
Ovariectomy-Induced Dysbiosis May Have a Minor Effect on Bone in Mice

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Ovariectomized Mice Model
2.3. Antibiotic Treatment
2.4. Transplantation of Cecal Luminal Contents
2.5. Micro-Computed Tomography Measurements
2.6. Serum Biomarkers of Bone Metabolism
2.7. Serum Lipopolysaccharide (LPS) Measurement
2.8. 16S rRNA Gene Sequencing for Microbial Analysis
2.9. RNA Isolation and Quantitative Reverse Transcription (qRT-)PCR
2.10. Statistical Analyses
3. Results
3.1. Ovariectomy Decreased Bone Mineral Density
3.2. Ovariectomy Increased the Level of Tumor Necrosis Factor Alpha mRNA and Altered Barrier Function in the Colon
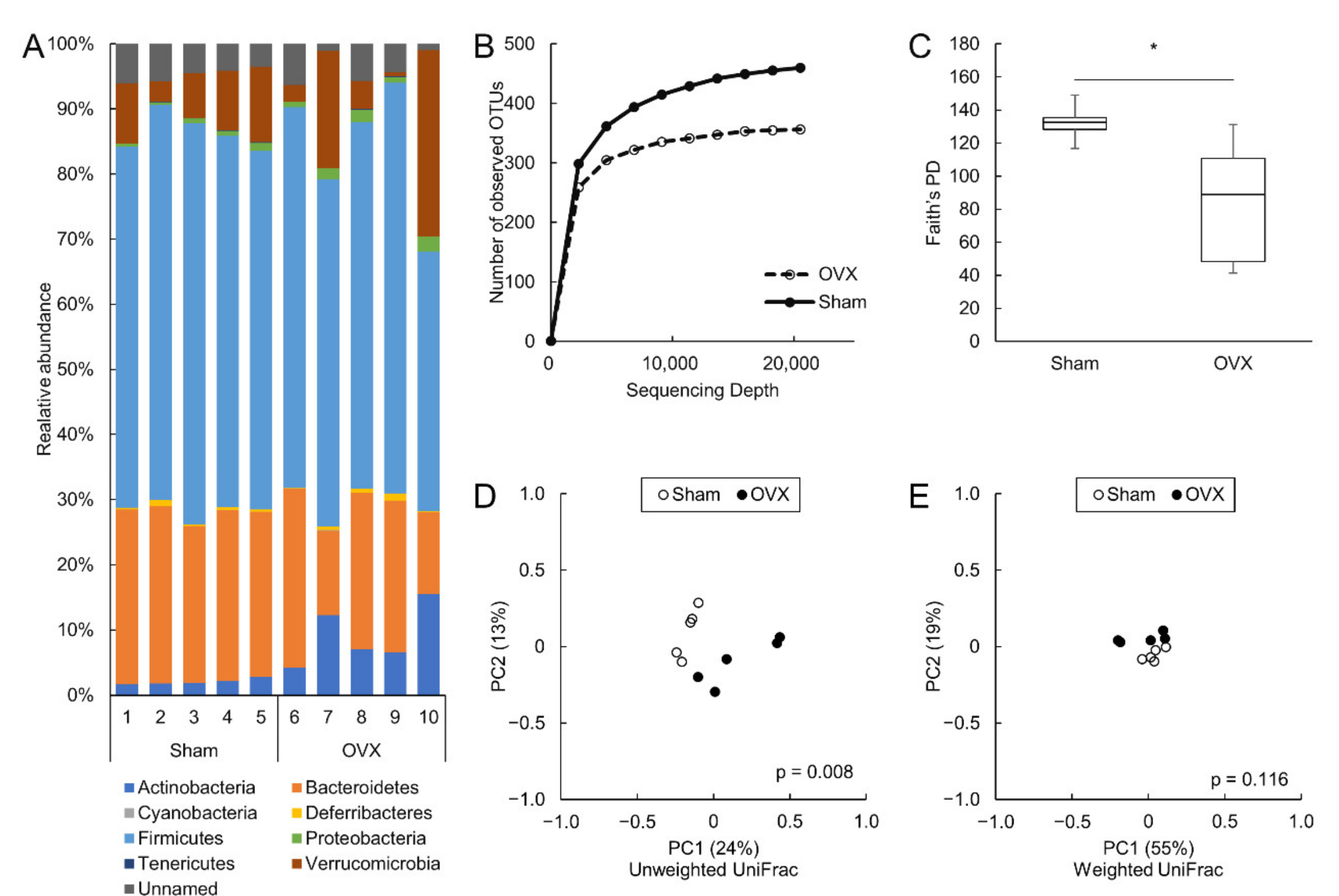
3.3. Ovariectomy Decreased Alpha Diversity in the Colon and Altered the Colonic Microbiota
3.4. Antibiotics Decreased Bone Formation but Did Not Affect Bone Mineral Density
3.5. Fecal Microbiota Transplantation from OVX Mice to Antibiotic-Treated Non-OVX- or OVX Mice Did Not Change Bone Homeostasis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eastell, R.; O’Neill, T.W.; Hofbauer, L.C.; Langdahl, B.; Reid, I.R.; Gold, D.T.; Cummings, S.R. Postmenopausal Osteoporosis. Nat. Rev. Dis. Primers 2016, 2, 16069. [Google Scholar] [CrossRef] [PubMed]
- Khosla, S.; Melton, L.J.; Riggs, B.L. The Unitary Model for Estrogen Deficiency and the Pathogenesis of Osteoporosis: Is a Revision Needed? J. Bone Miner. Res. 2011, 26, 441–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briot, K.; Geusens, P.; Em Bultink, I.; Lems, W.F.; Roux, C. Inflammatory Diseases and Bone Fragility. Osteoporos Int. 2017, 28, 3301–3314. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Hara, E. Relationship between Obesity, Gut Microbiome and Hepatocellular Carcinoma Development. Dig. Dis. 2015, 33, 346–350. [Google Scholar] [CrossRef]
- Jiang, C.; Li, G.; Huang, P.; Liu, Z.; Zhao, B. The Gut Microbiota and Alzheimer’s Disease. J Alzheimers Dis. 2017, 58, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cosola, C.; Rocchetti, M.T.; Sabatino, A.; Fiaccadori, E.; Di Iorio, B.R.; Gesualdo, L. Microbiota Issue in CKD: How Promising Are Gut-Targeted Approaches? J. Nephrol. 2019, 32, 27–37. [Google Scholar] [CrossRef]
- Nadatani, Y.; Watanabe, T.; Suda, W.; Nakata, A.; Matsumoto, Y.; Kosaka, S.; Higashimori, A.; Otani, K.; Hosomi, S.; Tanaka, F.; et al. Gastric Acid Inhibitor Aggravates Indomethacin-Induced Small Intestinal Injury via Reducing Lactobacillus Johnsonii. Sci. Rep. 2019, 9, 17490. