
Genomic and Phenotypic Characteristics in Geographically Separated Clinical Campylobacter jejuni ST353CC Isolates


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data on Participants and Microbiota Analyses
2.2. Bacterial Culture Conditions
2.3. Whole-Genome Sequencing
2.4. Genomics
2.5. RNA Preparation from Bacteria and cDNA Synthesis
2.6. Quantitative PCR
2.7. Cell Culture Conditions
2.8. In Vitro Cell Adhesion Assay and IL-8 ELISA
3. Results
3.1. Phylogenetic Characterization of C. jejuni Isolates
3.2. Genomic Characterization of C. jejuni ST353CC Isolates
3.3. Various Symptom Scores in Travelers Infected with C. jejuni ST353CC Isolates
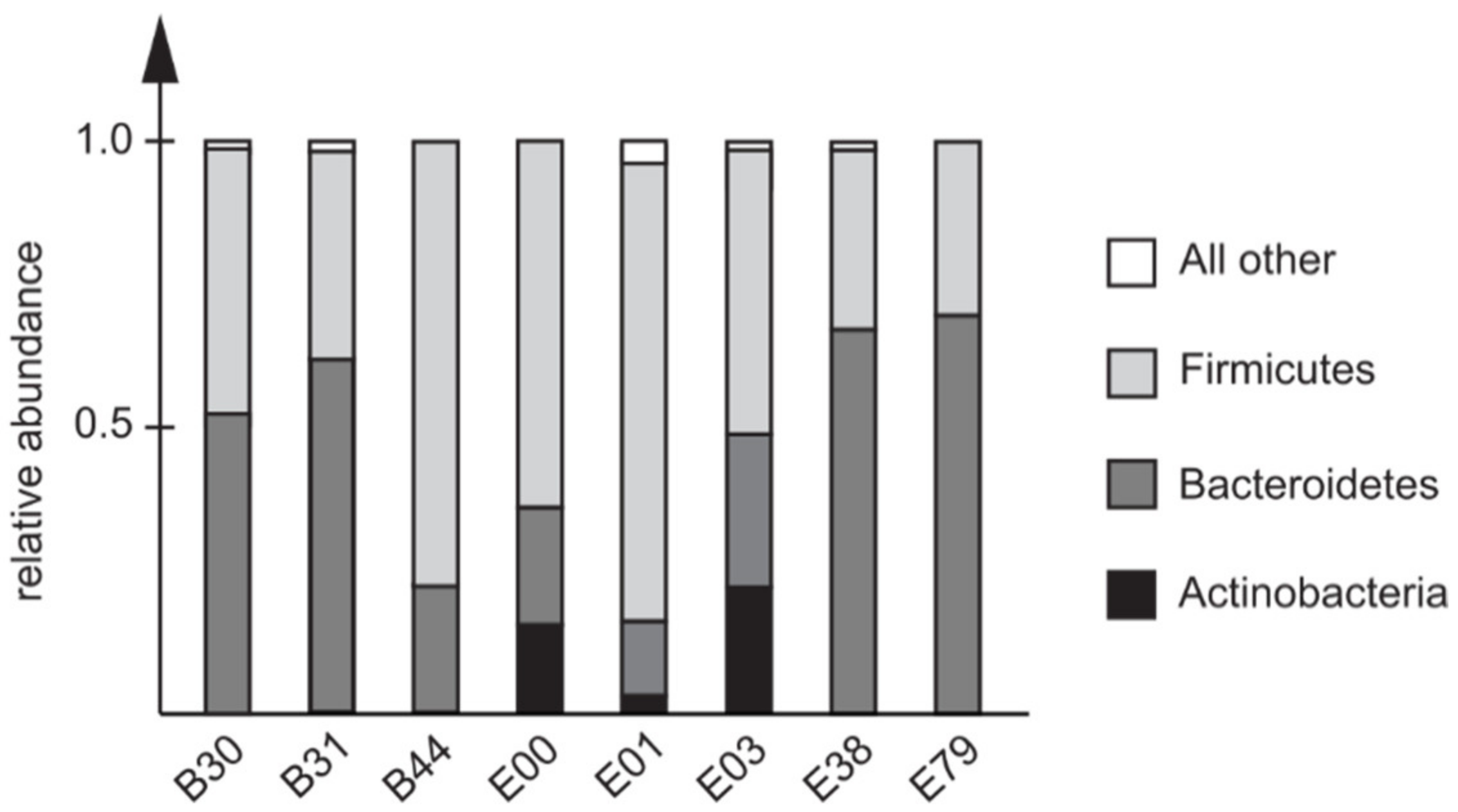
3.4. Pre-Travel Composition of the Fecal Microbiota in Travelers Infected with C. jejuni ST353CC Isolates
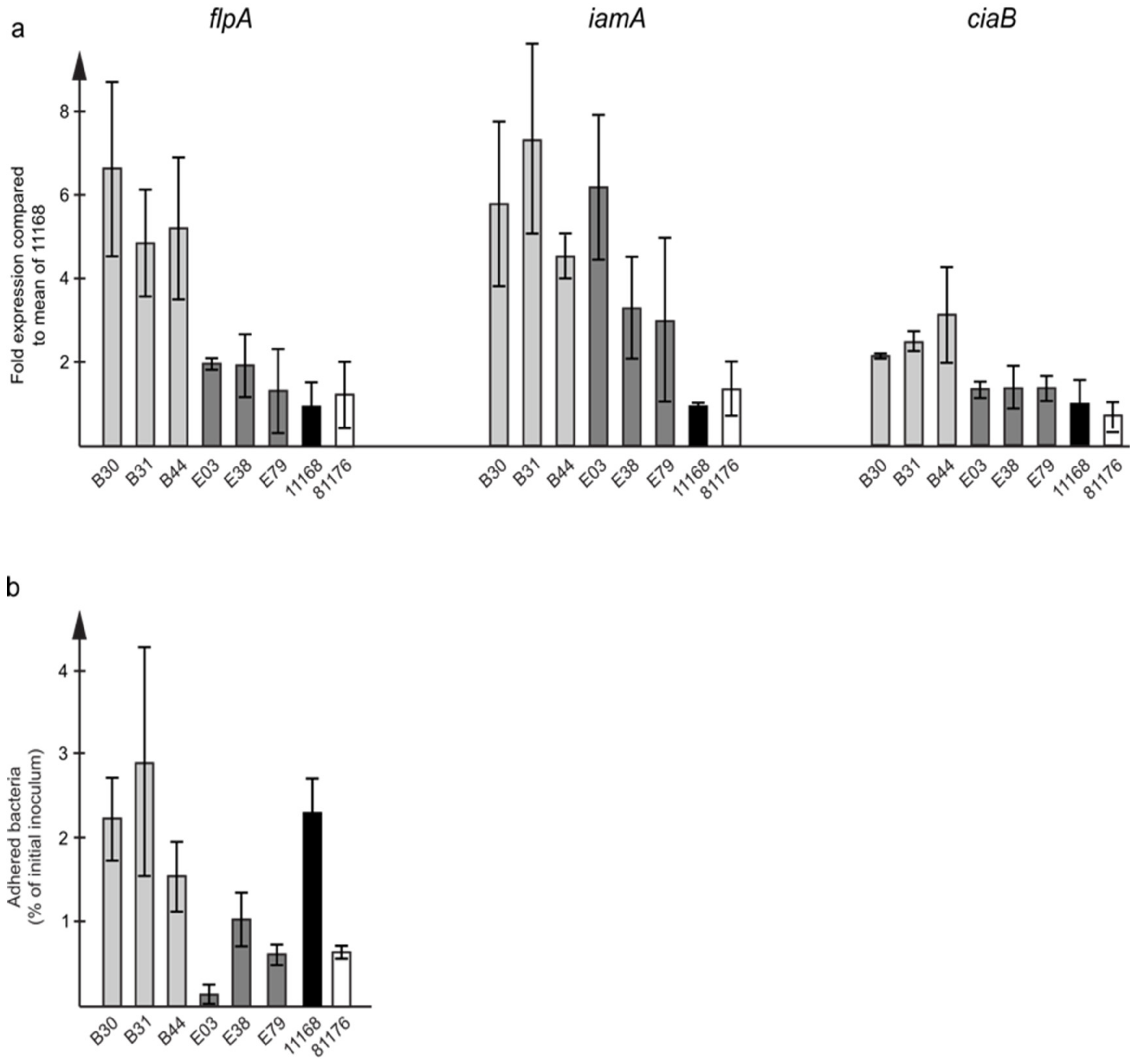
3.5. In Vitro Characteristics of C. jejuni ST353CC Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA; ECDC. The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, 5926. [Google Scholar]
