
Mobile Colistin Resistance Genetic Determinants of Non-Typhoid Salmonella enterica Isolates from Russia

 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Identification
2.2. Antibiotic Susceptibility Testing
2.3. Real-Time PCR Assays for mcr-1
2.4. Whole-Genome Sequencing and Assembling
2.5. Bioinformatic Analysis
3. Results
3.1. Isolates Description and Antimicrobial Susceptibility Testing
3.2. Multilocus Sequence Typing and WGS-Typing
3.3. Related Genetic Determinants of Salmonella Isolates to Antimicrobial Susceptibility
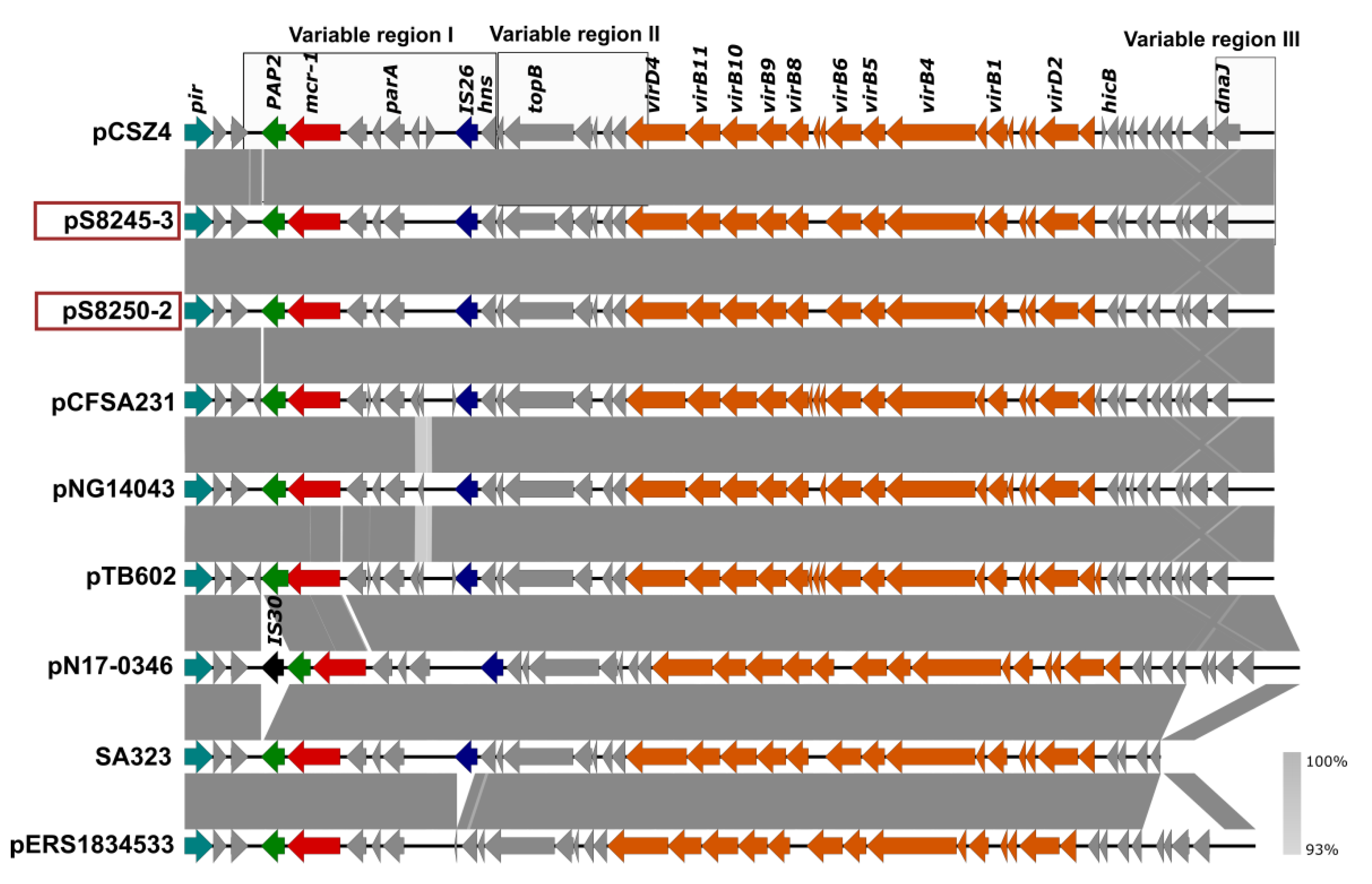
3.3.1. Salmonella Enteritidis SLR1_8250 and SLR1_8245
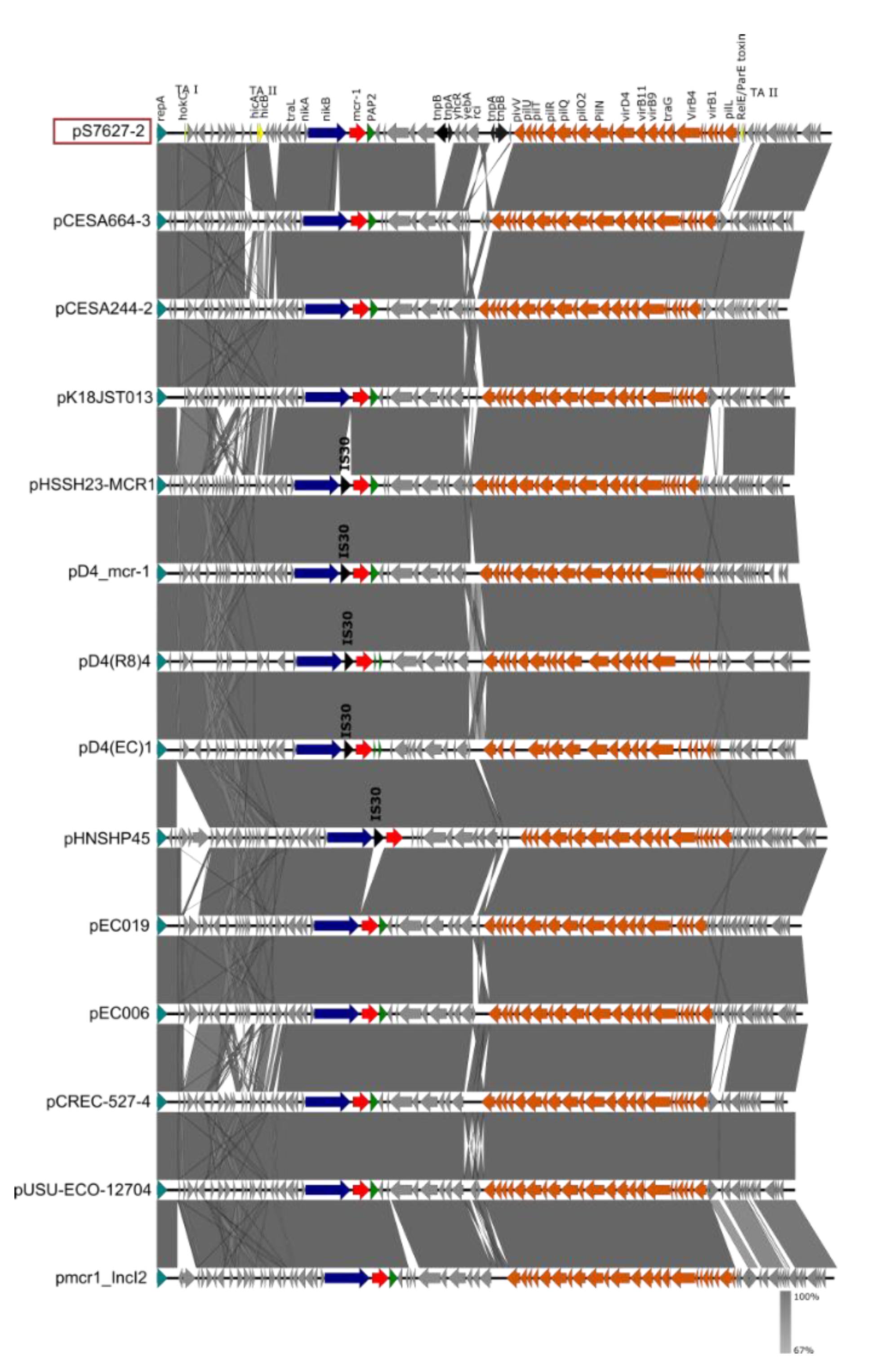
3.3.2. Salmonella Bovismorbificans SLR1_7627
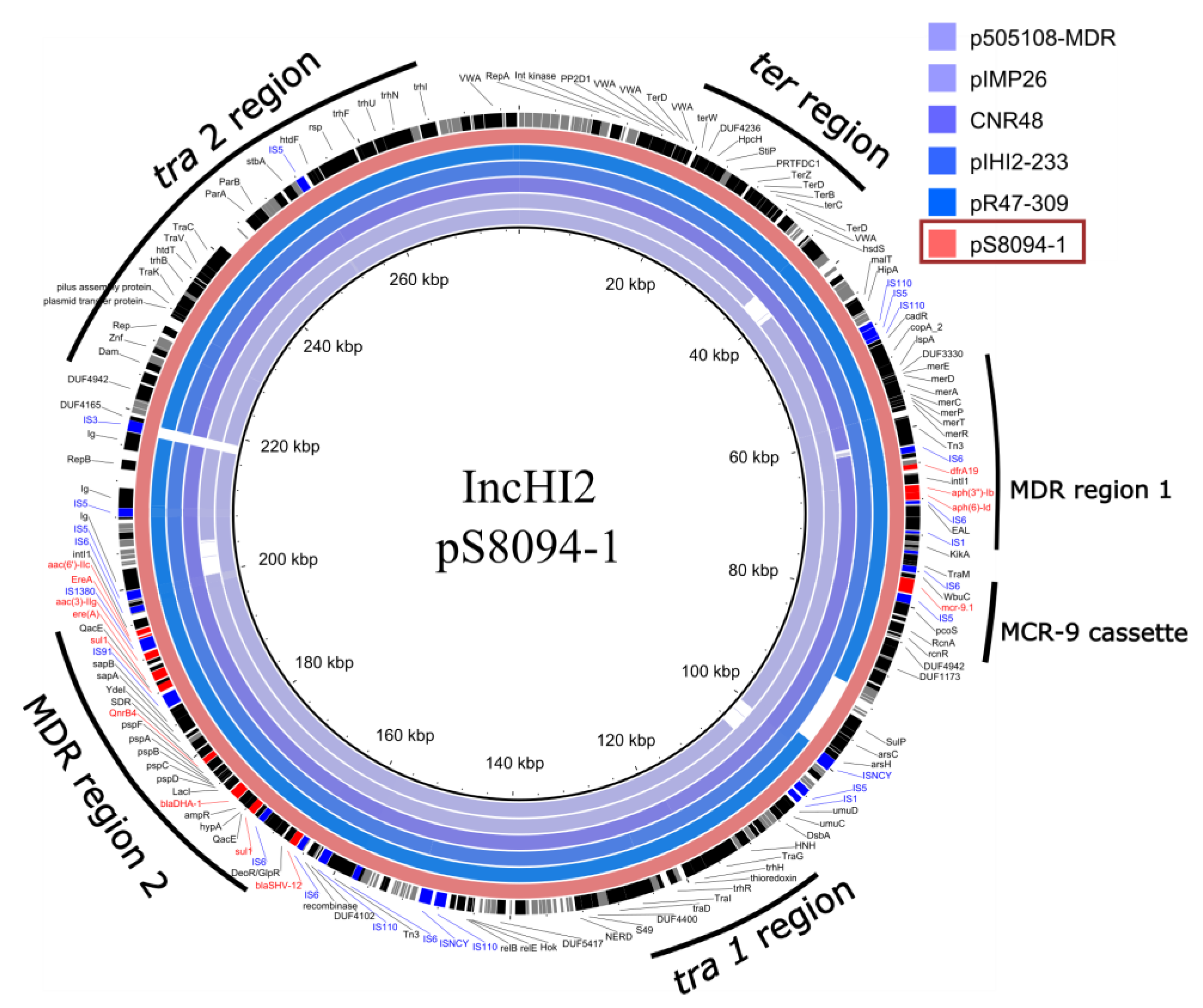
3.3.3. Monophasic Variant of Salmonella Typhimurium 4,5:i:- (SLR1_8094)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Centers for Disease Control and Prevention. National Salmonella Surveillance Annual Report, 2011. Atlanta, Georgia: US Department of Health and Human Services, CDC. 2013. Available online: https://www.cdc.gov/ncezid/dfwed/PDFs/salmonella-annual-report-2011-508c.pdf (accessed on 3 December 2021).
