
Pathophysiological Roles of Mucosal-Associated Invariant T Cells in the Context of Gut Microbiota-Liver Axis

 ,
,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
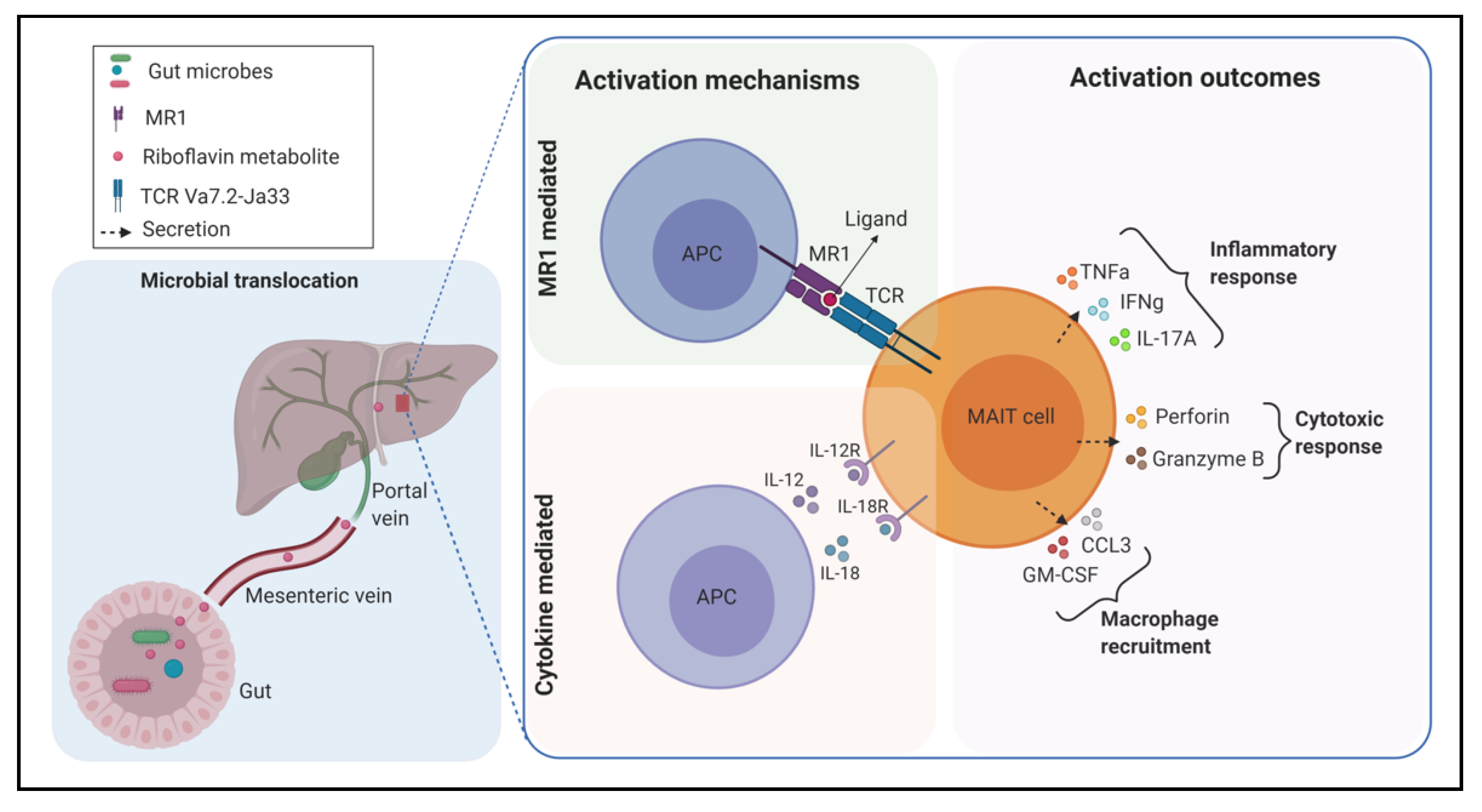
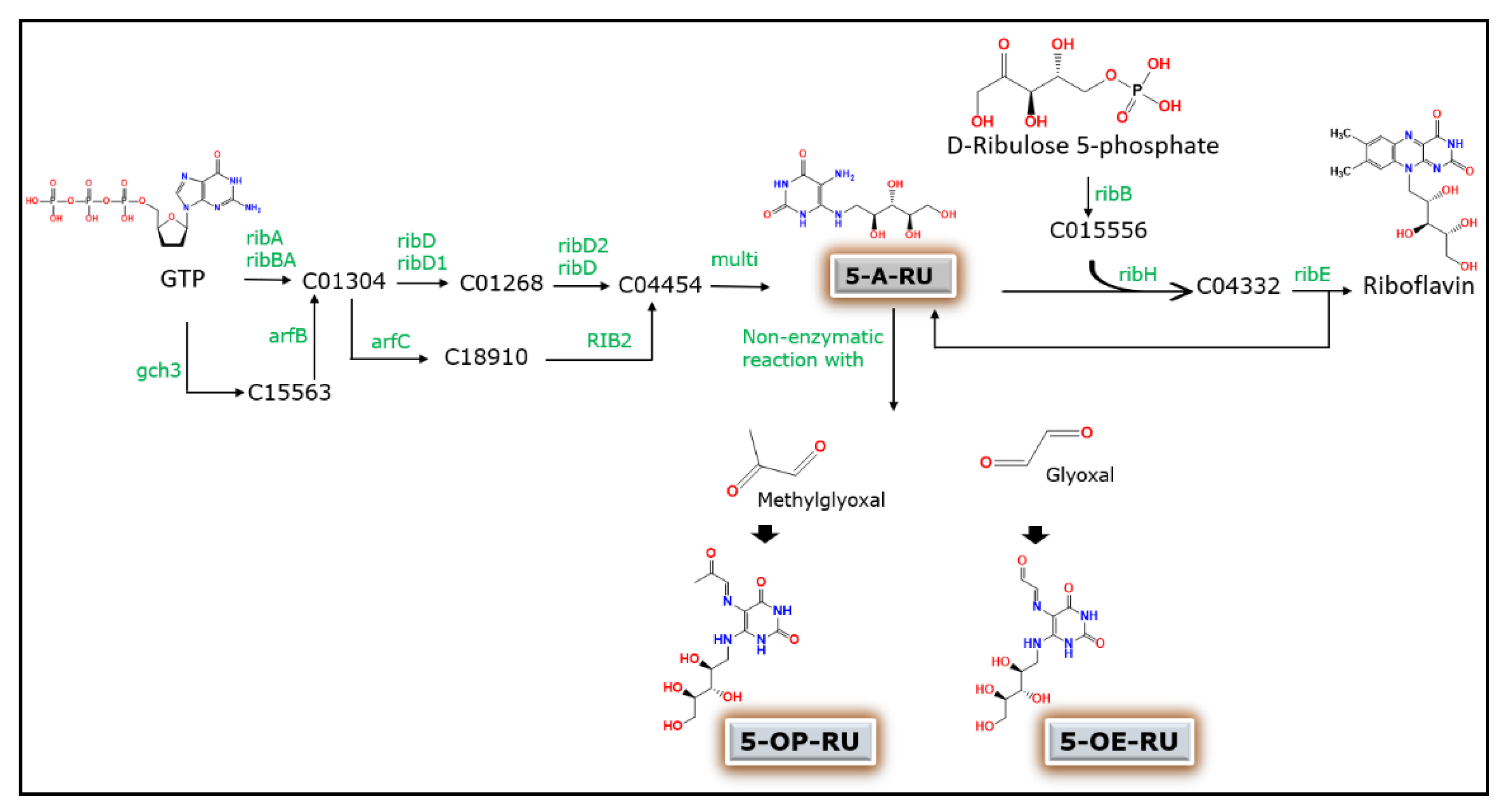
2. Microbial Riboflavin Biosynthesis Pathway and MAIT Cell Activation
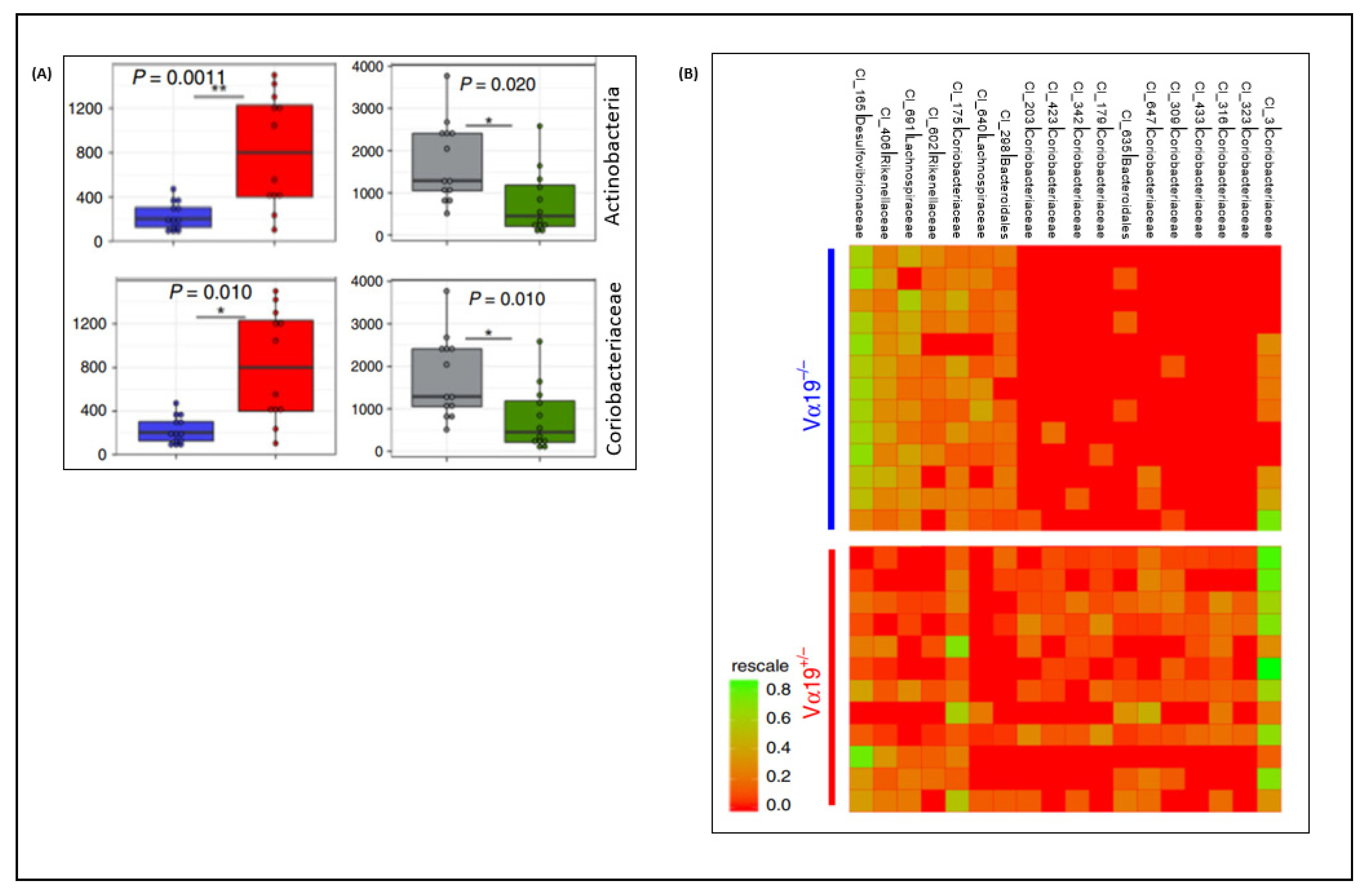
3. Gut Microbiota and MAIT Cells
4. MAIT Cells and Diseases
4.1. MAIT Cells and Liver Disease
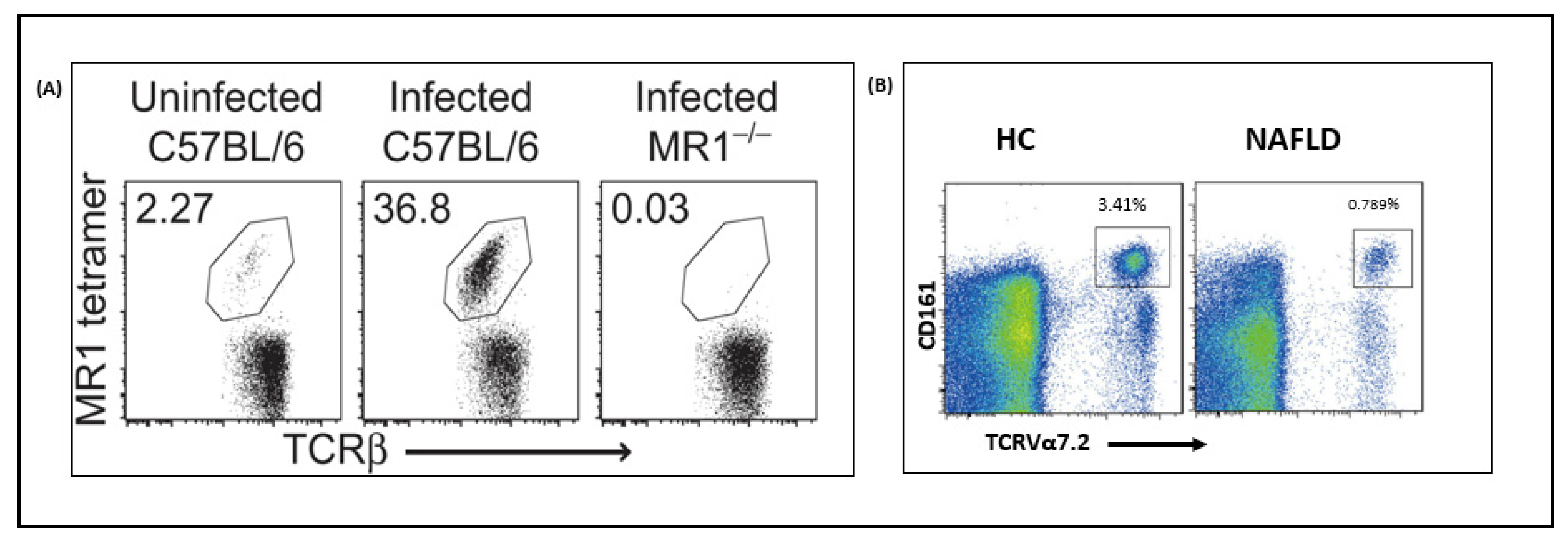
4.1.1. Nonalcoholic Fatty Liver Disease (NAFLD)
4.1.2. Alcoholic Fatty Liver Disease (ALD)
4.1.3. Viral Hepatitis
5. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhati, M.; Cole, D.K.; McCluskey, J.; Sewell, A.K.; Rossjohn, J. The versatility of the alphabeta t-cell antigen receptor. Protein Sci. 2014, 23, 260–272. [Google Scholar] [CrossRef] [Green Version]
- Rossjohn, J.; Gras, S.; Miles, J.J.; Turner, S.J.; Godfrey, D.I.; McCluskey, J. T cell antigen receptor recognition of antigen-presenting molecules. Annu. Rev. Immunol. 2015, 33, 169–200. [Google Scholar] [CrossRef]
- Scaviner, D.; Lefranc, M.P. The human t cell receptor alpha variable (trav) genes. Exp. Clin. Immunogenet. 2000, 17, 83–96. [Google Scholar] [CrossRef] [Green Version]
- Louis, C.U.; Savoldo, B.; Dotti, G.; Pule, M.; Yvon, E.; Myers, G.D.; Rossig, C.; Russell, H.V.; Diouf, O.; Liu, E.; et al. Antitumor activity and long-term fate of chimeric antigen receptor-positive t cells in patients with neuroblastoma. Blood 2011, 118, 6050–6056. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, M.; Ramos, C.A.; Durett, A.; Liu, E.; Dakhova, O.; Liu, H.; Creighton, C.J.; Gee, A.P.; Heslop, H.E.; et al. Closely related t-memory stem cells correlate with in vivo expansion of car Cd19-t cells and are preserved by il-7 and il-15. Blood 2014, 123, 3750–3759. [Google Scholar] [CrossRef] [Green Version]
- Tilloy, F.; Treiner, E.; Park, S.H.; Garcia, C.; Lemonnier, F.; de la Salle, H.; Bendelac, A.; Bonneville, M.; Lantz, O. An invariant t cell receptor alpha chain defines a novel tap-independent major histocompatibility complex class ib-restricted alpha/beta t cell subpopulation in mammals. J. Exp. Med. 1999, 189, 1907–1921. [Google Scholar] [CrossRef]
- Treiner, E.; Duban, L.; Bahram, S.; Radosavljevic, M.; Wanner, V.; Tilloy, F.; Affaticati, P.; Gilfillan, S.; Lantz, O. Selection of evolutionarily conserved mucosal-associated invariant t cells by mr1. Nature 2003, 422, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Porcelli, S.; Yockey, C.E.; Brenner, M.B.; Balk, S.P. Analysis of t cell antigen receptor (tcr) expression by human peripheral blood cd4-8- alpha/beta t cells demonstrates preferential use of several v beta genes and an invariant tcr alpha chain. J. Exp. Med. 1993, 178, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjer-Nielsen, L.; Patel, O.; Corbett, A.J.; Le Nours, J.; Meehan, B.; Liu, L.; Bhati, M.; Chen, Z.; Kostenko, L.; Reantragoon, R.; et al. Mr1 presents microbial vitamin b metabolites to mait cells. Nature 2012, 491, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Kjer-Nielsen, L.; Corbett, A.J.; Chen, Z.; Liu, L.; Mak, J.Y.; Godfrey, D.I.; Rossjohn, J.; Fairlie, D.P.; McCluskey, J.; Eckle, S.B. An overview on the identification of mait cell antigens. Immunol. Cell Biol. 2018, 96, 573–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McWilliam, H.E.G.; Mak, J.Y.W.; Awad, W.; Zorkau, M.; Cruz-Gomez, S.; Lim, H.J.; Yan, Y.; Wormald, S.; Dagley, L.F.; Eckle, S.B.G.; et al. Endoplasmic reticulum chaperones stabilize ligand-receptive mr1 molecules for efficient presentation of metabolite antigens. Proc. Natl. Acad. Sci. USA 2020, 117, 24974–24985. [Google Scholar] [CrossRef]
