
Potential of Bacterial Strains Isolated from Coastal Water for Wastewater Treatment and as Aqua-Feed Additives



Abstract
:
1. Introduction
2. Materials and Methods
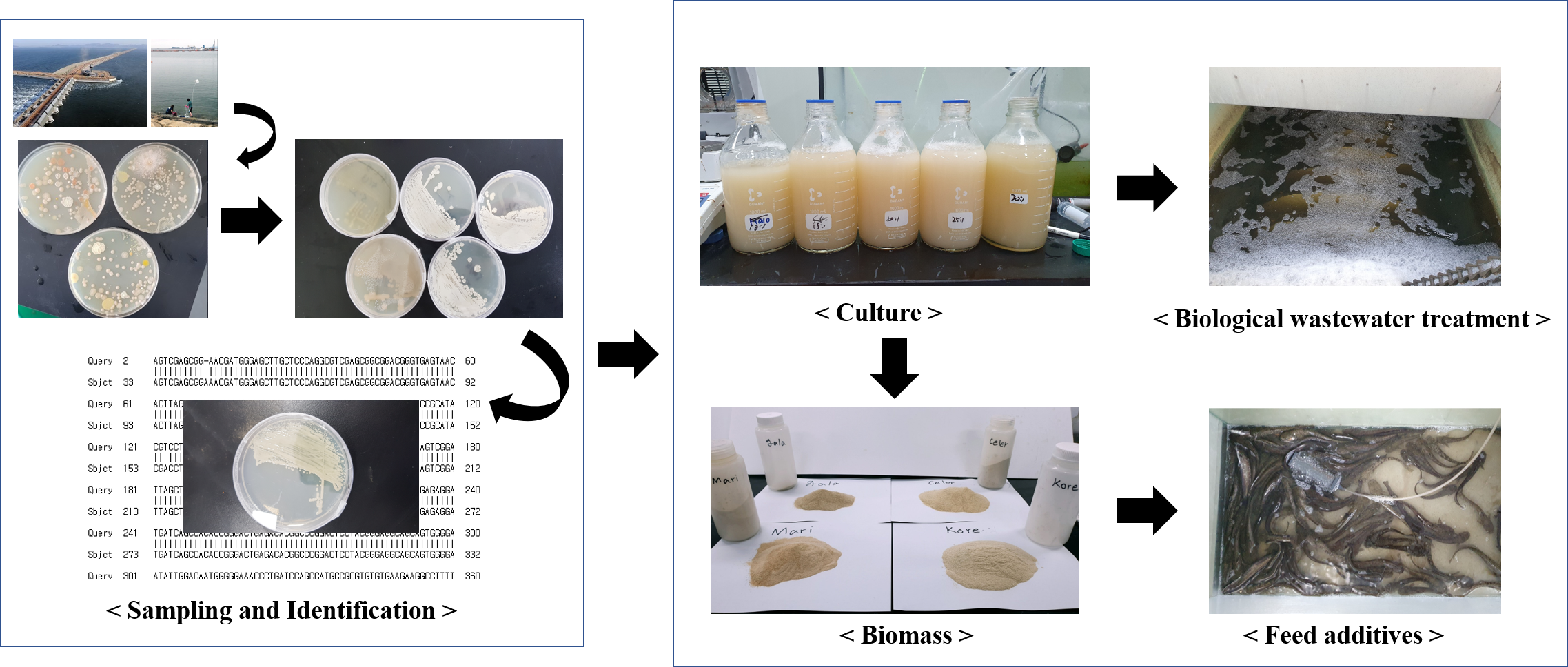
2.1. Site Description and Strains Collection
2.2. Cultivation and Harvesting of Bacteria
2.3. Evaluation of Biological Wastewater Treatment
2.4. Preparation of Experimental Diets (5% Bacterial Feed Additives), and Fish and Rearing Conditions
2.5. Growth Performance and Proximate Composition Analysis
2.6. Amino Acid Compositions
2.7. Fatty Acid Methyl Ester (FAME) Compositions
2.8. Statistical Analysis
3. Results and Discussion
3.1. Identification of Strains
3.2. Effects of 4 Candidate Strains on Nutrient Removal
3.3. Evaluation of Four Strains of Bacteria as Feed Additives for Fish-Growth Performance
3.4. Practical Applications and Future Research Prospects
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Kong, M.K.; Lee, P.C. Metabolic engineering of menaquinone-8 pathway of Escherichia coli as a microbial platform for vitamin K production. Biotechnol. Bioeng. 2011, 108, 1997–2002. [Google Scholar] [CrossRef]
- Tarvainen, M.; Fabritius, M.; Yang, B. Determination of vitamin K composition of fermented food. Food Chem. 2019, 275, 515–522. [Google Scholar] [CrossRef]
- Kijewski, A.; Witso, I.L.; Iversen, H.; Ronning, H.T.; L’Abee-Lund, T.; Wasteson, Y.; Lindback, T.; Aspholm, M. Vitamin K Analogs Influence the Growth and Virulence Potential of Enterohemorrhagic Escherichia coli. Appl. Environ. Microbiol. 2020, 86, e00583-20. [Google Scholar] [CrossRef]
- Williams, D.C.; Van Frank, R.M.; Muth, W.L.; Burnett, J.P. Cytoplasmic inclusion bodies in Escherichia coli producing biosynthetic human insulin proteins. Science 1982, 215, 687–689. [Google Scholar] [CrossRef]
- Jing, J.; Chen, Y.; Sheng, L.; Wu, M. Optimized production of insulin variant, a recombinant platelet aggregation inhibitor, by high cell-density fermentation of recombinant Escherichia coli. Protein Expr. Purif. 2018, 152, 7–12. [Google Scholar] [CrossRef]
- Goeddel, D.V.; Kleid, D.G.; Bolivar, F.; Heyneker, H.L.; Yansura, D.G.; Crea, R.; Hirose, T.; Kraszewski, A.; Itakura, K.; Riggs, A.D. Expression in Escherichia coli of chemically synthesized genes for human insulin. Proc. Natl. Acad. Sci. USA 1979, 76, 106–110. [Google Scholar] [CrossRef] [Green Version]
- DeVilbiss, S.E.; Steele, M.K.; Krometis, L.H.; Badgley, B.D. Freshwater salinization increases survival of Escherichia coli and risk of bacterial impairment. Water Res. 2021, 191, 116812. [Google Scholar] [CrossRef]
- Blaustein, R.A.; Pachepsky, Y.; Hill, R.L.; Shelton, D.R.; Whelan, G. Escherichia coli survival in waters: Temperature dependence. Water Res. 2013, 47, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, W.P.; Kim, M.; Thackston, E.L. Comparison of commercially available Escherichia coli enumeration tests: Implications for attaining water quality standards. Water Res. 2005, 39, 4869–4878. [Google Scholar] [CrossRef] [PubMed]
- OHair, J.; Jin, Q.; Yu, D.; Wu, J.; Wang, H.; Zhou, S.; Huang, H. Non-sterile fermentation of food waste using thermophilic and alkaliphilic Bacillus licheniformis YNP5-TSU for 2,3-butanediol production. Waste Manag. 2021, 120, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Petrova, P.; Petlichka, S.; Petrov, K. New Bacillus spp. with potential for 2,3-butanediol production from biomass. J. Biosci. Bioeng. 2020, 130, 20–28. [Google Scholar] [CrossRef]
