
Changes in the Microbial Composition of the Cecum and Histomorphometric Analysis of Its Epithelium in Broilers Fed with Feed Mixture Containing Fermented Rapeseed Meal

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fermentation of Rapeseed Meal by the Bacillus subtilis 87Y and 67 Strains in 50 kg SSF Bioreactor
2.2. Broiler Chicken Population and Experimental Design
2.3. Nutrition
2.4. Sampling to Determine the Microbiome of Broiler Chickens
2.5. Detection and/or Quantification of Cecal Microorganisms
2.6. MALDI-TOF-MS Analysis
2.7. Metagenome Analysis
2.8. Histomorphometrics of the Cecum
2.9. Statistical Analyses
3. Results
3.1. Detection of Microorganisms Using RAPID Salmonella agar and Identification by MALDI-TOF-MS
3.2. Microorganism Detection by Chromogenic Agar Media and Identification by MALDI-TOF-MS: Detection of Microorganisms Using StrepB Select Agar
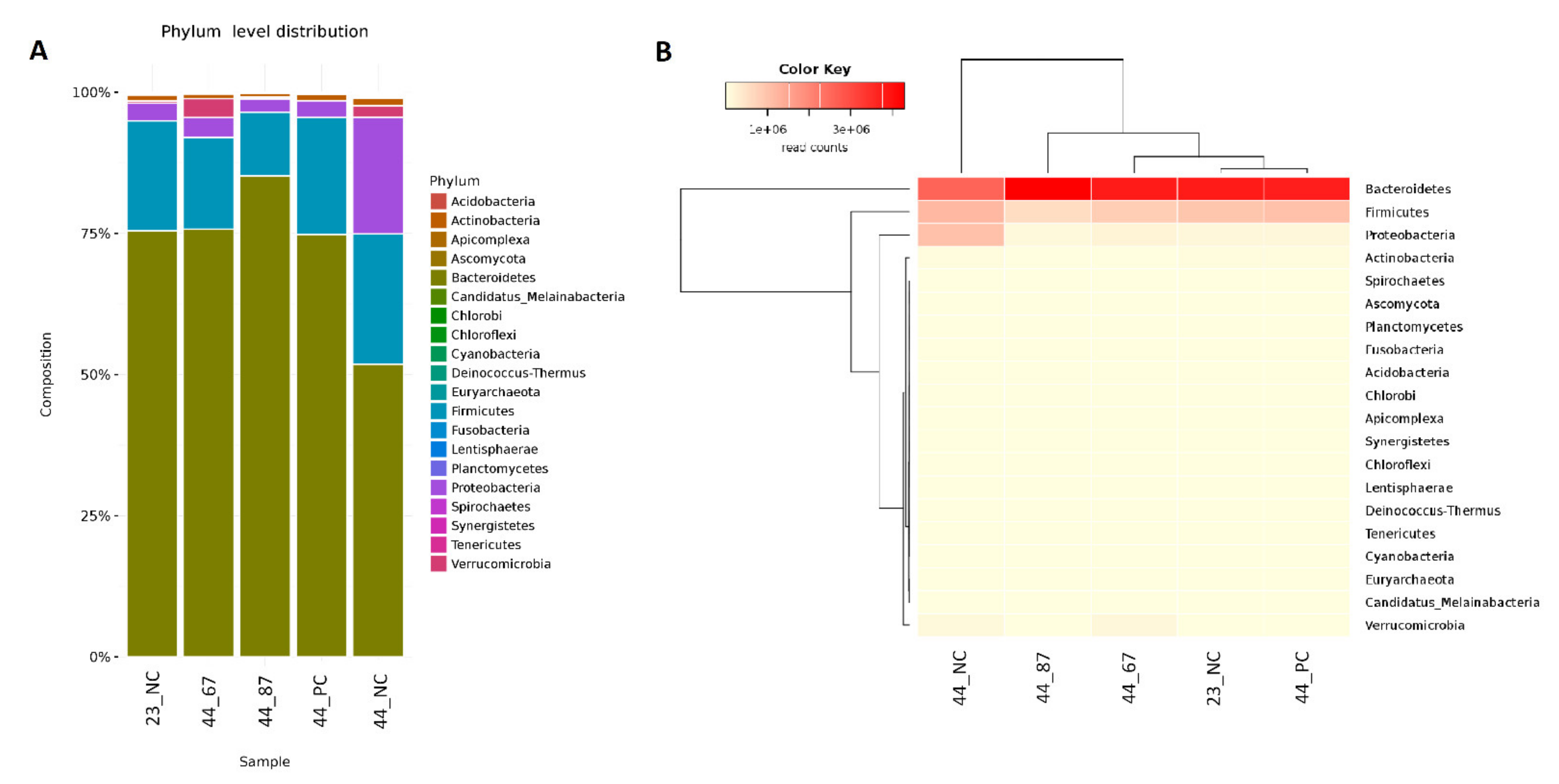
3.3. Metagenome Analysis
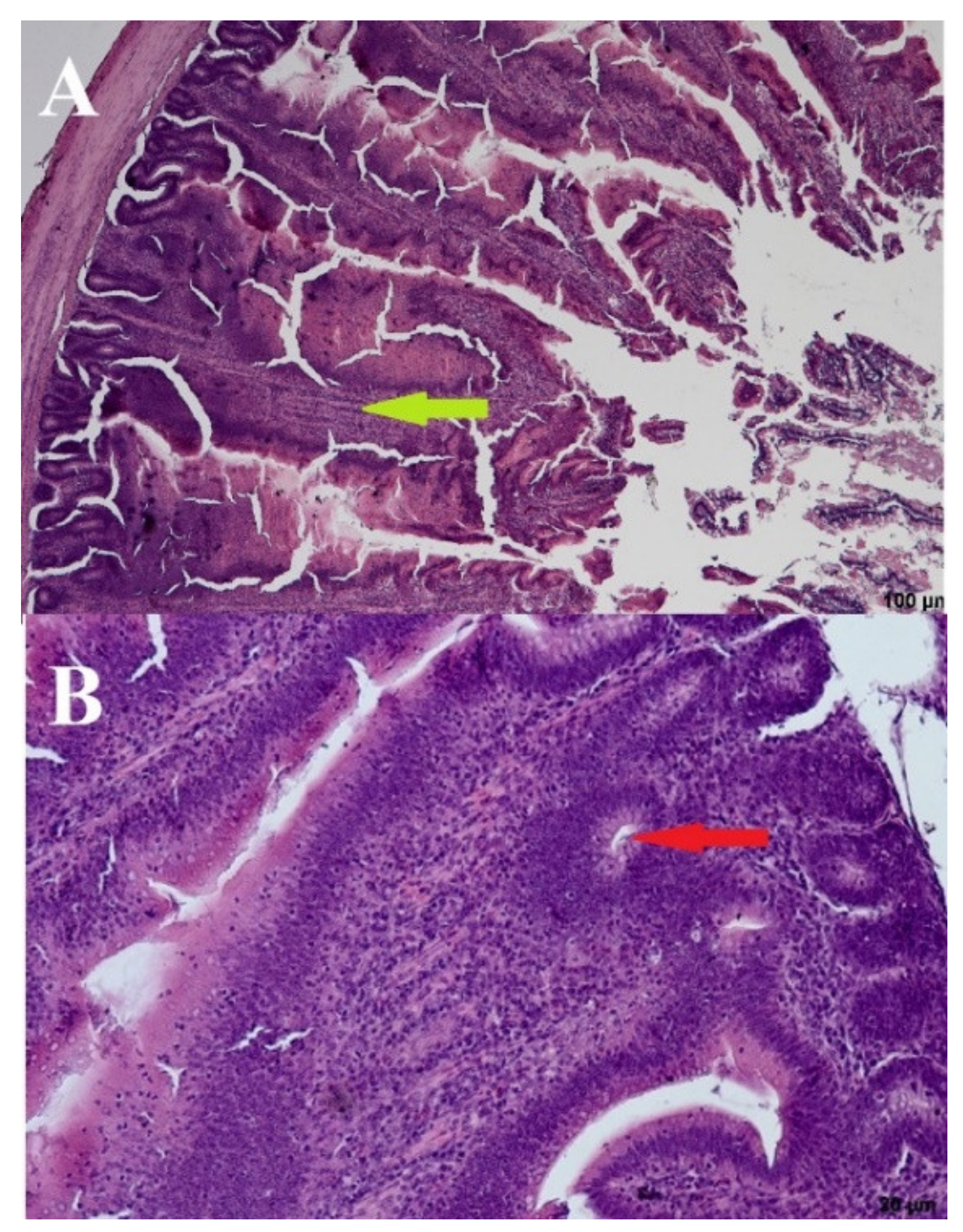
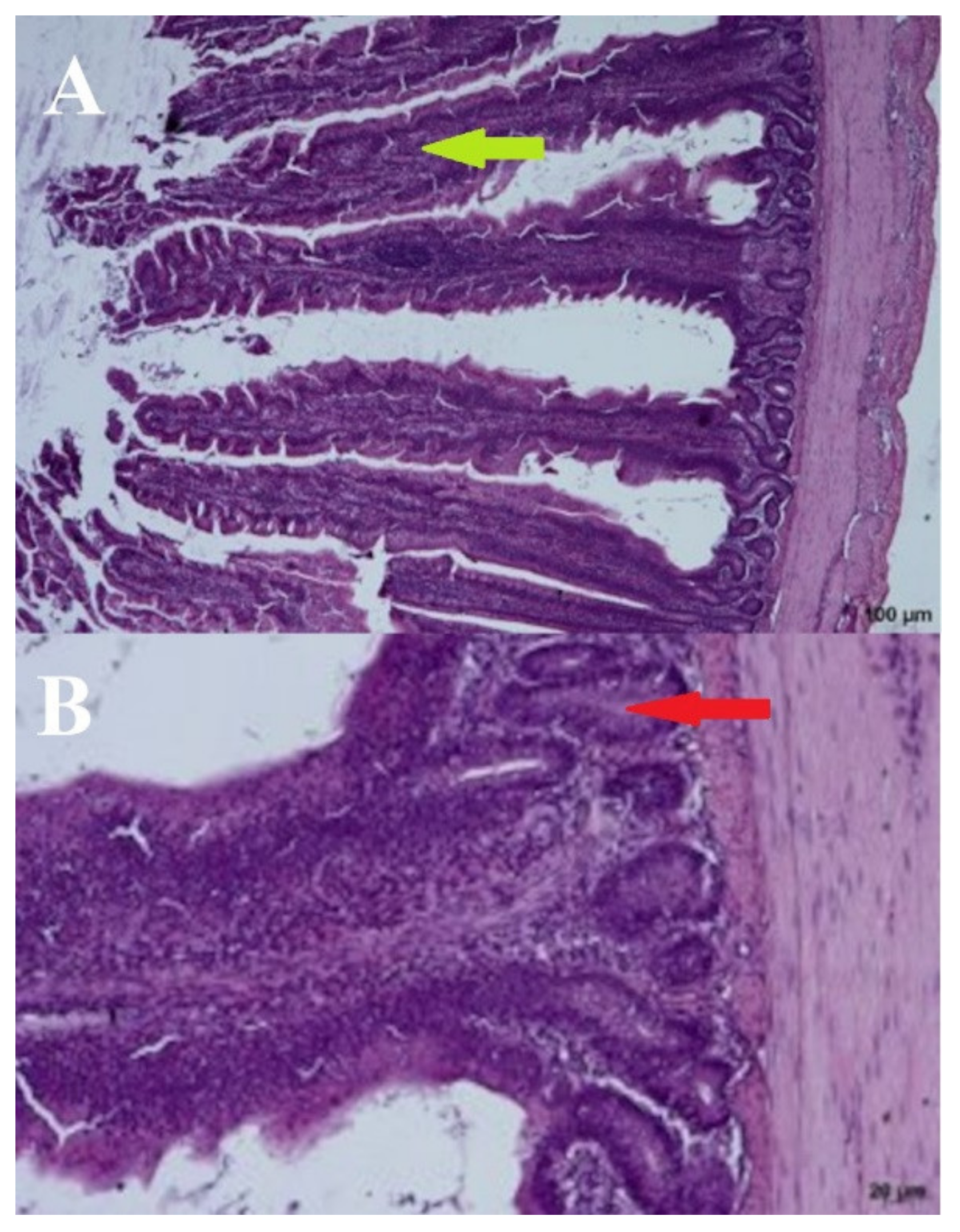
3.4. Histomorphometric Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wei, S.; Morrison, M.; Yu, Z. Bacterial census of poultry intestinal microbiome. Poult. Sci. 2013, 92, 671–683. [Google Scholar] [CrossRef]
- Brisbin, J.T.; Gong, J.; Orouji, S.; Esufali, J.; Mallick, A.I.; Parvizi, P.; Shewen, P.E.; Sharif, S. Oral treatment of chickens with lactobacilli influences elicitation of immune responses. Clin. Vaccine Immunol. 2011, 18, 1447–1455. [Google Scholar] [CrossRef]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the chicken gastrointestinal tract: Influence on health, productivity and disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef] [PubMed]
- Obst, B.S.; Diamond, J.M. Interspecific variation in sugar and amino acid transport by the avian cecum. J. Exp. Zool. 1989, 252, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Clench, M.; Mathias, J. The avian cecum: A review. Wilson Bull. Wilson Ornithol. Soc. 1995, 107, 93–121. [Google Scholar]
- Miterau, B.N.; Blair, R.; Bell, J.M.; Reichert, R.D. Tannin and fiber contents of rapeseed and canola hulls. Can. J. Anim. Sci. 1982, 62, 661–663. [Google Scholar] [CrossRef]
- Mejicanos, G.; Sanjayan, N.; Kim, I.H.; Nyachoti, C.M. Recent advances in canola meal utilization in swine nutrition. J. Anim. Sci. Technol. 2016, 58, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Wang, Y.; Li, A.; Wang, Z.; Zhang, X.; Yun, T.; Qiu, L.; Yin, Y. Effects of fermented rapeseed meal on antioxidant functions, serum biochemical parameters and intestinal morphology in broilers. Food Agric. Immunol. 2016, 27, 182–193. [Google Scholar] [CrossRef]