- Karamooz, E.; Harriff, M.J.; Lewinsohn, D.M. Mr1-dependent antigen presentation. Semin. Cell Dev. Biol. 2018, 84, 58–64. [Google Scholar] [CrossRef]
- Le Bourhis, L.; Martin, E.; Peguillet, I.; Guihot, A.; Froux, N.; Core, M.; Levy, E.; Dusseaux, M.; Meyssonnier, V.; Premel, V.; et al. Antimicrobial activity of mucosal-associated invariant t cells. Nat. Immunol. 2010, 11, 701–708. [Google Scholar] [CrossRef] [Green Version]
- Tastan, C.; Karhan, E.; Zhou, W.; Fleming, E.; Voigt, A.Y.; Yao, X.; Wang, L.; Horne, M.; Placek, L.; Kozhaya, L.; et al. Tuning of human mait cell activation by commensal bacteria species and mr1-dependent t-cell presentation. Mucosal. Immunol. 2018, 11, 1591–1605. [Google Scholar] [CrossRef] [Green Version]
- Reantragoon, R.; Kjer-Nielsen, L.; Patel, O.; Chen, Z.; Illing, P.T.; Bhati, M.; Kostenko, L.; Bharadwaj, M.; Meehan, B.; Hansen, T.H.; et al. Structural insight into mr1-mediated recognition of the mucosal associated invariant t cell receptor. J. Exp. Med. 2012, 209, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Ussher, J.E.; Bilton, M.; Attwod, E.; Shadwell, J.; Richardson, R.; de Lara, C.; Mettke, E.; Kurioka, A.; Hansen, T.H.; Klenerman, P.; et al. Cd161++ cd8+ t cells, including the mait cell subset, are specifically activated by il-12+il-18 in a tcr-independent manner. Eur. J. Immunol. 2014, 44, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Boulouis, C.; Sia, W.R.; Gulam, M.Y.; Teo, J.Q.M.; Png, Y.T.; Phan, T.K.; Mak, J.Y.W.; Fairlie, D.P.; Poon, I.K.H.; Koh, T.H.; et al. Human mait cell cytolytic effector proteins synergize to overcome carbapenem resistance in escherichia coli. PLoS Biol. 2020, 18, e3000644. [Google Scholar] [CrossRef] [PubMed]
- Dias, J.; Sobkowiak, M.J.; Sandberg, J.K.; Leeansyah, E. Human mait-cell responses to escherichia coli: Activation, cytokine production, proliferation, and cytotoxicity. J. Leukoc. Biol. 2016, 100, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Leeansyah, E.; Svard, J.; Dias, J.; Buggert, M.; Nystrom, J.; Quigley, M.F.; Moll, M.; Sonnerborg, A.; Nowak, P.; Sandberg, J.K. Arming of mait cell cytolytic antimicrobial activity is induced by il-7 and defective in hiv-1 infection. PLoS Pathog. 2015, 11, e1005072. [Google Scholar] [CrossRef]
- Dusseaux, M.; Martin, E.; Serriari, N.; Peguillet, I.; Premel, V.; Louis, D.; Milder, M.; Le Bourhis, L.; Soudais, C.; Treiner, E.; et al. Human mait cells are xenobiotic-resistant, tissue-targeted, cd161hi il-17-secreting t cells. Blood 2011, 117, 1250–1259. [Google Scholar] [CrossRef]
- Kurioka, A.; Walker, L.J.; Klenerman, P.; Willberg, C.B. Mait cells: New guardians of the liver. Clin. Transl. Immunol. 2016, 5, e98. [Google Scholar] [CrossRef]