- Humphrey, T.; O’Brien, S.; Madsen, M. Campylobacters as zoonotic pathogens: A food production perspective. Int. J. Food Microbiol. 2007, 117, 237–257. [Google Scholar] [CrossRef] [PubMed]
- Blaser, M.J. Epidemiologic and clinical features of Campylobacter jejuni infections. J. Infect. Dis. 1997, 176 (Suppl. 2), S103–S105. [Google Scholar] [CrossRef] [Green Version]
- Hannu, T.; Mattila, L.; Rautelin, H.; Pelkonen, P.; Lahdenne, P.; Siitonen, A.; Leirisalo-Repo, M. Campylobacter-triggered reactive arthritis: A population-based study. Rheumatology 2002, 41, 312–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, L.A.; Ruigomez, A. Increased risk of irritable bowel syndrome after bacterial gastroenteritis: Cohort study. BMJ 1999, 318, 565–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, N.; Giesecke, J. Incidence of Guillain-Barre syndrome following infection with Campylobacter jejuni. Am. J. Epidemiol. 2001, 153, 610–614. [Google Scholar] [CrossRef]
- PubMLST. Sequence Query—Campylobacter Locus/Sequence Definitions. 2021. Available online: https://pubmlst.org/bigsdb?db=pubmlst_campylobacter_seqdef&page=sequenceQuery (accessed on 5 September 2021).