- Gaggìa, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 2010, 141, S15–S28. [Google Scholar] [CrossRef] [PubMed]
- Kabir, S.M.L. The role of probiotics in the poultry industry. Int. J. Mol. Sci. 2009, 10, 3531–3546. [Google Scholar] [CrossRef]
- Kim, J.S.; Ingale, S.L.; Kim, Y.W.; Kim, K.H.; Sen, S.; Ryu, M.H.; Lohakare, J.D.; Kwon, I.K.; Chae, B.J. Effect of supplementation of multi-microbe probiotic product on growth performance, apparent digestibility, cecal microbiota and small intestinal morphology of broilers. J. Anim. Physiol. Anim. Nutr. (Berl.) 2012, 96, 618–626. [Google Scholar] [CrossRef]
- Sun, H.; Tang, J.; Yao, X.; Wu, Y.; Wang, X.; Feng, J. Effects of dietary inclusion of fermented cottonseed meal on growth, cecal microbial population, small intestinal morphology, and digestive enzyme activity of broilers. Trop. Anim. Health Prod. 2013, 45, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Konkol, D.; Szmigiel, I.; Domżał-Kędzia, M.; Kułażyński, M.; Krasowska, A.; Opaliński, S.; Korczyński, M.; Łukaszewicz, M. Biotransformation of rapeseed meal leading to production of polymers, biosurfactants, and fodder. Bioorg. Chem. 2019, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jajor, P.; Piłakowska-Pietras, D.; Krasowska, A.; Łukaszewicz, M. Surfactin analogues produced by Bacillus subtilis strains grown on rapeseed cake. J. Mol. Struct. 2016, 1126, 141–146. [Google Scholar] [CrossRef]
- Szmigiel, I.; Suchodolski, J.; Łukaszewicz., M.; Krasowska, A. The influence of Bacillus subtilis 87Y isolated from Eisenia fetida on the growth of pathogenic and probiotic microorganisms. Biomass Conv. Bioref. 2019. [Google Scholar] [CrossRef] [Green Version]
- Aviagen Ross Broiler Management Handbook 2018. Available online: http://online.anyflip.com/kmgi/zqpr/index.html#p=4 (accessed on 16 November 2020).
- Smulikowska, S.; Rutkowski, A. Normy żywienia drobiu; Instytut Fizjologii i Żywienia Zwierząt PAN: Jabłonna, Poland, 2005; ISBN 83-917097-7-9. [Google Scholar]
- Breitwieser, F.P.; Baker, D.N.; Salzberg, S.L. KrakenUniq: Confident and fast metagenomics classification using unique k-mer counts. Genome Biol. 2018, 19, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belote, B.L.; Soares, I.; Tujimoto-Silva, A.; Sanches, A.W.D.; Kraieski, A.L.; Santin, E. Applying I see inside histological methodology to evaluate gut health in broilers challenged with Eimeria. Vet. Parasitol. X 2019, 1, 100004. [Google Scholar] [CrossRef]