- Mondelli, M.U. Do liver mucosa-associated invariant t cells delay hepatitis c progression, or are they innocent bystanders? Gastroenterology 2017, 153, 1185–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toubal, A.; Lehuen, A. Lights on mait cells, a new immune player in liver diseases. J. Hepatol. 2016, 64, 1008–1010. [Google Scholar] [CrossRef] [Green Version]
- Keller, A.N.; Eckle, S.B.; Xu, W.; Liu, L.; Hughes, V.A.; Mak, J.Y.; Meehan, B.S.; Pediongco, T.; Birkinshaw, R.W.; Chen, Z.; et al. Drugs and drug-like molecules can modulate the function of mucosal-associated invariant t cells. Nat. Immunol. 2017, 18, 402–411. [Google Scholar] [CrossRef]
- Corbett, A.J.; Eckle, S.B.; Birkinshaw, R.W.; Liu, L.; Patel, O.; Mahony, J.; Chen, Z.; Reantragoon, R.; Meehan, B.; Cao, H.; et al. T-cell activation by transitory neo-antigens derived from distinct microbial pathways. Nature 2014, 509, 361–365. [Google Scholar] [CrossRef]
- Eckle, S.B.; Corbett, A.J.; Keller, A.N.; Chen, Z.; Godfrey, D.I.; Liu, L.; Mak, J.Y.; Fairlie, D.P.; Rossjohn, J.; McCluskey, J. Recognition of vitamin b precursors and byproducts by mucosal associated invariant t cells. J. Biol. Chem. 2015, 290, 30204–30211. [Google Scholar] [CrossRef] [Green Version]
- Gold, M.C.; Cerri, S.; Smyk-Pearson, S.; Cansler, M.E.; Vogt, T.M.; Delepine, J.; Winata, E.; Swarbrick, G.M.; Chua, W.J.; Yu, Y.Y.; et al. Human mucosal associated invariant t cells detect bacterially infected cells. PLoS Biol. 2010, 8, e1000407. [Google Scholar] [CrossRef] [Green Version]
- Gold, M.C.; McLaren, J.E.; Reistetter, J.A.; Smyk-Pearson, S.; Ladell, K.; Swarbrick, G.M.; Yu, Y.Y.; Hansen, T.H.; Lund, O.; Nielsen, M.; et al. Mr1-restricted mait cells display ligand discrimination and pathogen selectivity through distinct t cell receptor usage. J. Exp. Med. 2014, 211, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Eckle, S.B.; Birkinshaw, R.W.; Kostenko, L.; Corbett, A.J.; McWilliam, H.E.; Reantragoon, R.; Chen, Z.; Gherardin, N.A.; Beddoe, T.; Liu, L.; et al. A molecular basis underpinning the t cell receptor heterogeneity of mucosal-associated invariant t cells. J. Exp. Med. 2014, 211, 1585–1600. [Google Scholar] [CrossRef] [PubMed]
- Soudais, C.; Samassa, F.; Sarkis, M.; Le Bourhis, L.; Bessoles, S.; Blanot, D.; Herve, M.; Schmidt, F.; Mengin-Lecreulx, D.; Lantz, O. In vitro and in vivo analysis of the gram-negative bacteria-derived riboflavin precursor derivatives activating mouse mait cells. J. Immunol. 2015, 194, 4641–4649. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Wang, H.; D’Souza, C.; Sun, S.; Kostenko, L.; Eckle, S.B.; Meehan, B.S.; Jackson, D.C.; Strugnell, R.A.; Cao, H.; et al. Mucosal-associated invariant t-cell activation and accumulation after in vivo infection depends on microbial riboflavin synthesis and co-stimulatory signals. Mucosal. Immunol. 2017, 10, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Huang, B.; Jiang, X.; Chen, W.; Zhang, J.; Wei, Y.; Chen, Y.; Lian, M.; Bian, Z.; Miao, Q.; et al. Mucosal-associated invariant t cells improve nonalcoholic fatty liver disease through regulating macrophage polarization. Front. Immunol. 2018, 9, 1994. [Google Scholar] [CrossRef]