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M.; International Collaboration on Enteric Disease “Burden of Illness” Studies. The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [Green Version]
- State Report on the State of Sanitary and Epidemiological Well-Being of the Population in the Russian Federation in 2020. Available online: https://www.rospotrebnadzor.ru/bitrix/redirect.php?event1=file&event2=download&event3=gd-seb_02.06-_s-podpisyu_.pdf&goto=/upload/iblock/5fa/gd-seb_02.06-_s-podpisyu_.pdf (accessed on 1 November 2021). (In Russian).
- Rozhnova, S.S.; Kuleshov, K.V.; Pavlova, A.S.; Guseva, A.N.; Kozhakhmetova, T.A.; Akulova, N.K.; Podkolzin, A.T. Heterogeneity of Salmonella isolates obtained from various sources in Russian Federation 2010–2019. Epidemiol. Infect. Dis. 2020, 25, 26–34. (In Russian) [Google Scholar] [CrossRef]
- Caniaux, I.; van Belkum, A.; Zambardi, G.; Poirel, L.; Gros, M.F. MCR: Modern colistin resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 415–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; van Dorp, L.; Shaw, L.P.; Bradley, P.; Wang, Q.; Wang, X.; Jin, L.; Zhang, Q.; Liu, Y.; Rieux, A.; et al. The global distribution and spread of the mobilized colistin resistance gene mcr-1. Nat. Commun. 2018, 9, 1179. [Google Scholar] [CrossRef] [Green Version]
- Lima, T.; Domingues, S.; Da Silva, G.J. Plasmid-Mediated Colistin Resistance in Salmonella enterica: A Review. Microorganisms 2019, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Scott, H.M.; Acuff, G.; Bergeron, G.; Bourassa, M.W.; Gill, J.; Graham, D.W.; Kahn, L.H.; Morley, P.S.; Salois, M.J.; Simjee, S.; et al. Critically important antibiotics: Criteria and approaches for measuring and reducing their use in food animal agriculture. Ann. N. Y. Acad. Sci. 2019, 1441, 8–16. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.B.; Dong, B.L.; Huang, X.H.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [Green Version]
- Vallejos-Sanchez, K.; Tataje-Lavanda, L.; Villanueva-Perez, D.; Bendezu, J.; Montalvan, A.; Zimic-Peralta, M.; Fernandez-Sanchez, M.; Fernandez-Diaz, M. Whole-Genome Sequencing of a Salmonella enterica subsp. enterica Serovar Infantis Strain Isolated from Broiler Chicken in Peru. Microbiol. Resour. Announc. 2019, 8, e00826-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, N.; Hughes, V.M. Plasmids of the same Inc groups in Enterobacteria before and after the medical use of antibiotics. Nature 1983, 306, 616–617. [Google Scholar] [CrossRef] [PubMed]
- Matamoros, S.; van Hattem, J.M.; Arcilla, M.S.; Willemse, N.; Melles, D.C.; Penders, J.; Vinh, T.N.; Hoa, N.T.; Bootsma, M.C.J.; van Genderen, P.J.; et al. Global phylogenetic analysis of Escherichia coli and plasmids carrying the mcr-1 gene indicates bacterial diversity but plasmid restriction. Sci. Rep. 2017, 7, 15364. [Google Scholar] [CrossRef] [Green Version]
- Doumith, M.; Godbole, G.; Ashton, P.; Larkin, L.; Dallman, T.; Day, M.; Day, M.; Muller-Pebody, B.; Ellington, M.J.; de Pinna, E.; et al. Detection of the plasmid-mediated mcr-1 gene conferring colistin resistance in human and food isolates of Salmonella enterica and Escherichia coli in England and Wales. J. Antimicrob. Chemother. 2016, 71, 2300–2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manageiro, V.; Clemente, L.; Romao, R.; Silva, C.; Vieira, L.; Ferreira, E.; Canica, M. IncX4 Plasmid Carrying the New mcr-1.9 Gene Variant in a CTX-M-8-Producing Escherichia coli Isolate Recovered from Swine. Front. Microbiol. 2019, 10, 367. [Google Scholar] [CrossRef]
- Ye, H.; Li, Y.; Li, Z.; Gao, R.; Zhang, H.; Wen, R.; Gao, G.F.; Hu, Q.; Feng, Y. Diversified mcr-1-Harbouring Plasmid Reservoirs Confer Resistance to Colistin in Human Gut Microbiota. MBio 2016, 7, e00177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Di-ameters. Version 11.0, valid from 1 January 2021. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_11.0_Breakpoint_Tables.pdf (accessed on 29 November 2021).