- Shah, M.; Zaneb, H.; Masood, S.; Khan, R.U.; Mobashar, M.; Khan, I.; Din, S.; Khan, M.S.; Rehman, H.U.; Tinelli, A. Single or Combined Applications of Zinc and Multi-strain Probiotic on Intestinal Histomorphology of Broilers Under Cyclic Heat Stress. Probiotics Antimicrob. Proteins 2019, 2–9. [Google Scholar] [CrossRef]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liljebjelke, K.A.; Hofacre, C.L.; White, D.G.; Ayers, S.; Lee, M.D.; Maurer, J.J. Diversity of antimicrobial resistance phenotypes in Salmonella isolated from commercial poultry farms. Front. Vet. Sci. 2017, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Tellez, G.; Latorre, J.D.; Ray, P.M.; Hernandez, X.; Hargis, B.M.; Ricke, S.C.; Kwon, Y.M. Salmonella excludes Salmonella in poultry: Confirming an old paradigm using conventional and barcode-tagging approaches. Front. Vet. Sci. 2018, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-M.; Wang, Y.; Su, L.-H.; Chiu, C.-H. Nontyphoid Salmonella Infection: Microbiology, Clinical Features, and Antimicrobial Therapy. Pediatr. Neonatol. 2013, 54, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisse, S.; Fevre, C.; Passet, V.; Issenhuth-Jeanjean, S.; Tournebize, R.; Diancourt, L.; Grimont, P. Virulent clones of Klebsiella pneumoniae: Identification and evolutionary scenario based on genomic and phenotypic characterization. PLoS ONE 2009, 4, e4982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, A.B.; de Cássia de Andrade Melo, R.; Maciel, M.A.V.; Lopes, A.C.S. Multidrug resistance genes, including blaKPC and blaCTX-M-2, among Klebsiella pneumoniae isolated in Recife, Brazil. Rev. Soc. Bras. Med. Trop. 2012, 45, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Hamza, E.; Dorgham, S.M.; Hamza, D.A. Carbapenemase-producing Klebsiella pneumoniae in broiler poultry farming in Egypt. J. Glob. Antimicrob. Resist. 2016, 7, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Shiri, Y.; Solouki, M.; Saeidi, S. Activity of some Iranian plant extracts against Multi-Drug Resistant human pathogens isolated from urinary tract infections. Zahedan J. Res. Med. Sci. 2014, 16, 50–54. [Google Scholar]
- Adesiyun, A.; Offiah, N.; Seepersadsingh, N.; Rodrigo, S.; Lashley, V.; Musai, L. Frequency and antimicrobial resistance of enteric bacteria with spoilage potential isolated from table eggs. Food Res. Int. 2006, 39, 212–219. [Google Scholar] [CrossRef]