- Toubal, A.; Kiaf, B.; Beaudoin, L.; Cagninacci, L.; Rhimi, M.; Fruchet, B.; da Silva, J.; Corbett, A.J.; Simoni, Y.; Lantz, O.; et al. Mucosal-associated invariant t cells promote inflammation and intestinal dysbiosis leading to metabolic dysfunction during obesity. Nat. Commun. 2020, 11, 3755. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Mazmanian, S.K. Innate immune recognition of the microbiota promotes host-microbial symbiosis. Nat. Immunol. 2013, 14, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Takeda, K. Host-microbiota interactions in rheumatoid arthritis. Exp. Mol. Med. 2019, 51, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Valitutti, F.; Cucchiara, S.; Fasano, A. Celiac disease and the microbiome. Nutrients 2019, 11, 2403. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Sun, K.; Wu, Y.; Yang, Y.; Tso, P.; Wu, Z. Interactions between intestinal microbiota and host immune response in inflammatory bowel disease. Front. Immunol. 2017, 8, 942. [Google Scholar] [CrossRef] [Green Version]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, Y.; Sato, T.; Ohteki, T. Commensal gram-positive bacteria initiates colitis by inducing monocyte/macrophage mobilization. Mucosal. Immunol. 2015, 8, 152–160. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Francino, M.P. Early development of the gut microbiota and immune health. Pathogens 2014, 3, 769–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanahan, F. Physiological basis for novel drug therapies used to treat the inflammatory bowel diseases i. Pathophysiological basis and prospects for probiotic therapy in inflammatory bowel disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G417–G421. [Google Scholar] [CrossRef] [Green Version]
- Martin, E.; Treiner, E.; Duban, L.; Guerri, L.; Laude, H.; Toly, C.; Premel, V.; Devys, A.; Moura, I.C.; Tilloy, F.; et al. Stepwise development of mait cells in mouse and human. PLoS Biol. 2009, 7, e54. [Google Scholar] [CrossRef]
- Koay, H.F.; Gherardin, N.A.; Enders, A.; Loh, L.; Mackay, L.K.; Almeida, C.F.; Russ, B.E.; Nold-Petry, C.A.; Nold, M.F.; Bedoui, S.; et al. A three-stage intrathymic development pathway for the mucosal-associated invariant t cell lineage. Nat. Immunol. 2016, 17, 1300–1311. [Google Scholar] [CrossRef]
- Rahimpour, A.; Koay, H.F.; Enders, A.; Clanchy, R.; Eckle, S.B.; Meehan, B.; Chen, Z.; Whittle, B.; Liu, L.; Fairlie, D.P.; et al. Identification of phenotypically and functionally heterogeneous mouse mucosal-associated invariant t cells using mr1 tetramers. J. Exp. Med. 2015, 212, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.Z.; Jo, J.; Tan, A.T.; Sandalova, E.; Chia, A.; Tan, K.C.; Lee, K.H.; Gehring, A.J.; De Libero, G.; Bertoletti, A. Il-7 licenses activation of human liver intrasinusoidal mucosal-associated invariant t cells. J. Immunol. 2013, 190, 3142–3152. [Google Scholar] [CrossRef]
- Merlini, E.; Cerrone, M.; van Wilgenburg, B.; Swadling, L.; Cannizzo, E.S.; d’Arminio Monforte, A.; Klenerman, P.; Marchetti, G. Association between impaired valpha7.2+cd161++cd8+ (mait) and valpha7.2+cd161-cd8+ t-cell populations and gut dysbiosis in chronically hiv- and/or hcv-infected patients. Front. Microbiol. 2019, 10, 1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimaraes, V.; Sokol, H.; Dore, J.; Corthier, G.; Furet, J.P. The firmicutes/bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Krause, J.L.; Schape, S.S.; Schattenberg, F.; Muller, S.; Ackermann, G.; Rolle-Kampczyk, U.E.; Jehmlich, N.; Pierzchalski, A.; von Bergen, M.; Herberth, G. The activation of mucosal-associated invariant t (mait) cells is affected by microbial diversity and riboflavin utilization in vitro. Front. Microbiol. 2020, 11, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Ye, L.; Li, J.; Jin, L.; Wang, W.; Li, S.; Bao, M.; Wu, S.; Li, L.; Geng, B.; et al. Metagenomic and metabolomic analyses unveil dysbiosis of gut microbiota in chronic heart failure patients. Sci. Rep. 2018, 8, 635. [Google Scholar] [CrossRef] [PubMed]
