T., Padrik, P., & Morrell, J. M. (2021). Identification of Bull Semen Microbiome by 16S Sequencing and Possible Relationships with Fertility. Microorganisms, 9(12), 2431. https://doi.org/10.3390/microorganisms9122431