- Perego, P.; Converti, A.; Del Borghi, M. Effects of temperature, inoculum size and starch hydrolyzate concentration on butanediol production by Bacillus licheniformis. Bioresour. Technol. 2003, 89, 125–131. [Google Scholar] [CrossRef]
- Sharma, P.; Tripathi, S.; Chaturvedi, P.; Chaurasia, D.; Chandra, R. Newly isolated Bacillus sp. PS-6 assisted phytoremediation of heavy metals using Phragmites communis: Potential application in wastewater treatment. Bioresour. Technol. 2021, 320, 124353. [Google Scholar] [CrossRef]
- Yezza, A.; Tyagi, R.D.; Valero, J.R.; Surampalli, R.Y. Bioconversion of industrial wastewater and wastewater sludge into Bacillus thuringiensis based biopesticides in pilot fermentor. Bioresour. Technol. 2006, 97, 1850–1857. [Google Scholar] [CrossRef]
- Al-Ansari, M.M.; Benabdelkamel, H.; Al-Humaid, L. Degradation of sulfadiazine and electricity generation from wastewater using Bacillus subtilis EL06 integrated with an open circuit system. Chemosphere 2021, 276, 130–145. [Google Scholar] [CrossRef]
- Sharp, R.J.; Scawen, M.D.; Atkinson, T. Fermentation and Downstream Processing of Bacillus. In Bacillus; Harwood, C.R., Ed.; Springer: Boston, MA, USA, 1989; pp. 255–292. [Google Scholar]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Hur, J.W.; Kim, S.; Jung, J.Y.; Han, H.S. Biological wastewater treatment: Comparison of heterotrophs (BFT) with autotrophs (ABFT) in aquaculture systems. Bioresour. Technol. 2020, 296, 122293. [Google Scholar] [CrossRef]
- Jung, J.Y.; Damusaru, J.H.; Park, Y.; Kim, K.; Seong, M.; Je, H.W.; Kim, S.; Bai, S.C. Autotrophic biofloc technology system (ABFT) using Chlorella vulgaris and Scenedesmus obliquus positively affects performance of Nile tilapia (Oreochromis niloticus). Algal Res. 2017, 27, 259–264. [Google Scholar] [CrossRef]
- Cai, Z.; Li, H.; Pu, S.; Ke, J.; Wang, D.; Liu, Y.; Chen, J.; Guo, R. Development of autotrophic and heterotrophic consortia via immobilized microbial beads for chemical wastewater treatment, using PTA wastewater as an approach. Chemosphere 2021, 281, 131001. [Google Scholar] [CrossRef] [PubMed]
- Lang, X.; Li, Q.; Xu, Y.; Ji, M.; Yan, G.; Guo, S. Aerobic denitrifiers with petroleum metabolizing ability isolated from caprolactam sewage treatment pool. Bioresour. Technol. 2019, 290, 121719. [Google Scholar] [CrossRef]
- Xie, F.; Thiri, M.; Wang, H. Simultaneous heterotrophic nitrification and aerobic denitrification by a novel isolated Pseudomonas mendocina X49. Bioresour. Technol. 2021, 319, 124198. [Google Scholar] [CrossRef]
- Yang, J.R.; Wang, Y.; Chen, H.; Lyu, Y.K. Ammonium removal characteristics of an acid-resistant bacterium Acinetobacter sp. JR1 from pharmaceutical wastewater capable of heterotrophic nitrification-aerobic denitrification. Bioresour. Technol. 2019, 274, 56–64. [Google Scholar] [CrossRef]
- Arts, P.A.M.; Robertson, L.A.; Kuenen, J.G. Nitrification and denitrification by Thiosphaera pantotropha in aerobic chemostat cultures. FEMS Microbiol. Ecol. 1995, 18, 305–316. [Google Scholar] [CrossRef]