- Jie, Z.; Xia, H.; Zhong, S.L.; Feng, Q.; Li, S.; Liang, S.; Zhong, H.; Liu, Z.; Gao, Y.; Zhao, H.; et al. The gut microbiome in atherosclerotic cardiovascular disease. Nat. Commun. 2017, 8, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xu, H.; Li, Y.; Jiang, Y.; Hu, Y.; Liu, T.; Tian, X.; Zhao, X.; Zhu, Y.; Wang, S.; et al. Alterations of gut microbiota contribute to the progression of unruptured intracranial aneurysms. Nat. Commun. 2020, 11, 3218. [Google Scholar] [CrossRef]
- Gulden, E.; Palm, N.; Herold, K.C. Mait cells: A link between gut integrity and type 1 diabetes. Cell Metab. 2017, 26, 813–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottcher, K.; Rombouts, K.; Saffioti, F.; Roccarina, D.; Rosselli, M.; Hall, A.; Luong, T.; Tsochatzis, E.A.; Thorburn, D.; Pinzani, M. Mait cells are chronically activated in patients with autoimmune liver disease and promote profibrogenic hepatic stellate cell activation. Hepatology 2018, 68, 172–186. [Google Scholar] [CrossRef] [Green Version]
- Chiba, A.; Tamura, N.; Yoshikiyo, K.; Murayama, G.; Kitagaichi, M.; Yamaji, K.; Takasaki, Y.; Miyake, S. Activation status of mucosal-associated invariant t cells reflects disease activity and pathology of systemic lupus erythematosus. Arthritis Res. Ther. 2017, 19, 58. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Yoo, S.J.; Kang, S.W.; Kwon, J.; Choi, I.; Lee, C.H. Tnfalpha and il-1beta in the synovial fluid facilitate mucosal-associated invariant t (mait) cell migration. Cytokine 2017, 99, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Parrot, T.; Gorin, J.B.; Ponzetta, A.; Maleki, K.T.; Kammann, T.; Emgard, J.; Perez-Potti, A.; Sekine, T.; Rivera-Ballesteros, O.; Karolinska, C.-S.G.; et al. Mait cell activation and dynamics associated with COVID-19 disease severity. Sci. Immunol. 2020, 5, 1670. [Google Scholar] [CrossRef]
- D’Souza, C.; Pediongco, T.; Wang, H.; Scheerlinck, J.Y.; Kostenko, L.; Esterbauer, R.; Stent, A.W.; Eckle, S.B.G.; Meehan, B.S.; Strugnell, R.A.; et al. Mucosal-associated invariant t cells augment immunopathology and gastritis in chronic helicobacter pylori infection. J. Immunol. 2018, 200, 1901–1916. [Google Scholar]
- Jeffery, H.C.; van Wilgenburg, B.; Kurioka, A.; Parekh, K.; Stirling, K.; Roberts, S.; Dutton, E.E.; Hunter, S.; Geh, D.; Braitch, M.K.; et al. Biliary epithelium and liver b cells exposed to bacteria activate intrahepatic mait cells through mr1. J. Hepatol. 2016, 64, 1118–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Lian, M.; Li, Y.; Zhang, W.; Wang, Q.; Wei, Y.; Zhang, J.; Chen, W.; Xiao, X.; Miao, Q.; et al. The immunobiology of mucosal-associated invariant t cell (mait) function in primary biliary cholangitis: Regulation by cholic acid-induced interleukin-7. J. Autoimmun. 2018, 90, 64–75. [Google Scholar] [CrossRef]