- Sabarinath, A.; Guillaume, V.; Guillaume, B.; Mathew, V.; DeAllie, C. Bacterial contamination of commercial chicken eggs in Grenada, West Indies. West. Indian Vet. J. 2009, 9, 4–7. [Google Scholar]
- Musgrove, M.T.; Northcutt, J.K.; Jones, D.R.; Cox, N.A.; Harrison, M.A. Enterobacteriaceae and Related Organisms Isolated from Shell Eggs Collected During Commercial Processing. Poult. Sci. 2008, 87, 1211–1218. [Google Scholar] [CrossRef]
- Heres, L.; Engel, B.; van Knapen, F.; de Jong, M.; Wagenaar, J.; Urlings, H. Fermented liquid feed reduces susceptibility of broilers for Salmonella enteritidis. Poult. Sci. 2003, 82, 603–611. [Google Scholar] [CrossRef]
- Mellata, M. Human and avian extraintestinal pathogenic escherichia coli: Infections, zoonotic risks, and antibiotic resistance trends. Foodborne Pathog. Dis. 2013, 10, 916–932. [Google Scholar] [CrossRef] [Green Version]
- Arias, C.A.; Murray, B.E. The rise of the Enterococcus: Beyond vancomycin resistance. Nat. Rev. Microbiol. 2012, 10, 266–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolaia, V.; Espinosa-Gongora, C.; Guardabassi, L. Human health risks associated with antimicrobial-resistant enterococci and Staphylococcus aureus on poultry meat. Clin. Microbiol. Infect. 2016, 22, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Rinkinen, M.L.; Koort, J.M.K.; Ouwehand, A.C.; Westermarck, E.; Björkroth, K.J. Streptococcus alactolyticus is the dominating culturable lactic acid bacterium species in canine jejunum and feces of four fistulated dogs. FEMS Microbiol. Lett. 2004, 230, 35–39. [Google Scholar] [CrossRef] [Green Version]
- Almeida, P.; Railsback, J.; Gleason, J.B. A Rare Case of Streptococcus alactolyticus Infective Endocarditis Complicated by Septic Emboli and Mycotic Left Middle Cerebral Artery Aneurysm. Case Rep. Infect. Dis. 2016, 2016, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Cekmen, N.; Baysan, O.; Disbudak, E.; Gunt, C. A rare case of bacterial infective endocarditis caused by Streptococcus alactolyticus. Hear. Vessel. Transplant. 2019, 3, 109–133. [Google Scholar] [CrossRef]
- Al Marjani, M.F.; Abdul, J.; Salman, S.; Khudhaier, S.R.; Salim, M.Z.; Kadham, Z.A. First case of vancomycin resistant Streptococcus alactolyticus from raw milk in Baghdad-Iraq. Adv. Environ. Biol. 2016, 10, 4–9. [Google Scholar]