- Chen, P.; Li, J.; Li, Q.X.; Wang, Y.; Li, S.; Ren, T.; Wang, L. Simultaneous heterotrophic nitrification and aerobic denitrification by bacterium Rhodococcus sp. CPZ24. Bioresour. Technol. 2012, 116, 266–270. [Google Scholar] [CrossRef]
- Joo, H.S.; Hirai, M.; Shoda, M. Improvement in ammonium removal efficiency in wastewater treatment by mixed culture of Alcaligenes faecalis no. 4 and L1. J. Biosci. Bioeng. 2007, 103, 66–73. [Google Scholar] [CrossRef]
- Nishio, T.; Yoshikura, T.; Chiba, K.; Inouye, Z. Effects of organic acids on heterotrophic nitrification by Alcaligenes faecalis OKK17. Biosci. Biotechnol. Biochem. 1994, 58, 1574–1578. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Fish and Shrimp; National Academy Press: Washington, DC, USA, 2011.
- Dawood, M.A.O.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Barrows, F.T. Feed additives. In Encyclopedia of Aquaculture; Stickney, R.P., Ed.; John Wiley and Sons: Hoboken, NJ, USA, 2000; pp. 335–340. [Google Scholar]
- Geng, X.; Dong, X.H.; Tan, B.P.; Yang, Q.H.; Chi, S.Y.; Liu, H.Y.; Liu, X.Q. Effects of dietary probiotic on the growth performance, non-specific immunity and disease resistance of cobia, Rachycentron canadum. Aquac. Nutr. 2012, 18, 46–55. [Google Scholar] [CrossRef]
- Kim, J.S.; Harikrishnan, R.; Kim, M.C.; Balasundaram, C.; Heo, M.S. Dietary administration of Zooshikella sp. enhance the innate immune response and disease resistance of Paralichthys olivaceus against Sreptococcus iniae. Fish Shellfish Immunol. 2010, 29, 104–110. [Google Scholar] [CrossRef]
- Nayak, S.K. Role of gastrointestinal microbiota in fish. Aquac. Res. 2010, 41, 1553–1573. [Google Scholar] [CrossRef]
- Sáenz de Rodrigáñez, M.A.; DÍaz-Rosales, P.; Chabrillón, M.; Smidt, H.; Arijo, S.; León-Rubio, J.M.; Alarcón, F.J.; Balebona, M.C.; Morinigo, M.A.; Cara, J.B.; et al. Effect of dietary administration of probiotics on growth and intestine functionality of juvenile Senegalese sole (Solea senegalensis, Kaup 1858). Aquac. Nutr. 2009, 15, 177–185. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, Y.; Dong, H.; Wang, Y.; Zheng, X.; Zhang, J. Effect of dietary Clostridium butyricum on growth, intestine health status and resistance to ammonia stress in Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2017, 65, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.F.; Liu, X.L.; Chang, Y.Q.; Liu, M.T.; Wang, G.X. Effects of dietary supplementation of probiotic Shewanella colwelliana WA64, Shewanella olleyana WA65 on the innate immunity and disease resistance of abalone, Haliotis discus hannai Ino. Fish Shellfish Immunol. 2013, 35, 86–91. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Jung, J.Y.; Hur, J.W.; Kim, K.; Han, H.S. Evaluation of floc-harvesting technologies in biofloc technology (BFT) system for aquaculture. Bioresour. Technol. 2020, 314, 123719. [Google Scholar] [CrossRef] [PubMed]
- Paweloszek, R.; Briancon, S.; Chevalier, Y.; Gilon-Delepine, N.; Pelletier, J.; Bolzinger, M.A. Skin Absorption of Anions: Part Two. Skin Absorption of Halide Ions. Pharm. Res. 2016, 33, 1576–1586. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1995. [Google Scholar]