- Hegde, P.; Weiss, E.; Paradis, V.; Wan, J.; Mabire, M.; Sukriti, S.; Rautou, P.E.; Albuquerque, M.; Picq, O.; Gupta, A.C.; et al. Mucosal-associated invariant t cells are a profibrogenic immune cell population in the liver. Nat. Commun. 2018, 9, 2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotersztajn, S.; Julien, B.; Teixeira-Clerc, F.; Grenard, P.; Mallat, A. Hepatic fibrosis: Molecular mechanisms and drug targets. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 605–628. [Google Scholar] [CrossRef]
- Mallat, A.; Lotersztajn, S. Cellular mechanisms of tissue fibrosis. 5. Novel insights into liver fibrosis. Am. J. Physiol. Cell Physiol. 2013, 305, C789–C799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Lin, E.L.; Liangpunsakul, S.; Lan, J.; Chalasani, S.; Rane, S.; Puri, P.; Kamath, P.S.; Sanyal, A.J.; Shah, V.H.; et al. Alcohol abstinence does not fully reverse abnormalities of mucosal-associated invariant t cells in the blood of patients with alcoholic hepatitis. Clin. Transl. Gastroenterol. 2019, 10, e00052. [Google Scholar] [CrossRef]
- Duan, M.; Goswami, S.; Shi, J.Y.; Wu, L.J.; Wang, X.Y.; Ma, J.Q.; Zhang, Z.; Shi, Y.; Ma, L.J.; Zhang, S.; et al. Activated and exhausted mait cells foster disease progression and indicate poor outcome in hepatocellular carcinoma. Clin. Cancer Res. 2019, 25, 3304–3316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, Y.K.; Saeidi, A.; Tan, H.Y.; Rosmawati, M.; Enstrom, P.F.; Batran, R.A.; Vasuki, V.; Chattopadhyay, I.; Murugesan, A.; Vignesh, R.; et al. Hyper-expression of pd-1 is associated with the levels of exhausted and dysfunctional phenotypes of circulating cd161(++) tcr ivalpha7.2(+) mucosal-associated invariant t cells in chronic hepatitis b virus infection. Front. Immunol. 2018, 9, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.; Zheng, L.; Yoo, J.K.; Guo, H.; Zhang, Y.; Guo, X.; Kang, B.; Hu, R.; Huang, J.Y.; Zhang, Q.; et al. Landscape of infiltrating t cells in liver cancer revealed by single-cell sequencing. Cell 2017, 169, 1342–1356. [Google Scholar] [CrossRef] [Green Version]
- Bolte, F.J.; O’Keefe, A.C.; Webb, L.M.; Serti, E.; Rivera, E.; Liang, T.J.; Ghany, M.; Rehermann, B. Intra-hepatic depletion of mucosal-associated invariant t cells in hepatitis c virus-induced liver inflammation. Gastroenterology 2017, 153, 1392–1403. [Google Scholar] [CrossRef] [Green Version]
- Spaan, M.; Hullegie, S.J.; Beudeker, B.J.; Kreefft, K.; van Oord, G.W.; Groothuismink, Z.M.; van Tilborg, M.; Rijnders, B.; de Knegt, R.J.; Claassen, M.A.; et al. Frequencies of circulating mait cells are diminished in chronic hcv, hiv and hcv/hiv co-infection and do not recover during therapy. PLoS ONE 2016, 11, e0159243. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, P.; Wang, W.; Tan, X.; Liu, C.; Chen, Y.; Pei, R.; Cheng, X.; Wu, M.; Guo, Q.; et al. Mait cell dysregulation correlates with conjugated bilirubin level in chronic hepatitis b virus infection. Hepatology 2020. [Google Scholar] [CrossRef]