- Gripp, E.; Hlahla, D.; Didelot, X.; Kops, F.; Maurischat, S.; Tedin, K.; Alter, T.; Ellerbroek, L.; Schreiber, K.; Schomburg, D.; et al. Closely related Campylobacter jejuni strains from different sources reveal a generalist rather than a specialist lifestyle. BMC Genom. 2011, 12, 584. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, S.K.; Dallas, J.F.; MacRae, M.; McCarthy, N.D.; Sproston, E.L.; Gormley, F.J.; Strachan, N.J.; Ogden, I.D.; Maiden, M.C.; Forbes, K.J. Campylobacter genotypes from food animals, environmental sources and clinical disease in Scotland 2005/6. Int. J. Food Microbiol. 2009, 134, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Epping, L.; Antão, E.M.; Semmler, T. Population biology and comparative genomics of Campylobacter species. In Fighting Campylobacter Infections. Current Topics in Microbiology and Immunology; Backert, S., Ed.; Springer: Cham, Switzerland, 2021; p. 431. [Google Scholar]
- Flanagan, R.C.; Neal-McKinney, J.M.; Dhillon, A.S.; Miller, W.G.; Konkel, M.E. Examination of Campylobacter jejuni putative adhesins leads to the identification of a new protein, designated FlpA, required for chicken colonization. Infect. Immun. 2009, 77, 2399–2407. [Google Scholar] [CrossRef] [Green Version]
- Konkel, M.E.; Garvis, S.G.; Tipton, S.L.; Anderson, D.E., Jr.; Cieplak, W., Jr. Identification and molecular cloning of a gene encoding a fibronectin-binding protein (CadF) from Campylobacter jejuni. Mol. Microbiol. 1997, 24, 953–963. [Google Scholar] [CrossRef]
- Konkel, M.E.; Larson, C.L.; Flanagan, R.C. Campylobacter jejuni FlpA binds fibronectin and is required for maximal host cell adherence. J. Bacteriol. 2010, 192, 68–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziprin, R.L.; Young, C.R.; Stanker, L.H.; Hume, M.E.; Konkel, M.E. The absence of cecal colonization of chicks by a mutant of Campylobacter jejuni not expressing bacterial fibronectin-binding protein. Avian Dis. 1999, 43, 586–589. [Google Scholar] [CrossRef]
- Carvalho, A.C.; Ruiz-Palacios, G.M.; Ramos-Cervantes, P.; Cervantes, L.E.; Jiang, X.; Pickering, L.K. Molecular characterization of invasive and noninvasive Campylobacter jejuni and Campylobacter coli isolates. J. Clin. Microbiol. 2001, 39, 1353–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konkel, M.E.; Kim, B.J.; Rivera-Amill, V.; Garvis, S.G. Identification of proteins required for the internalization of Campylobacter jejuni into cultured mammalian cells. Adv. Exp. Med. Biol. 1999, 473, 215–224. [Google Scholar]
- Ziprin, R.L.; Young, C.R.; Byrd, J.A.; Stanker, L.H.; Hume, M.E.; Gray, S.A.; Kim, B.J.; Konkel, M.E. Role of Campylobacter jejuni potential virulence genes in cecal colonization. Avian Dis. 2001, 45, 549–557. [Google Scholar] [CrossRef]