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Higuchi, K.; Kobata, A.; Nishio, H.; Tanigawa, T.; Shiba, M.; Tominaga, K.; Fujiwara, Y.; Oshitani, N.; Asahara, T.; et al. Non-Steroidal Anti-Inflammatory Drug-Induced Small Intestinal Damage Is Toll-like Receptor 4 Dependent. Gut 2008, 57, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Nadatani, Y.; Suda, W.; Higashimori, A.; Otani, K.; Fukunaga, S.; Hosomi, S.; Tanaka, F.; Nagami, Y.; Taira, K.; et al. Long-Term Persistence of Gastric Dysbiosis after Eradication of Helicobacter Pylori in Patients Who Underwent Endoscopic Submucosal Dissection for Early Gastric Cancer. Gastric Cancer 2021, 24, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-Y.; Chassaing, B.; Tyagi, A.M.; Vaccaro, C.; Luo, T.; Adams, J.; Darby, T.M.; Weitzmann, M.N.; Mulle, J.G.; Gewirtz, A.T.; et al. Sex Steroid Deficiency-Associated Bone Loss Is Microbiota Dependent and Prevented by Probiotics. J. Clin. Investig. 2016, 126, 2049–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Herzog, J.W.; Tsang, K.; Brennan, C.A.; Bower, M.A.; Garrett, W.S.; Sartor, B.R.; Aliprantis, A.O.; Charles, J.F. Gut Microbiota Induce IGF-1 and Promote Bone Formation and Growth. Proc. Natl. Acad. Sci. USA 2016, 113, E7554–E7563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, M.; Cronin, O.; Keohane, D.M.; Cormac, E.M.; Nugent, H.; Nugent, M.; Molloy, C.; O’Toole, P.W.; Shanahan, F.; Molloy, M.G.; et al. Gut Microbiota Alterations Associated with Reduced Bone Mineral Density in Older Adults. Rheumatology 2019, 58, 2295–2304. [Google Scholar] [CrossRef] [Green Version]
- McCabe, L.R.; Irwin, R.; Schaefer, L.; Britton, R.A. Probiotic Use Decreases Intestinal Inflammation and Increases Bone Density in Healthy Male but Not Female Mice. J. Cell. Physiol. 2013, 228, 1793–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, R.A.; Irwin, R.; Quach, D.; Schaefer, L.; Zhang, J.; Lee, T.; Parameswaran, N.; McCabe, L.R.; Probiotic, L. Reuteri Treatment Prevents Bone Loss in a Menopausal Ovariectomized Mouse Model. J. Cell. Physiol. 2014, 229, 1822–1830. [Google Scholar] [CrossRef] [Green Version]
- Quach, D.; Collins, F.; Parameswaran, N.; McCabe, L.; Britton, R.A. Microbiota Reconstitution Does Not Cause Bone Loss in Germ-Free Mice. mSphere 2018, 3, e00545-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjögren, K.; Engdahl, C.; Henning, P.; Lerner, U.H.; Tremaroli, V.; Lagerquist, M.K.; Bäckhed, F.; Ohlsson, C. Ohlsson Claes The Gut Microbiota Regulates Bone Mass in Mice. J. Bone Miner. Res. 2012, 27, 1357–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvaneh, K.; Ebrahimi, M.; Sabran, M.R.; Karimi, G.; Hwei, A.N.M.; Abdul-Majeed, S.; Ahmad, Z.; Ibrahim, Z.; Jamaluddin, R. Probiotics (Bifidobacterium longum) Increase Bone Mass Density and Upregulate Sparc and Bmp-2 Genes in Rats with Bone Loss Resulting from Ovariectomy. Biomed. Res. Int. 2015, 2015, 897639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohlsson, C.; Engdahl, C.; Fåk, F.; Andersson, A.; Windahl, S.H.; Farman, H.H.; Movérare-Skrtic, S.; Islander, U.; Sjögren, K. Probiotics Protect Mice from Ovariectomy-Induced Cortical Bone Loss. PLoS ONE 2014, 9, e92368. [Google Scholar] [CrossRef]