- Folch, J.; Lees, M.; Stanley, G.H.S. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Li, W.; Ai, Q.; Mai, K.; Xu, W.; Luo, Y.; Zhang, Y. Effects of dietary amino acid patterns on growth and protein metabolism of large yellow croaker (Larimichthys crocea) larvae. Aquaculture 2013, 406–407, 1–8. [Google Scholar] [CrossRef]
- Wirasnita, R.; Hadibarata, T.; Novelina, Y.M.; Yusoff, A.R.M.; Yusop, Z. A Modified Methylation Method to Determine Fatty Acid Content by Gas Chromatography. Bull. Korean Chem. Soc. 2013, 34, 3239–3242. [Google Scholar] [CrossRef] [Green Version]
- Zaghloul, T.I.; Embaby, A.M.; Elmahdy, A.R. Biodegradation of chicken feathers waste directed by Bacillus subtilis recombinant cells: Scaling up in a laboratory scale fermentor. Bioresour. Technol. 2011, 102, 2387–2393. [Google Scholar] [CrossRef]
- Lu, L.; Tan, H.; Luo, G.; Liang, W. The effects of Bacillus subtilis on nitrogen recycling from aquaculture solid waste using heterotrophic nitrogen assimilation in sequencing batch reactors. Bioresour. Technol. 2012, 124, 180–185. [Google Scholar] [CrossRef]
- Wattiau, P.; Renard, M.E.; Ledent, P.; Debois, V.; Blackman, G.; Agathos, S.N. A PCR test to identify Bacillus subtilis and closely related species and its application to the monitoring of wastewater biotreatment. Appl. Microbiol. Biotechnol. 2001, 56, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Hureiki, L.; Croué, J.P.; Legube, B. Chlorination studies of free and combined amino acids. Water Res. 1994, 28, 2521–2531. [Google Scholar] [CrossRef]
- Pehlivanoglu-Mantas, E.; Sedlak, D.L. Wastewater-Derived Dissolved Organic Nitrogen: Analytical Methods, Characterization, and Effects—A Review. Crit. Rev. Environ. Sci. Technol. 2006, 36, 261–285. [Google Scholar] [CrossRef]
- Huo, S.; Xi, B.; Yu, H.; Qin, Y.; Zan, F.; Zhang, J. Characteristics and transformations of dissolved organic nitrogen in municipal biological nitrogen removal wastewater treatment plants. Environ. Res. Lett. 2013, 8, 044005. [Google Scholar] [CrossRef]
- Chen, J.; Xu, J.; Zhang, S.; Liu, F.; Peng, J.; Peng, Y.; Wu, J. Nitrogen removal characteristics of a novel heterotrophic nitrification and aerobic denitrification bacteria, Alcaligenes faecalis strain WT14. J. Environ. Manage. 2021, 282, 111961. [Google Scholar] [CrossRef]
- Rout, P.R.; Bhunia, P.; Dash, R.R. Simultaneous removal of nitrogen and phosphorous from domestic wastewater using Bacillus cereus GS-5 strain exhibiting heterotrophic nitrification, aerobic denitrification and denitrifying phosphorous removal. Bioresour. Technol. 2017, 244, 484–495. [Google Scholar] [CrossRef]
- Xu, N.; Liao, M.; Liang, Y.; Guo, J.; Zhang, Y.; Xie, X.; Fan, Q.; Zhu, Y. Biological nitrogen removal capability and pathways analysis of a novel low C/N ratio heterotrophic nitrifying and aerobic denitrifying bacterium (Bacillus thuringiensis strain WXN-23). Environ. Res. 2021, 195, 110797. [Google Scholar] [CrossRef]
- Ebeling, J.M.; Timmons, M.B.; Bisogni, J.J. Engineering analysis of the stoichiometry of photoautotrophic, autotrophic, and heterotrophic removal of ammonia–nitrogen in aquaculture systems. Aquaculture 2006, 257, 346–358. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Zhao, B.; An, Q.; Huang, Y.S. Nitrogen removal by Providencia rettgeri strain YL with heterotrophic nitrification and aerobic denitrification. Environ. Technol. 2016, 37, 2206–2213. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Jung, J.Y.; Han, H.S. Utilization of Microalgae in Aquaculture System: Biological Wastewater Treatment. Emerg. Sci. J. 2019, 3, 209–221. [Google Scholar] [CrossRef]