- EUCAST. Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. Version 2.0. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Resistance_mechanisms/EUCAST_detection_of_resistance_mechanisms_170711.pdf (accessed on 29 November 2021).
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seibt, K.M.; Schmidt, T.; Heitkam, T. FlexiDot: Highly customizable, ambiguity-aware dotplots for visual sequence analyses. Bioinformatics 2018, 34, 3575–3577. [Google Scholar] [CrossRef]
- Achtman, M.; Wain, J.; Weill, F.X.; Nair, S.; Zhou, Z.M.; Sangal, V.; Krauland, M.G.; Hale, J.L.; Harbottle, H.; Uesbeck, A.; et al. Multilocus Sequence Typing as a Replacement for Serotyping in Salmonella enterica. PLoS Pathog. 2012, 8, e1002776. [Google Scholar] [CrossRef] [Green Version]
- Robertson, J.; Nash, J.H.E. MOB-suite: Software tools for clustering, reconstruction and typing of plasmids from draft assemblies. Microb. Genom. 2018, 4, e000206. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In Silico Detection and Typing of Plasmids Using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.F.; Petty, N.K.; Zakour, N.L.B.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Fang, L.X.; Wu, Z.; Deng, H.; Yang, R.S.; Li, X.P.; Li, S.M.; Liao, X.P.; Feng, Y.; Liu, Y.H. Genetic Analysis of the IncX4 Plasmids: Implications for a Unique Pattern in the mcr-1 Acquisition. Sci. Rep. 2017, 7, 424. [Google Scholar] [CrossRef]
- Hu, Y.; Nguyen, S.V.; Wang, W.; Gan, X.; Dong, Y.; Liu, C.; Cui, X.; Xu, J.; Li, F.; Fanning, S. Antimicrobial Resistance and Genomic Characterization of Two mcr-1-Harboring Foodborne Salmonella Isolates Recovered in China, 2016. Front. Microbiol. 2021, 12, 636284. [Google Scholar] [CrossRef]
- Tang, B.; Chang, J.; Zhang, L.; Liu, L.; Xia, X.; Hassan, B.H.; Jia, X.; Yang, H.; Feng, Y. Carriage of Distinct mcr-1-Harboring Plasmids by Unusual Serotypes of Salmonella. Adv. Biosyst. 2020, 4, e1900219. [Google Scholar] [CrossRef]
- Moon, D.C.; Kim, S.J.; Mechesso, A.F.; Kang, H.Y.; Song, H.J.; Choi, J.H.; Yoon, S.S.; Lim, S.K. Mobile Colistin Resistance Gene mcr-1 Detected on an IncI2 Plasmid in Salmonella Typhimurium Sequence Type 19 from a Healthy Pig in South Korea. Microorganisms 2021, 9, 398. [Google Scholar] [CrossRef]
- Shi, L.; Liang, Q.; Zhan, Z.; Feng, J.; Zhao, Y.; Chen, Y.; Huang, M.; Tong, Y.; Wu, W.; Chen, W.; et al. Co-occurrence of 3 different resistance plasmids in a multi-drug resistant Cronobacter sakazakii isolate causing neonatal infections. Virulence 2018, 9, 110–120. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Chen, Q.; Qian, C.; Shen, K.; Zhu, X.; Zhou, D.; Lu, W.; Sun, Z.; Liu, H.; Li, K.; et al. In Vitro Susceptibility and Florfenicol Resistance in Citrobacter Isolates and Whole-Genome Analysis of Multidrug-Resistant Citrobacter freundii. Int. J. Genom. 2019, 2019, 7191935. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Wannemeuhler, Y.M.; Scaccianoce, J.A.; Johnson, S.J.; Nolan, L.K. Complete DNA sequence, comparative genomics, and prevalence of an IncHI2 plasmid occurring among extraintestinal pathogenic Escherichia coli isolates. Antimicrob. Agents Chemother. 