- Manafi, M.; Hedayati, M.; Mirzaie, S. Probiotic Bacillus species and Saccharomyces boulardii improve performance, gut histology and immunity in broiler chickens. South Afr. J. Anim. Sci. 2018, 48, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Buahom, J.; Siripornadulsil, S.; Siripornadulsil, W. Feeding with Single Strains Versus Mixed Cultures of Lactic Acid Bacteria and Bacillus subtilis KKU213 Affects the Bacterial Community and Growth Performance of Broiler Chickens. Arab. J. Sci. Eng. 2018, 43, 3417–3427. [Google Scholar] [CrossRef]
- Zou, X.; Xiao, R.; Li, H.; Liu, T.; Liao, Y.; Wang, Y.; Wu, S.; Li, Z. Effect of a novel strain of Lactobacillus brevis M8 and tea polyphenol diets on performance, meat quality and intestinal microbiota in broilers. Ital. J. Anim. Sci. 2018, 17, 396–407. [Google Scholar] [CrossRef] [Green Version]
- Słońska, A.; Klimuszko, D. Bacteriocins produced by probiotic rods of the genus Lactobacillus. Postep. Mikrobiol. 2010, 49, 87–96. [Google Scholar]
- Herbel, S.R.; Vahjen, W.; Wieler, L.H.; Guenther, S. Timely approaches to identify probiotic species of the genus Lactobacillus. Gut Pathog. 2013, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.S.; Kim, I.H. Effect of Bacillus subtilis C-3102 spores as a probiotic feed supplement on growth performance, noxious gas emission, and intestinal microflora in broilers. Poult. Sci. 2014, 93, 3097–3103. [Google Scholar] [CrossRef]
- Zeyaullah, M.; Kamli, M.R.; Islam, B.; Atif, M.; Benkhayal, F.A.; Nehal, M.; Rizvi, M.A.; Ali, A. Metagenomics -An advanced approach for non- cultivable micro-organisms. Biotechnol. Mol. Biol. Rev. 2009, 4, 49–54. [Google Scholar]
- Lamendella, R.; Santo Domingo, J.W.; Ghosh, S.; Martinson, J.; Oerther, D.B. Comparative fecal metagenomics unveils unique functional capacity of the swine gut. BMC Microbiol. 2011, 11. [Google Scholar] [CrossRef] [Green Version]
- Sekirov, I.; Gill, N.; Jogova, M.; Tam, N.; Robertson, M.; de Llanos, R.; Li, Y.; Finlay, B.B. Salmonella SPI-1-mediated neutrophil recruitment during enteric colitis is associated with reduction and alteration in intestinal microbiota. Gut Microbes 2010, 1, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Pajarillo, E.A.B.; Chae, J.P.; Balolong, M.P.; Kim, H.B.; Park, C.S.; Kang, D.K. Effects of probiotic Enterococcus faecium NCIMB 11181 administration on swine fecal microbiota diversity and composition using barcoded pyrosequencing. Anim. Feed Sci. Technol. 2015, 201, 80–88. [Google Scholar] [CrossRef]