- Yong, Y.K.; Tan, H.Y.; Saeidi, A.; Rosmawati, M.; Atiya, N.; Ansari, A.W.; Rajarajeswaran, J.; Vadivelu, J.; Velu, V.; Larsson, M.; et al. Decrease of cd69 levels on tcr valpha7.2(+) cd4(+) innate-like lymphocytes is associated with impaired cytotoxic functions in chronic hepatitis b virus-infected patients. Innate Immun. 2017, 23, 459–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renand, A.; Habes, S.; Mosnier, J.F.; Auble, H.; Judor, J.P.; Vince, N.; Hulin, P.; Nedellec, S.; Metairie, S.; Archambeaud, I.; et al. Immune alterations in patients with type 1 autoimmune hepatitis persist upon standard immunosuppressive treatment. Hepatol. Commun. 2018, 2, 968–981. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.K.; Cho, Y.N.; Park, K.J.; Kwak, H.D.; Jin, H.M.; Park, S.Y.; Kim, H.S.; Kee, S.J.; Park, Y.W. Activation, deficiency, and reduced ifn-gamma production of mucosal-associated invariant t cells in patients with inflammatory bowel disease. J. Innate Immun. 2020, 12, 422–434. [Google Scholar] [CrossRef]
- Serriari, N.E.; Eoche, M.; Lamotte, L.; Lion, J.; Fumery, M.; Marcelo, P.; Chatelain, D.; Barre, A.; Nguyen-Khac, E.; Lantz, O.; et al. Innate mucosal-associated invariant t (mait) cells are activated in inflammatory bowel diseases. Clin. Exp. Immunol. 2014, 176, 266–274. [Google Scholar] [CrossRef]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of nafld and nash: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef]
- Younossi, Z.M. Non-alcoholic fatty liver disease—A global public health perspective. J. Hepatol. 2019, 70, 531–544. [Google Scholar] [CrossRef] [Green Version]
- Leung, C.; Rivera, L.; Furness, J.B.; Angus, P.W. The role of the gut microbiota in nafld. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 412–425. [Google Scholar] [CrossRef]
- Lamichhane, R.; Ussher, J.E. Expression and trafficking of mr1. Immunology 2017, 151, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Seitz, H.K.; Bataller, R.; Cortez-Pinto, H.; Gao, B.; Gual, A.; Lackner, C.; Mathurin, P.; Mueller, S.; Szabo, G.; Tsukamoto, H. Alcoholic liver disease. Nat. Rev. Dis. Primers 2018, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Betrapally, N.S.; Gillevet, P.M.; Bajaj, J.S. Changes in the intestinal microbiome and alcoholic and nonalcoholic liver diseases: Causes or effects? Gastroenterology 2016, 150, 1745–1755. [Google Scholar] [CrossRef] [Green Version]
- Llopis, M.; Cassard, A.M.; Wrzosek, L.; Boschat, L.; Bruneau, A.; Ferrere, G.; Puchois, V.; Martin, J.C.; Lepage, P.; Le Roy, T.; et al. Intestinal microbiota contributes to individual susceptibility to alcoholic liver disease. Gut 2016, 65, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Riva, A.; Patel, V.; Kurioka, A.; Jeffery, H.C.; Wright, G.; Tarff, S.; Shawcross, D.; Ryan, J.M.; Evans, A.; Azarian, S.; et al. Mucosa-associated invariant t cells link intestinal immunity with antibacterial immune defects in alcoholic liver disease. Gut 2018, 67, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the global burden of disease study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Franciszkiewicz, K.; Salou, M.; Legoux, F.; Zhou, Q.; Cui, Y.; Bessoles, S.; Lantz, O. Mhc class i-related molecule, mr1, and mucosal-associated invariant t cells. Immunol. Rev. 2016, 272, 120–138. [Google Scholar] [CrossRef]
- Chattergoon, M.A.; Latanich, R.; Quinn, J.; Winter, M.E.; Buckheit, R.W., 3rd; Blankson, J.N.; Pardoll, D.; Cox, A.L. Hiv and hcv activate the inflammasome in monocytes and macrophages via endosomal toll-like receptors without induction of type 1 interferon. PLoS Pathog. 2014, 10, e1004082. [Google Scholar] [CrossRef]