- Johansson, C.; Nilsson, A.; Kaden, R.; Rautelin, H. Campylobacter coli clade 3 isolates induce rapid cell death in vitro. Appl. Environ. Microbiol. 2019, 85, e02993-18. [Google Scholar] [CrossRef] [Green Version]
- Johansson, C.; Nilsson, A.; Kaden, R.; Rautelin, H. Differences in virulence gene expression between human blood and stool Campylobacter coli clade 1 ST828CC isolates. Gut Pathog. 2019, 11, 42. [Google Scholar] [CrossRef]
- Nilsson, A.; Johansson, C.; Skarp, A.; Kaden, R.; Engstrand, L.; Rautelin, H. Genomic and phenotypic characteristics of Swedish C. jejuni water isolates. PLoS ONE 2017, 12, e0189222. [Google Scholar] [CrossRef] [Green Version]
- Skarp, C.P.A.; Akinrinade, O.; Kaden, R.; Johansson, C.; Rautelin, H. Accessory genetic content in Campylobacter jejuni ST21CC isolates from feces and blood. Int. J. Med. Microbiol. 2017, 307, 233–240. [Google Scholar] [CrossRef]
- Nilsson, A.; Skarp, A.; Johansson, C.; Kaden, R.; Engstrand, L.; Rautelin, H. Characterization of Swedish Campylobacter coli clade 2 and clade 3 water isolates. Microbiologyopen 2018, 7, e00583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dicksved, J.; Ellström, P.; Engstrand, L.; Rautelin, H. Susceptibility to Campylobacter infection is associated with the species composition of the human fecal microbiota. mBio 2014, 5, e01212-14. [Google Scholar] [CrossRef] [Green Version]
- Kampmann, C.; Dicksved, J.; Engstrand, L.; Rautelin, H. Composition of human faecal microbiota in resistance to Campylobacter infection. Clin. Microbiol. Infect. 2016, 22, 61.e1–61.e8. [Google Scholar] [CrossRef] [Green Version]
- Riddle, M.S.; DuPont, H.L.; Connor, B.A. ACG Clinical Guideline: Diagnosis, treatment, and prevention of acute diarrheal infections in adults. Am. J. Gastroenterol. 2016, 111, 602–622. [Google Scholar] [CrossRef]
- Riddle, M.S.; Connor, B.A.; Beeching, N.J.; DuPont, H.L.; Hamer, D.H.; Kozarsky, P.; Libman, M.; Steffen, R.; Taylor, D.; Tribble, D.R.; et al. Guidelines for the prevention and treatment of travelers’ diarrhea: A graded expert panel report. J. Travel Med. 2017, 24 (Suppl. 1), S63–S80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Center for Biotechnology Information. Genbank. 2019. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 19 June 2019).
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Ågren, J.; Sundstrom, A.; Hafstrom, T.; Segerman, B. Gegenees: Fragmented alignment of multiple genomes for determining phylogenomic distances and genetic signatures unique for specified target groups. PLoS ONE 2012, 7, e39107. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [Green Version]
- National Center for Biotechnology Information. COG List—Camjej 2019. Available online: https://ftp.ncbi.nih.gov/pub/COG/COG2014/static/lists/listCamjej.html (accessed on 23 August 2019).