- Lu, L.; Wang, G.; Yeung, M.; Xi, J.; Hu, H.-Y. Shift of microbial community in gas-phase biofilters with different inocula, inlet loads and nitrogen sources. Process Biochem. 2020, 91, 57–64. [Google Scholar] [CrossRef]
- Schilling, O.; Frick, O.; Herzberg, C.; Ehrenreich, A.; Heinzle, E.; Wittmann, C.; Stulke, J. Transcriptional and metabolic responses of Bacillus subtilis to the availability of organic acids: Transcription regulation is important but not sufficient to account for metabolic adaptation. Appl. Environ. Microbiol. 2007, 73, 499–507. [Google Scholar] [CrossRef] [Green Version]
- Lins Rodigues, M.; Damasceno, D.Z.; Gomes, R.L.M.; Sosa, B.d.S.; Moro, E.B.; Boscolo, W.R.; Bittencourt, F.; Signor, A. Probiotic effects (Bacillus cereus and Bacillus subtilis) on growth and physiological parameters of silver catfish (Rhamdia quelen). Aquac. Nutr. 2020, 27, 454–467. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Xing, K.; Jiang, P.; Wang, J. Dietary cinnamaldehyde and Bacillus subtilis improve growth performance, digestive enzyme activity, and antioxidant capability and shape intestinal microbiota in tongue sole, Cynoglossus semilaevis. Aquaculture 2021, 531, 735798. [Google Scholar] [CrossRef]
- Zhou, S.; Song, D.; Zhou, X.; Mao, X.; Zhou, X.; Wang, S.; Wei, J.; Huang, Y.; Wang, W.; Xiao, S.M.; et al. Characterization of Bacillus subtilis from gastrointestinal tract of hybrid Hulong grouper (Epinephelus fuscoguttatus x E. lanceolatus) and its effects as probiotic additives. Fish Shellfish Immunol. 2019, 84, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Park, Y.; Je, H.W.; Seong, M.; Damusaru, J.H.; Kim, S.; Jung, J.Y.; Bai, S.C. Tuna byproducts as a fish-meal in tilapia aquaculture. Ecotoxicol. Environ. Saf. 2019, 172, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Robinson, E.H.; Jackson, L.S.; Li, M.H.; Kingsbury, S.K.; Tucker, C.S. Effect of Time of Feeding on Growth of Channel Catfish. J. World Aquacult. Soc. 1995, 26, 320–322. [Google Scholar] [CrossRef]
- Hallier, A.; Serot, T.; Prost, C. Influence of rearing conditions and feed on the biochemical composition of fillets of the European catfish (Silurus glanis). Food Chem. 2007, 103, 808–815. [Google Scholar] [CrossRef]
- Jiang, J.; Xu, S.; Feng, L.; Liu, Y.; Jiang, W.; Wu, P.; Wang, Y.; Zhao, Y.; Zhou, X. Lysine and methionine supplementation ameliorates high inclusion of soybean meal inducing intestinal oxidative injury and digestive and antioxidant capacity decrease of yellow catfish. Fish Physiol. Biochem. 2018, 44, 319–328. [Google Scholar] [CrossRef]
- Mondal, K.; Kaviraj, A.; Mukhopadhyay, P.K. Evaluation of fermented fish-offal in the formulated diet of the freshwater catfish Heteropneustes fossilis. Aquac. Res. 2008, 39, 1443–1449. [Google Scholar] [CrossRef]
- Wang, F.; Ma, X.; Wang, W.; Liu, J. Comparison of proximate composition, amino acid and fatty acid profiles in wild, pond- and cage-cultured longsnout catfish (Leiocassis longirostris). Int. J. Food Sci. Technol. 2012, 47, 1772–1776. [Google Scholar] [CrossRef]