- Zhu, N.; Wang, J.; Yu, L.; Zhang, Q.; Chen, K.; Liu, B. Modulation of growth performance and intestinal microbiota in chickens fed plant extracts or virginiamycin. Front. Microbiol. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, C.; Selvaraj, J.; Gomathinayagam, S.; Thangapandiyan, M.; Ravikumar, G.; Roy, P.; Balachandran, C. Blastocystis sp. from food animals in India. J. Parasit. Dis. 2014, 38, 440–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jazi, V.; Boldaji, F.; Dastar, B.; Hashemi, S.R.; Ashayerizadeh, A. Effects of fermented cottonseed meal on the growth performance, gastrointestinal microflora population and small intestinal morphology in broiler chickens. Br. Poult. Sci. 2017, 58, 402–408. [Google Scholar] [CrossRef]
- Chu, Y.T.; Lo, C.T.; Chang, S.C.; Lee, T.T. Effects of Trichoderma fermented wheat bran on growth performance, intestinal morphology and histological findings in broiler chickens. Ital. J. Anim. Sci. 2017, 16, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Chiang, G.; Lu, W.Q.; Piao, X.S.; Hu, J.K.; Gong, L.M.; Thacker, P.A. Effects of feeding solid-state fermented rapeseed meal on performance, nutrient digestibility, intestinal ecology and intestinal morphology of broiler chickens. Asian Australas. J. Anim. Sci. 2010, 23, 263–271. [Google Scholar] [CrossRef]
- Borsoi, A.; do Santos, L.R.; Rodrigues, L.B.; de Souza Moraes, H.L.; Salle, C.T.P.; do Nascimento, V.P. Behavior of salmonella heidelberg and salmonella enteritidis strains following broiler chick inoculation: Evaluation of cecal morphometry, liver and cecum bacterial counts and fecal excretion patterns. Braz. J. Microbiol. 2011, 42, 266–273. [Google Scholar] [CrossRef]
- Caspary, W.F. Physiology and pathophysiology of intestinal absorption. Am. J. Clin. Nutr. 1992, 55, S299–S308. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.D. Antibiotic use in animal feed and its impact on human healt. Nutr. Res. Rev. 2000, 13, 279–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croom, W.J.; McBride, B.; Bird, A.R.; Fan, Y.K.; Odle, J.; Froetschel, M.; Taylor, I.L. Regulation of intestinal glucose absorption: A new issue in animal science. Can. J. Anim. Sci. 1998, 78, 1–13. [Google Scholar] [CrossRef]
- Xu, Z.; Hu, C.; Xia, M.; Zhan, X.; Wang, M. Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers. Poult. Sci. 2003, 82, 1030–1036. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Amount (kg) |
|---|---|
| Maize | 361.87 |
| Wheat | 300.00 |
| Soybean meal 45 | 217.00 |
| Animal fat | 30.00 |
| Sunflower seed cake | 25.00 |