- Flores, R.; Shi, J.; Fuhrman, B.; Xu, X.; Veenstra, T.D.; Gail, M.H.; Gajer, P.; Ravel, J.; Goedert, J.J. Fecal Microbial Determinants of Fecal and Systemic Estrogens and Estrogen Metabolites: A Cross-Sectional Study. J. Transl. Med. 2012, 10, 253. [Google Scholar] [CrossRef] [Green Version]
- Sophocleous, A.; Idris, A.I. Rodent Models of Osteoporosis. Bonekey Rep. 2014, 3, 614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eri, T.; Kawahata, K.; Kanzaki, T.; Imamura, M.; Michishita, K.; Akahira, L.; Bannai, E.; Yoshikawa, N.; Kimura, Y.; Satoh, T.; et al. Intestinal Microbiota Link Lymphopenia to Murine Autoimmunity via PD-1+CXCR5-/Dim B-Helper T Cell Induction. Sci. Rep. 2017, 7, 46037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usui, Y.; Kimura, Y.; Satoh, T.; Takemura, N.; Ouchi, Y.; Ohmiya, H.; Kobayashi, K.; Suzuki, H.; Koyama, S.; Hagiwara, S.; et al. Effects of Long-Term Intake of a Yogurt Fermented with Lactobacillus Delbrueckii Subsp. Bulgaricus 2038 and Streptococcus Thermophilus 1131 on Mice. Int. Immunol. 2018, 30, 319–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, K.; Kawaguchi, Y.; Shimohigoshi, M.; Gotoh, Y.; Nakano, Y.; Usui, Y.; Hayashi, T.; Kimura, Y.; Uematsu, M.; Yamamoto, T.; et al. Antigen-Specific Mucosal Immunity Regulates Development of Intestinal Bacteria-Mediated Diseases. Gastroenterology 2019, 157, 1530–1543. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimura, N.; Otani, K.; Watanabe, T.; Nakatsu, G.; Shimada, S.; Fujimoto, K.; Nadatani, Y.; Hosomi, S.; Tanaka, F.; Kamata, N.; et al. High-Fat Diet-Mediated Dysbiosis Exacerbates NSAID-Induced Small Intestinal Damage through the Induction of Interleukin-17A. Sci. Rep. 2019, 9, 16796. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Hwang, Y.-J.; Shin, M.-J.; Yi, H. Difference in the Gut Microbiome between Ovariectomy-Induced Obesity and Diet-Induced Obesity. J. Microbiol. Biotechnol. 2017, 27, 2228–2236. [Google Scholar] [CrossRef] [PubMed]
- Cox-York, K.A.; Sheflin, A.M.; Foster, M.T.; Gentile, C.L.; Kahl, A.; Koch, L.G.; Britton, S.L.; Weir, T.L. Ovariectomy Results in Differential Shifts in Gut Microbiota in Low versus High Aerobic Capacity Rats. Physiol. Rep. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Pacifici, R. Bone Remodeling and the Microbiome. Cold Spring Harb. Perspect. Med. 2018, 8, a031203. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The Gut Microbiota Shapes Intestinal Immune Responses during Health and Disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Rakoff-Nahoum, S.; Paglino, J.; Eslami-Varzaneh, F.; Edberg, S.; Medzhitov, R. Recognition of Commensal Microflora by Toll-Like Receptors Is Required for Intestinal Homeostasis. Cell 2004, 118, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Rey, K.; Manku, S.; Enns, W.; Van Rossum, T.; Bushell, K.; Morin, R.D.; Brinkman, F.S.L.; Choy, J.C. Disruption of the Gut Microbiota With Antibiotics Exacerbates Acute Vascular Rejection. Transplantation 2018, 102, 1085–1095. [Google Scholar] [CrossRef]