- Eddy, F.B.; Williams, E.M. Nitrite and Freshwater Fish. Chem. Ecol. 1987, 3, 1–38. [Google Scholar] [CrossRef]
- Ip, Y.K.; Chew, S.F.; Leong, I.A.; Jin, Y.; Lim, C.B.; Wu, R.S. The sleeper Bostrichthys sinensis (family Eleotridae) stores glutamine and reduces ammonia production during aerial exposure. J. Comp. Physiol. B 2001, 171, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Svobodova, Z.; Lloyd, R.; Machova, J. Water Quality and Fish Health; Food and Agriculture Organization of the United Nations: Roma, Italy, 1993. [Google Scholar]
- Kim, Y.-S.; Kim, S.-E.; Kim, S.-J.; Jung, H.-K.; Park, J.; Jeon, Y.J.; Kim, D.-H.; Kang, J.-H.; Kim, K.-H. Effects of wheat flour and culture period on bacterial community composition in digestive tracts of Litopenaeus vannamei and rearing water in biofloc aquaculture system. Aquaculture 2021, 531, 735908. [Google Scholar] [CrossRef]
- Liu, W.; Du, X.; Tan, H.; Xie, J.; Luo, G.; Sun, D. Performance of a recirculating aquaculture system using biofloc biofilters with convertible water-treatment efficiencies. Sci. Total Environ. 2021, 754, 141918. [Google Scholar] [CrossRef]
- Chang, Y.C.; Ma, C.H.; Lee, H.T.; Hsu, T.H. Polyculture of Juvenile Dog Conch Laevistrombus canarium Reveals High Potentiality in Integrated Multitrophic Aquaculture (IMTA). Biology 2021, 10, 812. [Google Scholar] [CrossRef]
- Lobanov, V.P.; Combot, D.; Pelissier, P.; Labbe, L.; Joyce, A. Improving Plant Health Through Nutrient Remineralization in Aquaponic Systems. Front Plant Sci. 2021, 12, 683690. [Google Scholar] [CrossRef] [PubMed]
- Gorito, A.M.; Lado Ribeiro, A.R.; Pereira, M.F.R.; Almeida, C.M.R.; Silva, A.M.T. Advanced oxidation technologies and constructed wetlands in aquaculture farms: What do we know so far about micropollutant removal? Environ. Res. 2021, 204, 111955. [Google Scholar] [CrossRef] [PubMed]
- Giraud, E.; Douet, D.G.; Le Bris, H.; Bouju-Albert, A.; Donnay-Moreno, C.; Thorin, C.; Pouliquen, H. Survey of antibiotic resistance in an integrated marine aquaculture system under oxolinic acid treatment. FEMS Microbiol. Ecol. 2006, 55, 439–448. [Google Scholar] [CrossRef] [Green Version]
- Pagand, P.; Blancheton, J.-P.; Lemoalle, J.; Casellas, C. The use of high rate algal ponds for the treatment of marine effluent from a recirculating fish rearing system. Aquac. Res. 2000, 31, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Azim, M.E.; Wahab, M.A.; Dam, A.A.v.; Beveridge, M.C.M.; Huisman, E.A.; Verdegem, M.C.J. Optimization of stocking ratios of two Indian major carps, rohu (Labeo rohita Ham.) and catla (Catla catla Ham.) in a periphyton-based aquaculture system. Aquaculture 2001, 203, 33–49. [Google Scholar] [CrossRef]
- Brune, D.E.; Schwartz, G.; Eversole, A.G.; Collier, J.A.; Schwedler, T.E. Partitioned Aquaculture Systems. In Biology and Culture of Channel Catfish; Tucker, C.S., Hargreaves, J.A., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2004; pp. 561–584. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. F 6 (Con) | B.subtilis (Semi-control) | B. albus | B. safensis | P. mariniglutinosa | P. celer | |
|---|---|---|---|---|---|---|
| Initial mean weight (g fish−1) | 34.2 ± 0.1 | 34.1 ± 0.1 | 34.0 ± 0.1 | 34.1 ± 0.1 | 34.2 ± 0.1 | 34.1 ± 0.1 |