| Sunflower meal | 15.00 |
| Medium chain fatty acids | 12.00 |
| Guar 60 | 10.00 |
| Chalk | 7.30 |
| Dicalcium phosphate | 7.00 |
| L-methionine 99 | 2.40 |
| Salt | 2.10 |
| L-lysine SO4 | 2.00 |
| Premix | 2.00 |
| L-lysine 98 | 1.70 |
| Sodium bicarbonate (NAHCO3) | 1.20 |
| L-Threonine | 1.10 |
| Choline chloride 75% | 0.67 |
| Sacox 120/Kokcisan/Salinomax | 0.58 |
| Mycofix select 5.E | 0.50 |
| L-valine | 0.44 |
| Hiphos Liquid (L) (phytase) | 0.08 |
| Hostazym (xylanase) | 0.06 |
| Colony Color | MALDI Identification | Score Value |
|---|---|---|
| Violet | Salmonella spp. | 2.3–3.0 |
| Blue | Klebsiella pneumoniae | 2.3–3.0 |
| Green | Enterobacter cloacae | 2.3–3.0 |
| White | Escherichia coli | 2.3–3.0 |
| Day of Experiment | Group | Salmonella spp. | Klebsiella pneumoniae | Enterobacter cloacae | Escherichia coli |
|---|---|---|---|---|---|
| 21 | NC | ¼ | 0/4 | 0/4 | ¾ |
| 44 | NC | 0/4 | 2/4 | ¾ | ¾ |
| 44 | PC | 0/4 | 0/4 | ¾ | ¾ |
| 44 | 67 | 0/4 | 0/4 | 0/4 | 4/4 |
| 44 | 87Y | 0/4 | 0/4 | 0/4 | 4/4 |
| Microorganisms (log10 cfu/mg of Feces) | Start Day 21 | NC Day 44 | PC Day 44 | 67 Day 44 | 87Y Day 44 | SEM | p-Value |
|---|---|---|---|---|---|---|---|
| Escherichia coli (RS) | 2.90 Aa | 3.47 b | 3.06 AaBb | 3.56 B | 3.54 B | 0.06 | 0.000754 |
| Enterococcus faecalis (SB) | 1.23 | 0.90 | 1.99 | 1.00 | 2.58 | 0.22 | 0.0812 |
| Lactobacillus gasseri (SB) | 4.36 | 3.73 | 4.13 | 2.65 | 3.20 | 0.19 | 0.0574 |
| Streptococcus alactolyticus (SB) | 1.06 | 2.33 | 1.11 | 2.14 | 1.50 | 0.24 | 0.439 |
| Lactobacillus salivarus (SB) | 4.94 A | 2.94 B | 3.81 B | n.d. | 2.58 B | 0.26 | 0.00001 |
| Enterococcus faecium (SB) | 1.94 | 1.45 | 2.98 | n.d. | 1.25 | 0.23 | 0.175 |
| Lactobacillus crispatus/johnsonii (MRS) | 2.44 A | 2.61 A | 3.02 AB | 0.76 A | 4.41 B | 0.26 | 0.00001 |
| Lactobacillus reuterii (MRS) | 5.19 a | 4.19 A | 3.61 Ab | 5.53 B | n.d. | 0.28 | 0.00001 |
| Lactobacillus salivarus (MRS) | 4.21 A | 2.40 B | 4.24 A | 3.62 B | 3.58 B | 0.16 | 0.0047 |
| Chromogenic Medium | Colony Color | MALDI Identification | Score Value |
|---|---|---|---|
| StrepB agar | Blue | Enterococcus faecalis | 2.3–3.0 |
| Lilac | Enterococcus faecium | 2.3–3.0 | |
| Violet matte | Lactobacillus gasseri | 2.3–3.0 | |
| Light violet | Streptococcus alactolyticus | 1.7–1.99 | |
| Violet/slight pink | Lactobacillus salivarus | 2.0–2.29 | |
| MRS | Large white | Lactobacillus salivarus | 1.7–1.99 |
| Matt white | Lactobacillus crispatus/johnsonii | 2.3–3.0 | |