- Serti, E.; Werner, J.M.; Chattergoon, M.; Cox, A.L.; Lohmann, V.; Rehermann, B. Monocytes activate natural killer cells via inflammasome-induced interleukin 18 in response to hepatitis c virus replication. Gastroenterology 2014, 147, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Study Models | Frequency and Phenotype of MAIT Cells | Remarks | Reference |
|---|---|---|---|---|
| NAFLD | Mr1 KO mice model -NAFLD Patients | Frequency: Blood↓, Liver↑ Markers: Blood CD69, CD38, HLA-DR, PD-1, CXCR6, CCR5↑; Liver MR1 (Kupffer cells) ↑ | Deficiency triggers steatohepatitis | [32] |
| ALD | AH patients | Frequency: Blood↓, Liver↑ Markers: CD161↑, CD69↑, CD38↑, HLA-DR↑, Granzyme B↑ Activation factors ↑ | Alcohol cessation reverses MAIT cell abnormalities | [66] |
| Cirrhosis | -Cirrhotic ALD patients -CCl4-induced liver injury mice model | Frequency: Blood↓, Liver↑ Markers: Blood CD25↑, CD69↑, IL-17↑, Granzyme B↑, IFN-γ↑, TNF↑; Liver: IL-17 ↑, MR1 ↑ | -Profibrogenic -Macrophage recruitment | [63] |
| HCC | HCC patients | Frequency: Blood↓, Tumor ↓ Markers: Tumor CD38↑, HLA-DR↑, CCR6↓, CXCR6↓, CCR9↓ Blood CD45RO; Liver CD28↓, CD127↓ | Infiltration of tumor by MAIT results in dismal clinical outcomes | [67,68,69] |
| HCV | Chronic HCV infection | Frequency: Blood↓, Liver↓ Markers: Granzyme B↓, IFN-γ↓, CD69↓, IL-12↑, IL-18↑, | Inverse correlation between MAIT cell frequency and liver inflammation | [70,71] |
| HBV | Chronic HBV infection | Frequency: Blood↓, Liver↓ Markers: Granzyme B↓, IFN- γ↓, CD69↓, | Impaired cytotoxic function with reduced CD69 expression | [72,73] |
| AILD | AILD patients | Frequency: Blood↓, Liver↓ Markers: CD38↓, HLA-DR↓, CTLA-4↓, IFN-γ↓, Foxp3↑, CD8↑, CD161↑, Granzyme B↑ | Promote Profibrogenic Hepatic Stellate Cell Activation | [56,74] |
| IBD | IBD patients | Frequency: Blood↓, Inflamed tissue↑ Markers: IFN-γ↓, IL-17↑, CD69↑, PD-1↑, annexin V↑, CCL20↑, CXCL10↑, CXCL16↑, CCL25↑ | MAIT cells migrate to inflamed tissue | [75,76] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebru, Y.A.; Choi, M.R.; Raja, G.; Gupta, H.; Sharma, S.P.; Choi, Y.R.; Kim, H.S.; Yoon, S.J.; Kim, D.J.; Suk, K.T. Pathophysiological Roles of Mucosal-Associated Invariant T Cells in the Context of Gut Microbiota-Liver Axis. Microorganisms 2021, 9, 296. https://doi.org/10.3390/microorganisms9020296
Gebru YA, Choi MR, Raja G, Gupta H, Sharma SP, Choi YR, Kim HS, Yoon SJ, Kim DJ, Suk KT. Pathophysiological Roles of Mucosal-Associated Invariant T Cells in the Context of Gut Microbiota-Liver Axis. Microorganisms. 2021; 9(2):296. https://doi.org/10.3390/microorganisms9020296
Chicago/Turabian StyleGebru, Yoseph Asmelash, Mi Ran Choi, Ganesan Raja, Haripriya Gupta, Satya Priya Sharma, Ye Rin Choi, Hyeong Seop Kim, Sang Jun Yoon, Dong Joon Kim, and Ki Tae Suk. 2021. "Pathophysiological Roles of Mucosal-Associated Invariant T Cells in the Context of Gut Microbiota-Liver Axis" Microorganisms 9, no. 2: 296. https://doi.org/10.3390/microorganisms9020296
APA StyleGebru, Y. A., Choi, M. R., Raja, G., Gupta, H., Sharma, S. P., Choi, Y. R., Kim, H. S., Yoon, S. J., Kim, D. J., & Suk, K. T. (2021). Pathophysiological Roles of Mucosal-Associated Invariant T Cells in the Context of Gut Microbiota-Liver Axis. Microorganisms, 9(2), 296. https://doi.org/10.3390/microorganisms9020296