| Final mean weight (g fish−1) | 75.3 ± 1.5 c | 79.8 ± 1.1 b | 81.7 ± 1.7 b | 81.1 ± 1.6 b | 92.8 ± 2.5 a | 91.3 ± 2.5 a |
| WG (%) 2 | 120.2 ± 4.5 d | 134.0 ± 3.3 c | 140.3 ± 5.0 b | 137.8 ± 4.6 bc | 171. ± 7.2 a | 167.7 ± 7.3 a |
| FE (%) 3 | 123.3 ± 2.1 c | 137.1 ± 5.4 b | 143.1 ± 3.8 b | 141.0 ± 6.6 b | 175.8 ± 5.3 a | 171.6 ± 7.1 a |
| SGR (%/day) 4 | 1.41 ± 0.04 d | 1.52 ± 0.03 c | 1.57 ± 0.04 b | 1.55 ± 0.03 bc | 1.78 ± 0.05 a | 1.76 ± 0.05 a |
| Survival (%) 5 | 100 | 100 | 100 | 100 | 100 | 100 |
| Microorganisms (a) 2 | Amur Catfish (b) 3 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| B. subtilis | B. albus | B. safensis | P. mariniglutinosa | P. celer | C.F 4 (Con) | B. subtilis (Semi-control) | B. albus | B. safensis | P. mariniglutinosa | P. celer | |
| Moisture | 1.26 a | 1.39 a | 1.28 a | 0.16 b | 0.11 b | 76.03 b | 75.94 b | 75.76 b | 77.84 a | 78.52 a | 75.01 b |
| Protein | 58.36 c | 42.05 e | 56.81 d | 70.61 a | 68.34 b | 15.57 b | 15.45 b | 15.41 b | 14.10 c | 14.08 c | 16.35 a |
| Lipid | 0.56 a | 0.44 b | 0.48 ab | 0.43 b | 0.33 c | 6.25 b | 5.23 c | 6.26 b | 5.37 c | 5.57 c | 6.82 a |
| Microorganisms (a) | Amur Catfish (b) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| B. subtilis | B. albus | B. safensis | P. mariniglutinosa | P. celer | C.F 3 | B. subtilis | B. albus | B. safensis | P. mariniglutinosa | P. celer | |
| Essential amino acids (EAA) | |||||||||||
| Arginine | 2.50 cd | 2.29 d | 3.43 a | 2.80 b | 2.52 c | 0.98 bc | 0.97 bcd | 1.07 a | 0.89 d | 0.91 cd | 1.02 ab |
| Threonine | 2.05 b | 1.53 c | 2.02 b | 2.25 a | 2.02 b | 0.69 a | 0.65 ab | 0.66 ab | 0.61 bc | 0.56 c | 0.68 a |
| Valine | 2.96 c | 2.13 e | 2.56 d | 3.87 a | 3.54 b | 0.73 a | 0.71 a | 0.71 a | 0.64 b | 0.57 c | 0.73 a |
| Isoleucine | 2.30 c | 1.84 d | 2.21 c | 2.92 a | 2.64 b | 0.66 a | 0.66 a | 0.66 a | 0.58 b | 0.53 b | 0.66 a |
| Leucine | 3.44 bc | 2.55 d | 3.19 c | 3.76 a | 3.47 b | 1.15 a | 1.11 a | 1.14 a | 1.01 b | 0.93 b | 1.19 a |
| Methionine | 0.65 c | 0.80 b | 1.19 a | 0.67 c | 0.46 d | 2.57 a | 1.86 b | 1.36 c | 1.39 c | 1.84 b | 1.42 c |
| Lysine | 3.35 b | 2.63 c | 3.13 b | 3.91 a | 3.81 a | 1.26 a | 1.25 a | 1.25 a | 1.10 b | 1.01 b | 1.27 a |
| Phenylalanine | 2.15 b | 1.47 d | 1.90 c | 2.33 a | 2.12 b | 0.65 a | 0.63 a | 0.64 a | 0.56 b | 0.53 b | 0.66 a |
| Histidine | 1.91 d | 2.73 c | 2.64 c | 3.30 b | 4.44 a | 0.50 b | 0.51 ab | 0.49 b | 0.44 c | 0.41 c | 0.54 a |
| Total | 21.31 b | 17.97 c | 22.27 b | 25.81 a | 25.02 a | 9.19 a | 8.35 b | 7.98 b | 7.22 c | 7.29 c | 8.17 b |
| Non-essential amino acids (NEAA) | |||||||||||
| Serine | 1.87 a | 1.17 d | 1.65 b | 1.65 b | 1.47 c | 0.71 ab | 0.66 bcd | 0.68 abc | 0.63 cd | 0.61 d | 0.73 a |
| Glutamic acid | 7.24 c | 6.16 d | 7.53 c | 9.63 a | 8.48 b | 2.16 ab | 2.12 bc | 2.16 ab | 1.96 cd | 1.86 d | 2.31 a |
| Proline | 2.15 b | 1.47 c | 1.52 c | 2.42 a | 2.50 a | 0.87 b | 0.78 c | 0.86 b | 0.83 bc | 0.97 a | 0.98 a |
| Glycine | 2.40 b | 1.58 d | 2.15 c | 2.62 a | 2.41 b | 1.17 b | 1.16 b | 1.06 c | 1.16 b | 1.34 a | 1.20 b |
| Alanine | 4.17 b | 2.96 c | 3.29 c | 5.50 a | 5.17 a | 1.04 a | 1.00 ab | 0.94 b | 0.94 b | 0.97 ab | 1.03 a |
| Tyrosine | 1.24 b | 1.18 b | 1.44 a | 1.38 a | 1.19 b | 0.52 ab | 0.48 bc | 0.52 ab | 0.45 cd | 0.41 d | 0.53 a |