- Islam, Z.; van Belkum, A.; Wagenaar, J.A.; Cody, A.J.; de Boer, A.G.; Sarker, S.K.; Jacobs, B.C.; Talukder, K.A.; Endtz, H.P. Comparative population structure analysis of Campylobacter jejuni from human and poultry origin in Bangladesh. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 2173–2181. [Google Scholar] [CrossRef] [Green Version]
- Vasco, K.; Graham, J.P.; Trueba, G. Detection of zoonotic enteropathogens in children and domestic animals in a semirural community in Ecuador. Appl. Environ. Microbiol. 2016, 82, 4218–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinueza-Burgos, C.; Wautier, M.; Martiny, D.; Cisneros, M.; Van Damme, I.; De Zutter, L. Prevalence, antimicrobial resistance and genetic diversity of Campylobacter coli and Campylobacter jejuni in Ecuadorian broilers at slaughter age. Poult. Sci. 2017, 96, 2366–2374. [Google Scholar] [CrossRef]
- Konkel, M.E.; Christensen, J.E.; Keech, A.M.; Monteville, M.R.; Klena, J.D.; Garvis, S.G. Identification of a fibronectin-binding domain within the Campylobacter jejuni CadF protein. Mol. Microbiol. 2005, 57, 1022–1035. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Fischer, A.; Plickert, R.; Haag, L.M.; Otto, B.; Kühl, A.A.; Dashti, J.I.; Zautner, A.E.; Muñoz, M.; Loddenkemper, C.; et al. Novel murine infection models provide deep insights into the “menage a trois” of Campylobacter jejuni, microbiota and host innate immunity. PLoS ONE 2011, 6, e20953. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef]
- Gentile, C.R.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [Green Version]
- Losasso, C.; Eckert, E.M.; Mastrorilli, E.; Villiger, J.; Mancin, M.; Patuzzi, I.; Di Cesare, A.; Cibin, V.; Barrucci, F.; Pernthaler, J.; et al. Assessing the influence of vegan, vegetarian and omnivore oriented westernized dietary styles on human gut microbiota: A cross sectional study. Front. Microbiol. 2018, 9, 317. [Google Scholar] [CrossRef] [PubMed]
- Franco-de-Moraes, A.C.; de Almeida-Pititto, B.; da Rocha Fernandes, G.; Gomes, E.P.; da Costa Pereira, A.; Ferreira, S.R.G. Worse inflammatory profile in omnivores than in vegetarians associates with the gut microbiota composition. Diabetol. Metab. Syndr. 2017, 9, 62. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Travel Destination | Gender | Age (Years) | Isolate | CC/ST # | Accession |
|---|---|---|---|---|---|
| Bangladesh | F | 24 | B29 | 354/354 | JAGFPS000000000 |
| F | 24 | B30 | 353/9438 ^ | JAGFPT000000000 | |
| F | 28 | B31 | 353/9438 ^ | JAGFPR000000000 | |
| F | 26 | B44 | 353/9438 ^ | JAGFPU000000000 | |
| Ecuador | F | 29 | E00 * | 353/462 | JAGFPV000000000 |
| M | 24 | E01 * | 353/9336 | JAGFPN000000000 | |
| F | 20 | E03 | 353/9336 | JAGFPO000000000 | |
| F | 20 | E38 | 353/9437 ^ | JAGFPP000000000 | |
| F | 23 | E61 | 607/607 | JAGFPQ000000000 | |
| F | 19 | E63 | 607/607 | JAGFPL000000000 | |
| F | 22 | E79 | 353/3515 | JAGFPM000000000 |
| Score | Symptom | Description |
|---|---|---|
| 1 | No symptoms | |
| 2 | Mild symptoms | Slight abdominal discomfort, ≤3 loose stools in 24 h, symptoms did not force any change in planned daily activities |
| 3 | Severe diarrhea | >3 loose stools in 24 h + enteric symptoms (e.g., fecal urgency, nausea, abdominal pain/cramps, emesis), symptoms did not force a significant change in planned daily activities, duration ≤3 consecutive days |
| 4 | Severe diarrhea | >3 loose stools in 24 h + enteric symptoms for >3 consecutive days, symptoms forced a significant change in planned daily activities and/or affected general health |
| 5 | Severe diarrhea | Bloody diarrhea, and/or fever, and/or severely affected general health |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johansson, C.; Kampmann, C.; Nilsson, A.; Dicksved, J.; Engstrand, L.; Rautelin, H. Genomic and Phenotypic Characteristics in Geographically Separated Clinical Campylobacter jejuni ST353CC Isolates. Microorganisms 2021, 9, 2540. https://doi.org/10.3390/microorganisms9122540
Johansson C, Kampmann C, Nilsson A, Dicksved J, Engstrand L, Rautelin H. Genomic and Phenotypic Characteristics in Geographically Separated Clinical Campylobacter jejuni ST353CC Isolates. Microorganisms. 2021; 9(12):2540. https://doi.org/10.3390/microorganisms9122540
Chicago/Turabian StyleJohansson, Cecilia, Christian Kampmann, Anna Nilsson, Johan Dicksved, Lars Engstrand, and Hilpi Rautelin. 2021. "Genomic and Phenotypic Characteristics in Geographically Separated Clinical Campylobacter jejuni ST353CC Isolates" Microorganisms 9, no. 12: 2540. https://doi.org/10.3390/microorganisms9122540
APA StyleJohansson, C., Kampmann, C., Nilsson, A., Dicksved, J., Engstrand, L., & Rautelin, H. (2021). Genomic and Phenotypic Characteristics in Geographically Separated Clinical Campylobacter jejuni ST353CC Isolates. Microorganisms, 9(12), 2540. https://doi.org/10.3390/microorganisms9122540