2006, 50, 3929–3933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Dai, X.; Zeng, J.; Gao, Y.; Zhang, Z.; Zhang, L. Characterization of the global distribution and diversified plasmid reservoirs of the colistin resistance gene mcr-9. Sci. Rep. 2020, 10, 8113. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Zhang, J.; Gu, Z.; Li, R.; Chan, E.W.; Yan, M.; Wu, C.; Xu, X.; Chen, S. Prevalence and Molecular Characterization of mcr-1-Positive Salmonella Strains Recovered from Clinical Specimens in China. Antimicrob. Agents Chemother. 2017, 61, e02471-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sia, C.M.; Greig, D.R.; Day, M.; Hartman, H.; Painset, A.; Doumith, M.; Meunier, D.; Jenkins, C.; Chattaway, M.A.; Hopkins, K.L.; et al. The characterization of mobile colistin resistance (mcr) genes among 33,000 Salmonella enterica genomes from routine public health surveillance in England. Microb. Genom. 2020, 6, e000331. [Google Scholar] [CrossRef]
- Cooper, A.L.; Low, A.J.; Koziol, A.G.; Thomas, M.C.; Leclair, D.; Tamber, S.; Wong, A.; Blais, B.W.; Carrillo, C.D. Systematic Evaluation of Whole Genome Sequence-Based Predictions of Salmonella Serotype and Antimicrobial Resistance. Front. Microbiol. 2020, 11, 549. [Google Scholar] [CrossRef] [Green Version]
- Migura-Garcia, L.; Gonzalez-Lopez, J.J.; Martinez-Urtaza, J.; Sanchez, J.R.A.; Moreno-Mingorance, A.; Rozas, A.P.d.; Hofle, U.; Ramiro, Y.; Gonzalez-Escalona, N. mcr-Colistin Resistance Genes Mobilized by IncX4, IncHI2, and IncI2 Plasmids in Escherichia coli of Pigs and White Stork in Spain. Front. Microbiol. 2019, 10, 3072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Yi, L.X.; Yu, L.F.; Wang, J.; Liu, Y.; Chen, X.; Lv, L.; Yang, J.; Liu, J.H. Fitness Advantage of mcr-1-Bearing IncI2 and IncX4 Plasmids In Vitro. Front. Microbiol. 2018, 9, 331. [Google Scholar] [CrossRef]
- Lo, W.U.; Chow, K.H.; Law, P.Y.; Ng, K.Y.; Cheung, Y.Y.; Lai, E.L.; Ho, P.L. Highly conjugative IncX4 plasmids carrying blaCTX-M in Escherichia coli from humans and food animals. J. Med. Microbiol. 2014, 63, 835–840. [Google Scholar] [CrossRef]
- Rodrigue, D.C.; Tauxe, R.V.; Rowe, B. International increase in Salmonella enteritidis: A new pandemic? Epidemiol. Infect. 1990, 105, 21–27. [Google Scholar] [CrossRef]
- Christenson, J.C. Salmonella infections. Pediatr. Rev. 2013, 34, 375–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Zeng, M.; Xu, J.; Zhou, H.; Gu, B.; Li, Z.; Jin, H.; Wang, X.; Zhang, W.; Hu, Y.; et al. Epidemiologic and genomic insights on mcr-1-harbouring Salmonella from diarrhoeal outpatients in Shanghai, China, 2006–2016. EBioMedicine 2019, 42, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of Novel Mobilized Colistin Resistance Gene mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella enterica Serotype Typhimurium Isolate. MBio 2019, 10, e00853-19. [Google Scholar] [CrossRef] [Green Version]