| Transparent | Lactobacillus reuterii | 2.0–2.29 |
| Sample Name | Reads | Classified | Unclassified |
|---|---|---|---|
| 21_NC | 66,689,620 | 14,624,156 (21.93%) | 52,065,464 (78.07%) |
| 44_NC | 59,653,504 | 11,070,866 (18.56%) | 48,582,638 (81.44%) |
| 44_PC | 52,907,882 | 13,069,908 (24.7%) | 39,837,974 (75.30%) |
| 44_67 | 62,205,312 | 17,429,830 (28.02%) | 44,775,482 (71.98%) |
| 44_87Y | 61,418,084 | 21,317,558 (34.71%) | 40,100,526 (65.29%) |
| Kingdom | 21_NC | 44_NC | 44_PC | 44_67 | 44_87Y |
|---|---|---|---|---|---|
| Archaea | 3436 (0.02%) | 25,676 (0.23%) | 3714 (0.03%) | 4010 (0.02%) | 4114 (0.02%) |
| Bacteria | 14,551,854 (99.51%) | 10,301,176 (93.06%) | 13,015,864 (99.59%) | 17,363,714 (99.62%) | 21,262,068 (99.74%) |
| Eukaryote | 19,684 (0.13%) | 663,974 (6.0%) | 6522 (0.05%) | 6582 (0.04%) | 6450 (0.03%) |
| Fungi | 6252 (0.04%) | 8746 (0.08%) | 4526 (0.03%) | 5142 (0.03%) | 4822 (0.02%) |
| Viruses | 2622 (0.02%) | 4970 (0.04%) | 5460 (0.04%) | 9946 (0.06%) | 1010 (0.00%) |
| Ambiguous | 40,308 (0.28%) | 65,324 (0.59%) | 33,822 (0.26%) | 40,440 (0.23%) | 39,094 (0.18%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szmigiel, I.; Konkol, D.; Korczyński, M.; Łukaszewicz, M.; Krasowska, A. Changes in the Microbial Composition of the Cecum and Histomorphometric Analysis of Its Epithelium in Broilers Fed with Feed Mixture Containing Fermented Rapeseed Meal. Microorganisms 2021, 9, 360. https://doi.org/10.3390/microorganisms9020360
Szmigiel I, Konkol D, Korczyński M, Łukaszewicz M, Krasowska A. Changes in the Microbial Composition of the Cecum and Histomorphometric Analysis of Its Epithelium in Broilers Fed with Feed Mixture Containing Fermented Rapeseed Meal. Microorganisms. 2021; 9(2):360. https://doi.org/10.3390/microorganisms9020360
Chicago/Turabian StyleSzmigiel, Ida, Damian Konkol, Mariusz Korczyński, Marcin Łukaszewicz, and Anna Krasowska. 2021. "Changes in the Microbial Composition of the Cecum and Histomorphometric Analysis of Its Epithelium in Broilers Fed with Feed Mixture Containing Fermented Rapeseed Meal" Microorganisms 9, no. 2: 360. https://doi.org/10.3390/microorganisms9020360
APA StyleSzmigiel, I., Konkol, D., Korczyński, M., Łukaszewicz, M., & Krasowska, A. (2021). Changes in the Microbial Composition of the Cecum and Histomorphometric Analysis of Its Epithelium in Broilers Fed with Feed Mixture Containing Fermented Rapeseed Meal. Microorganisms, 9(2), 360. https://doi.org/10.3390/microorganisms9020360