| Aspartic acid | 4.68 a | 3.57 c | 5.00 a | 4.15 b | 4.02 b | 1.50 a | 1.48 a | 1.48 a | 1.33 b | 1.25 b | 1.55 a |
| Cysteine | 0.71 b | 0.57 c | 0.80 a | 0.40 e | 0.49 d | 0.13 | 0.10 | 0.10 | 0.09 | 0.09 | 0.09 |
| Total | 24.46 bc | 18.66 d | 23.38 c | 27.75 a | 25.73 b | 8.10 ab | 7.78 abc | 7.80 abc | 7.39 c | 7.50 bc | 8.42 a |
| Microorganisms (a) | Amur Catfish (b) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| B. subtilis | B. albus | B. safensis | P. mariniglutinosa | P. celer | C.F 6 | B. subtilis | B. albus | B. safensis | P. mariniglutinosa | P. celer | |
| C12:0 | 2.65 b | 3.44 a | 1.27 d | 0.34 e | 1.85 c | 0.18 a | 0.13 b | 0.13 b | 0.11 b | 0.10 b | 0.11 b |
| C14:0 | 4.26 e | 5.05 d | 11.14 b | 5.94 c | 13.60 a | 5.04 a | 4.74 ab | 4.38 bc | 4.00 d | 4.01 d | 4.22 cd |
| C14:1 | 0.06 | 0.06 | 0.07 | 0.06 | 0.05 | 0.06 | |||||
| C16:0 | 42.07 c | 48.08 b | 51.65 ab | 41.33 c | 52.67 a | 37.16 | 37.70 | 38.27 | 36.08 | 38.34 | 37.95 |
| C16:1 | 2.24 b | 1.47 c | 3.35 a | 1.51 c | 3.22 a | 3.76 b | 4.07 ab | 4.36 a | 3.86 b | 3.91 b | 3.89 b |
| C18:0 | 11.59 c | 13.86 a | 13.73 a | 12.37 bc | 12.93 ab | 9.90 b | 9.96 b | 10.36 b | 10.02 b | 11.22 a | 10.72 ab |
| C18:1n9 | 19.15 a | 16.66 b | 10.84 d | 15.24 c | 7.87 e | 18.17 | 18.40 | 18.63 | 17.68 | 18.66 | 18.95 |
| C18:2n6 | 15.43 b | 9.85 c | 3.92 d | 18.70 a | 3.63 d | 13.82 a | 13.79 ab | 13.55 ab | 12.71 b | 13.01 ab | 13.64 ab |
| C18:3n3 | 1.15 | 0.65 | 1.93 | 2.25 a | 2.14 a | 1.97 b | 1.85 b | 1.89 b | 2.15 a | ||
| C18:3n6 | 0.23 c | 0.25 bc | 0.29 a | 0.26 b | 0.25 bc | 0.26 b | |||||
| C20:0 | 0.34 | 0.35 bc | 0.36 bc | 0.36 bc | 0.34 c | 0.44 a | 0.38 b | ||||
| C20:3n3 | 0.13 a | 0.11 ab | 0.09 bc | 0.08 c | 0.09 bc | 0.10 bc | |||||
| C20:5n3 | 0.51 | 0.95 | 1.00 a | 0.87 b | 0.71 de | 0.67 e | 0.76 cd | 0.79 c | |||
| C22:1n9 | 0.13 ab | 0.14 ab | 0.13 ab | 0.11 b | 0.15 a | 0.12 ab | |||||
| C22:2 | 0.05 a | 0.04 ab | 0.03 b | 0.03 b | 0.04 ab | 0.04 ab | |||||
| C22:6n3 | 0.23 | 0.40 | 2.98 a | 2.63 b | 2.22 de | 2.12 e | 2.44 bc | 2.33 cd | |||
| C24:0 | 0.05 ab | 0.05 ab | 0.05 ab | 0.04 b | 0.07 a | 0.05 ab | |||||
| SFA 3 | 61.64 c | 71.37 b | 81.79 a | 61.42 c | 83.37 a | 54.07 | 54.28 | 54.82 | 51.78 | 55.55 | 54.63 |
| MUFA 4 | 21.39 a | 18.13 b | 14.29 c | 17.21 b | 11.65 d | 23.85 | 24.37 | 24.90 | 23.30 | 24.51 | 24.69 |
| PUFA 5 | 16.97 b | 10.50 c | 3.92 d | 21.37 a | 4.98 d | 22.08 b | 21.35 bc | 20.28 c | 24.92 a | 19.94 c | 20.68 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.; Jung, J.-Y.; Kwon, J.-H. Potential of Bacterial Strains Isolated from Coastal Water for Wastewater Treatment and as Aqua-Feed Additives. Microorganisms 2021, 9, 2441. https://doi.org/10.3390/microorganisms9122441
Kim K, Jung J-Y, Kwon J-H. Potential of Bacterial Strains Isolated from Coastal Water for Wastewater Treatment and as Aqua-Feed Additives. Microorganisms. 2021; 9(12):2441. https://doi.org/10.3390/microorganisms9122441
Chicago/Turabian StyleKim, Kyochan, Joo-Young Jung, and Jong-Hee Kwon. 2021. "Potential of Bacterial Strains Isolated from Coastal Water for Wastewater Treatment and as Aqua-Feed Additives" Microorganisms 9, no. 12: 2441. https://doi.org/10.3390/microorganisms9122441