- Tyson, G.H.; Li, C.; Hsu, C.H.; Ayers, S.; Borenstein, S.; Mukherjee, S.; Tran, T.T.; McDermott, P.F.; Zhao, S. The mcr-9 Gene of Salmonella and Escherichia coli Is Not Associated with Colistin Resistance in the United States. Antimicrob. Agents Chemother. 2020, 64, e00573-20. [Google Scholar] [CrossRef]
- Cha, M.H.; Woo, G.J.; Lee, W.; Kim, S.H.; Woo, J.H.; Kim, J.; Ryu, J.G.; Kwak, H.S.; Chi, Y.M. Emergence of Transferable mcr-9 Gene-Carrying Colistin-Resistant Salmonella enterica Dessau ST14 Isolated from Retail Chicken Meat in Korea. Foodborne Pathog. Dis. 2020, 17, 720–727. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Z.; Lyu, N.; Ma, S.; Liu, F.; Hu, Y.; Gao, G.F.; Zhu, B. Comparative genomic analysis of mobile colistin resistance gene mcr-9 in Salmonella enterica. J. Infect. 2021, 82, e15–e17. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Serotype | Isolation Date (mm/yyyy) | Country:City | Epidemiological Background | Source | mcr-Type | |
|---|---|---|---|---|---|---|
| SLR1_8250 | S. Enteritidis | 04/2019 | Russia:Yakutiya | ND | food (chicken meat) | 1 |
| SLR1_8245 | S. Enteritidis | 07/2019 | Russia:El’ban | Outbreak | human | 1 |
| SLR1_7627 | S. Bovismorbificans | 09/2018 | Russia:Irkutsk | Sporadic | human | 1 |
| SLR1_8094 | S. Typhimurium monophasic variant [4,5:i:-] | 06/2019 | Russia:Ulan-Ude | Sporadic | human | 9 |
| Antimicrobial Class | Antimicrobial Agent | SLR1_8250 | SLR1_7627 | SLR1_8245 | SLR1_8094 | ||||
|---|---|---|---|---|---|---|---|---|---|
| polymyxins | colistin | 8 (R) | mcr-1.1 | 8 (R) | mcr-1.1 | 8 (R) | mcr-1.1 | 1 (S) | mcr-9 |
| penicillins | ampicillin | 2 (S) | >128 (R) | blaTEM-1B | 2 (S) | >128 (R) | blaDHA-1, blaSHV-12 | ||
| ampicillin-sulbactam | 2/1 (S) | 32/8 (R) | 2/1 (S) | 64/32 (R) | |||||
| piperacillin | 2 (S) | >128 (R) | 2 (S) | >128 (R) | |||||
| piperacillin-tazobactam | ≤2/4 (S) | ≤1/4 (S) | 2/4 (S) | 2/4 (S) | |||||
| cephems | cefotaxime | 0.12 (S) | 0.12 (S) | 0.12 (S) | >8 (R) | blaDHA-1, blaSHV-12 | |||
| ceftazidime | 0.5 (S) | 0.25 (S) | 0.5 (S) | >16 (R) | |||||
| cefepime | ≤0.12 (S) | ≤0.12 (S) | ≤0.12 (S) | 2 (S) | |||||
| monobactams | aztreonam | ≤0.12 (S) | ≤0.12 (S) | ≤0.12 (S) | >16 (R) | blaDHA-1, blaSHV-12 | |||
| carbapenems | meropenem | ≤0.12 (S) | ≤0.12 (S) | ≤0.12 (S) | ≤0.12 (S) | ||||
| ertapenem | ≤0.015 (S) | ≤0.015 (S) | ≤0.015 (S) | 0.03 (S) | |||||
| aminoglycosides | gentamicin | 0.5 (S) | 0.5 (S) | aph(3’’)-Ib, aph(6)-Id, aadA1 | 0.5 (S) | >32 (R) | aac(3)-II, aac(6’)-IIc, aph(6)-Id, aph(3’’)-Ib | ||
| amikacin | ≤1 (S) | 2 (S) | ≤1 (S) | 2(S) | |||||
| tobramycin | 1 (S) | 1 (S) | 0.5 (S) | >8 (R) | |||||
| trimethoprim-sulfonamide | trimethoprim-sulfamethoxazole | 0.06/1.19 (S) | >4/76 (R) | sul1, sul2, dfrA1 | 0.06/1.19 (S) | >4/76 (R) | sul1, dfrA19 | ||
| quinolones | ciprofloxacin | ≤0.06 (S) | ≤0.06 (S) | ≤0.06 (S) | 0.25 (R) | qnrB4 | |||