- Takashima, S.; Tanaka, F.; Kawaguchi, Y.; Usui, Y.; Fujimoto, K.; Nadatani, Y.; Otani, K.; Hosomi, S.; Nagami, Y.; Kamata, N.; et al. Proton Pump Inhibitors Enhance Intestinal Permeability via Dysbiosis of Gut Microbiota under Stressed Conditions in Mice. Neurogastroenterol. Motil. 2020, 32, e13841. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Yamanishi, S.; Sohn, J.; Alekseyenko, A.V.; Leung, J.M.; Cho, I.; Kim, S.G.; Li, H.; Gao, Z.; Mahana, D.; et al. Altering the Intestinal Microbiota during a Critical Developmental Window Has Lasting Metabolic Consequences. Cell 2014, 158, 705–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guss, J.D.; Horsfield, M.W.; Fontenele, F.F.; Sandoval, T.N.; Luna, M.; Apoorva, F.; Lima, S.F.; Bicalho, R.C.; Singh, A.; Ley, R.E.; et al. Alterations to the Gut Microbiome Impair Bone Strength and Tissue Material Properties. J. Bone Min. Res 2017, 32, 1343–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, F.L.; Rios-Arce, N.D.; Schepper, J.D.; Parameswaran, N.; McCabe, L.R. The Potential of Probiotics as a Therapy for Osteoporosis. Microbiol. Spectr. 2017, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Seely, K.D.; Kotelko, C.A.; Douglas, H.; Bealer, B.; Brooks, A.E. The Human Gut Microbiota: A Key Mediator of Osteoporosis and Osteogenesis. Int. J. Mol. Sci. 2021, 22, 9452. [Google Scholar] [CrossRef] [PubMed]
- Orcel, P.; Feuga, M.; Bielakoff, J.; De Vernejoul, M.C. Local Bone Injections of LPS and M-CSF Increase Bone Resorption by Different Pathways in Vivo in Rats. Am. J. Physiol 1993, 264, E391–E397. [Google Scholar] [CrossRef] [PubMed]
- Miyaura, C.; Inada, M.; Matsumoto, C.; Ohshiba, T.; Uozumi, N.; Shimizu, T.; Ito, A. An Essential Role of Cytosolic Phospholipase A2α in Prostaglandin E2–Mediated Bone Resorption Associated with Inflammation. J. Exp. Med. 2003, 197, 1303–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausmann, E.; Raisz, L.G.; Miller, W.A. Endotoxin: Stimulation of Bone Resorption in Tissue Culture. Science 1970, 168, 862–864. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosaka, S.; Nadatani, Y.; Higashimori, A.; Otani, K.; Fujimoto, K.; Nagata, Y.; Ominami, M.; Fukunaga, S.; Hosomi, S.; Kamata, N.; et al. Ovariectomy-Induced Dysbiosis May Have a Minor Effect on Bone in Mice. Microorganisms 2021, 9, 2563. https://doi.org/10.3390/microorganisms9122563
Kosaka S, Nadatani Y, Higashimori A, Otani K, Fujimoto K, Nagata Y, Ominami M, Fukunaga S, Hosomi S, Kamata N, et al. Ovariectomy-Induced Dysbiosis May Have a Minor Effect on Bone in Mice. Microorganisms. 2021; 9(12):2563. https://doi.org/10.3390/microorganisms9122563
Chicago/Turabian StyleKosaka, Satoshi, Yuji Nadatani, Akira Higashimori, Koji Otani, Kosuke Fujimoto, Yuki Nagata, Masaki Ominami, Shusei Fukunaga, Shuhei Hosomi, Noriko Kamata, and et al. 2021. "Ovariectomy-Induced Dysbiosis May Have a Minor Effect on Bone in Mice" Microorganisms 9, no. 12: 2563. https://doi.org/10.3390/microorganisms9122563