| amphenicols | chloramphenicol | 8 (S) | 4 (S) | >32 (R) | catA1 | 4 (S) | |||
| tetracyclines | tetracycline | 2 (S) | >32 (R) | tet(A) | >32 (R) | tet(A) | >32 (R) | tet(B) | |
| Serotype, Strain Name, NCBI GenBank acc. | Replicon Name | Size, bp | AMR Genes 1 | Replion Type(s) 2 | Mobility Prediction for the Plasmid 3 |
|---|---|---|---|---|---|
| S. Enteritidis, SLR1_8250, CP060522-CP060525 | chromosome | 4,679,617 | ND | chromosome | ND |
| pS8250-1 | 59,372 | ND | IncFII(S), IncFIB(S) | non-mobilizable | |
| pS8250-2 | 33,310 | mcr-1.1 | IncX4 | conjugative | |
| pS8250-3 | 2096 | ND | ColpVC | mobilizable | |
| S. Enteritidis, SLR1_8245 | chromosome | 4,680,323 | ND | chromosome | - |
| pS8245-1 | 59,372 | ND | IncFII(S), IncFIB(S) | non-mobilizable | |
| pS8245-2 | 56,813 | catA1, tet(A) | IncX1 | conjugative | |
| pS8245-3 | 33,310 | mcr-1.1 | IncX4 | conjugative | |
| S. Bovismorbificans, SLR1_7627, CP060517-CP060521 | chromosome | 4,715,485 | ND | chromosome | ND |
| pS7627-1 | 233,305 | aph(3’’)-Ib, aph(6)-Id, aadA1, dfrA1, tet(A), blaTEM-1B, sul1, sul2 | IncHI2, IncQ1 | non-mobilizable | |
| pS7627-2 | 64,443 | mcr-1.1 | IncI2 | conjugative | |
| pS7627-3 | 4073 | ND | rep_cluster_2350 3 | mobilizable | |
| pS7627-4 | 3830 | ND | rep_cluster_2335 3 | mobilizable | |
| S. Typhimurium [4,5:i:-], SLR1_8094, CP060515-CP060516 | chromosome | 5,017,156 | tet(B) | chromosome | ND |
| pS8094-1 | 278,034 | aph(3’’)-Ib, aph(6)-Id, aac(3)-IIg, aac(6’)-IIc, ere(A), qnrB4, dfrA19,mcr-9, blaSHV-12, blaDHA-1, sul1 x2 | IncHI2 | conjugative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuleshov, K.V.; Pavlova, A.S.; Shedko, E.D.; Mikhaylova, Y.V.; Margos, G.; Hepner, S.; Chebotar, I.V.; Korneenko, E.V.; Podkolzin, A.T.; Akimkin, V.G. Mobile Colistin Resistance Genetic Determinants of Non-Typhoid Salmonella enterica Isolates from Russia. Microorganisms 2021, 9, 2515. https://doi.org/10.3390/microorganisms9122515
Kuleshov KV, Pavlova AS, Shedko ED, Mikhaylova YV, Margos G, Hepner S, Chebotar IV, Korneenko EV, Podkolzin AT, Akimkin VG. Mobile Colistin Resistance Genetic Determinants of Non-Typhoid Salmonella enterica Isolates from Russia. Microorganisms. 2021; 9(12):2515. https://doi.org/10.3390/microorganisms9122515
Chicago/Turabian StyleKuleshov, Konstantin V., Anastasia S. Pavlova, Elizaveta D. Shedko, Yulia V. Mikhaylova, Gabriele Margos, Sabrina Hepner, Igor V. Chebotar, Elena V. Korneenko, Alexander T. Podkolzin, and Vasiliy G. Akimkin. 2021. "Mobile Colistin Resistance Genetic Determinants of Non-Typhoid Salmonella enterica Isolates from Russia" Microorganisms 9, no. 12: 2515. https://doi.org/10.3390/microorganisms9122515
APA StyleKuleshov, K. V., Pavlova, A. S., Shedko, E. D., Mikhaylova, Y. V., Margos, G., Hepner, S., Chebotar, I. V., Korneenko, E. V., Podkolzin, A. T., & Akimkin, V. G. (2021). Mobile Colistin Resistance Genetic Determinants of Non-Typhoid Salmonella enterica Isolates from Russia. Microorganisms, 9(12), 2515. https://doi.org/10.3390/microorganisms9122515