
How the Soil Microbial Communities and Activities Respond to Long-Term Heavy Metal Contamination in Electroplating Contaminated Site



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Collection
2.3. Soil Physicochemical and HMs
2.4. Microbial Biomass Carbon and Soil EAs
2.5. DNA Extraction and Sequencing
2.6. Data Analysis
3. Results and Discussion
3.1. Soil HMs Contamination Level and Physicochemical Properties
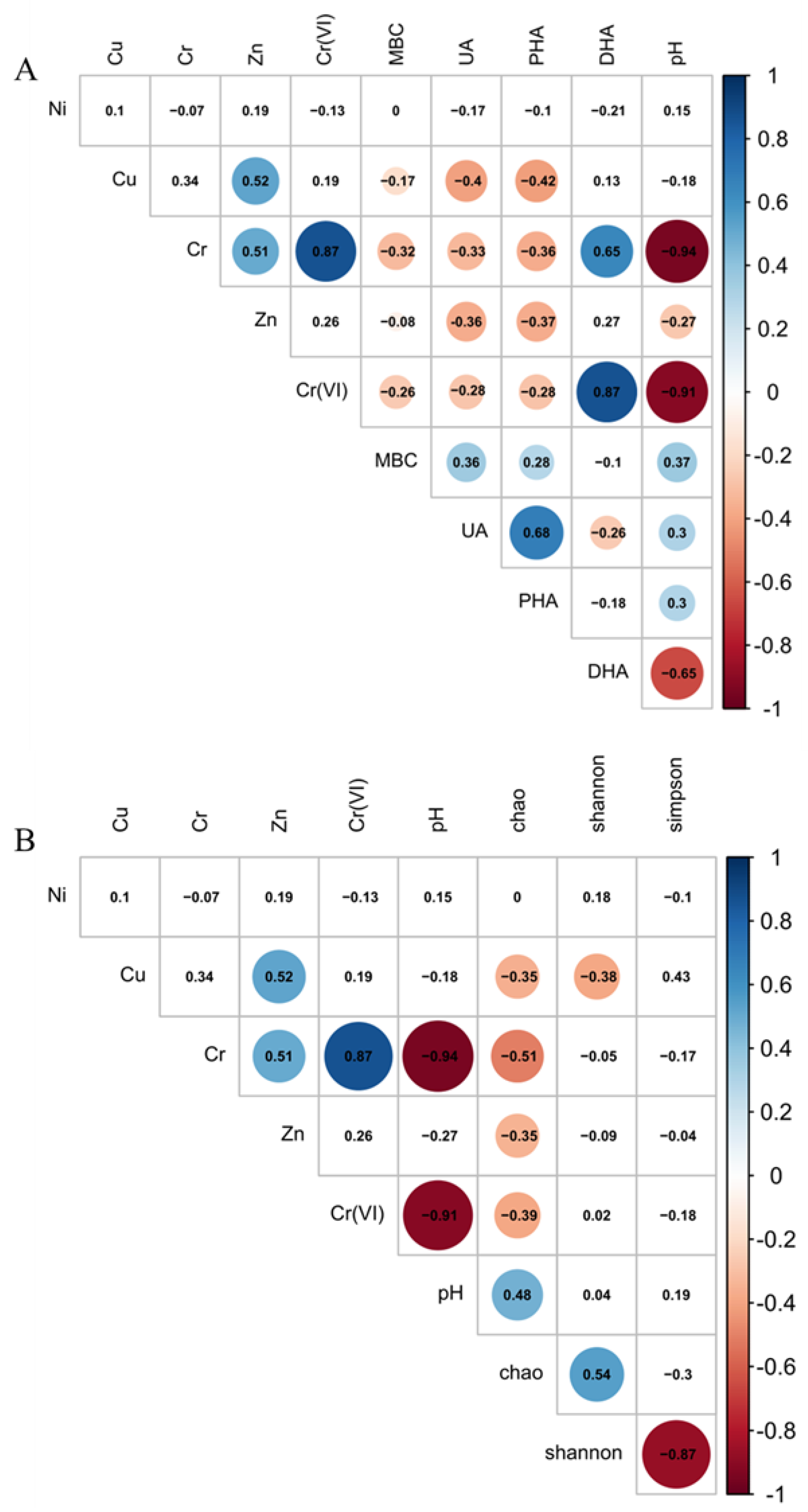
3.2. Relationship Between HMs, EAs, and MBC
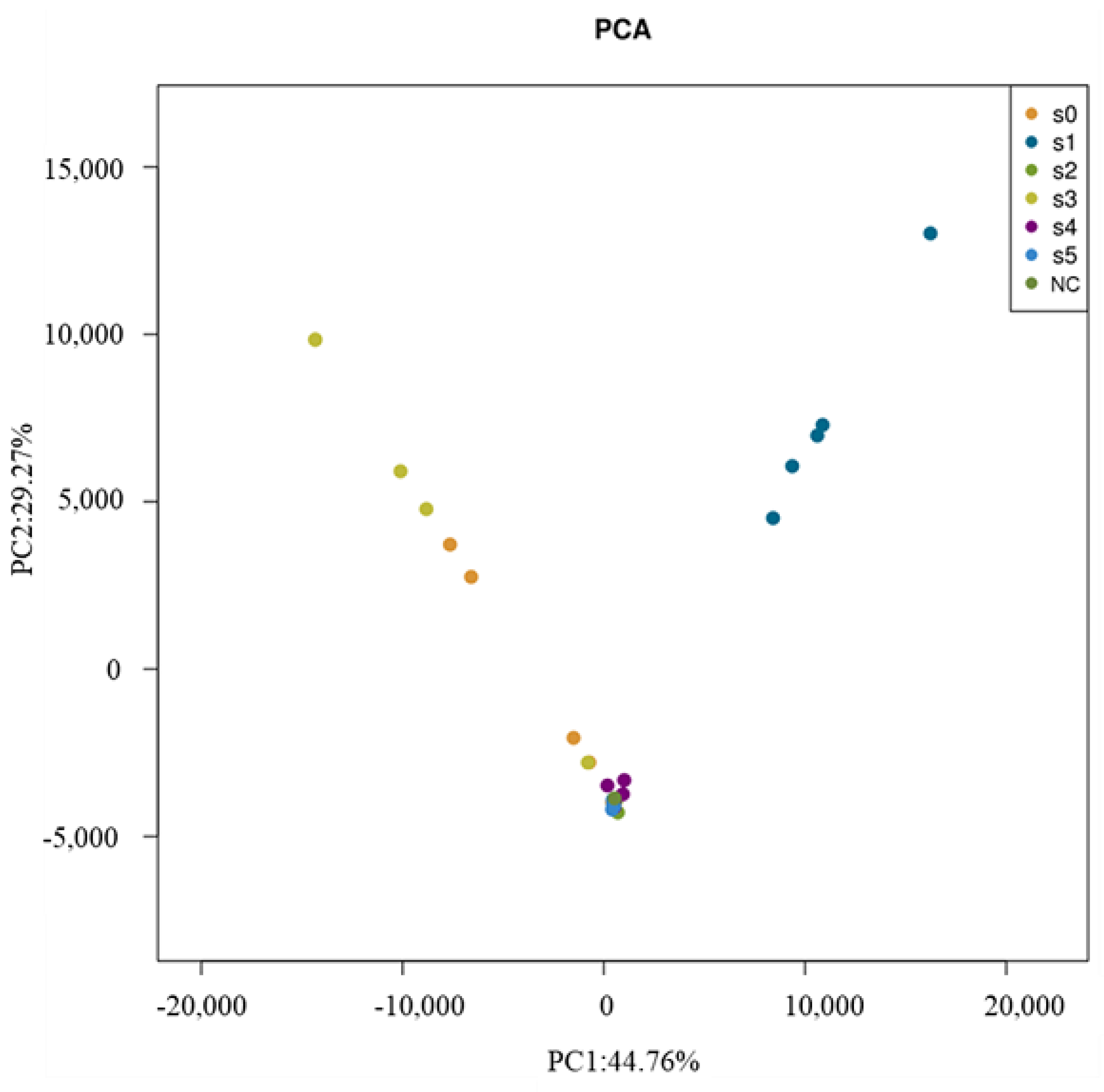
3.3. Relationship between Bacterial Diversity and HMs
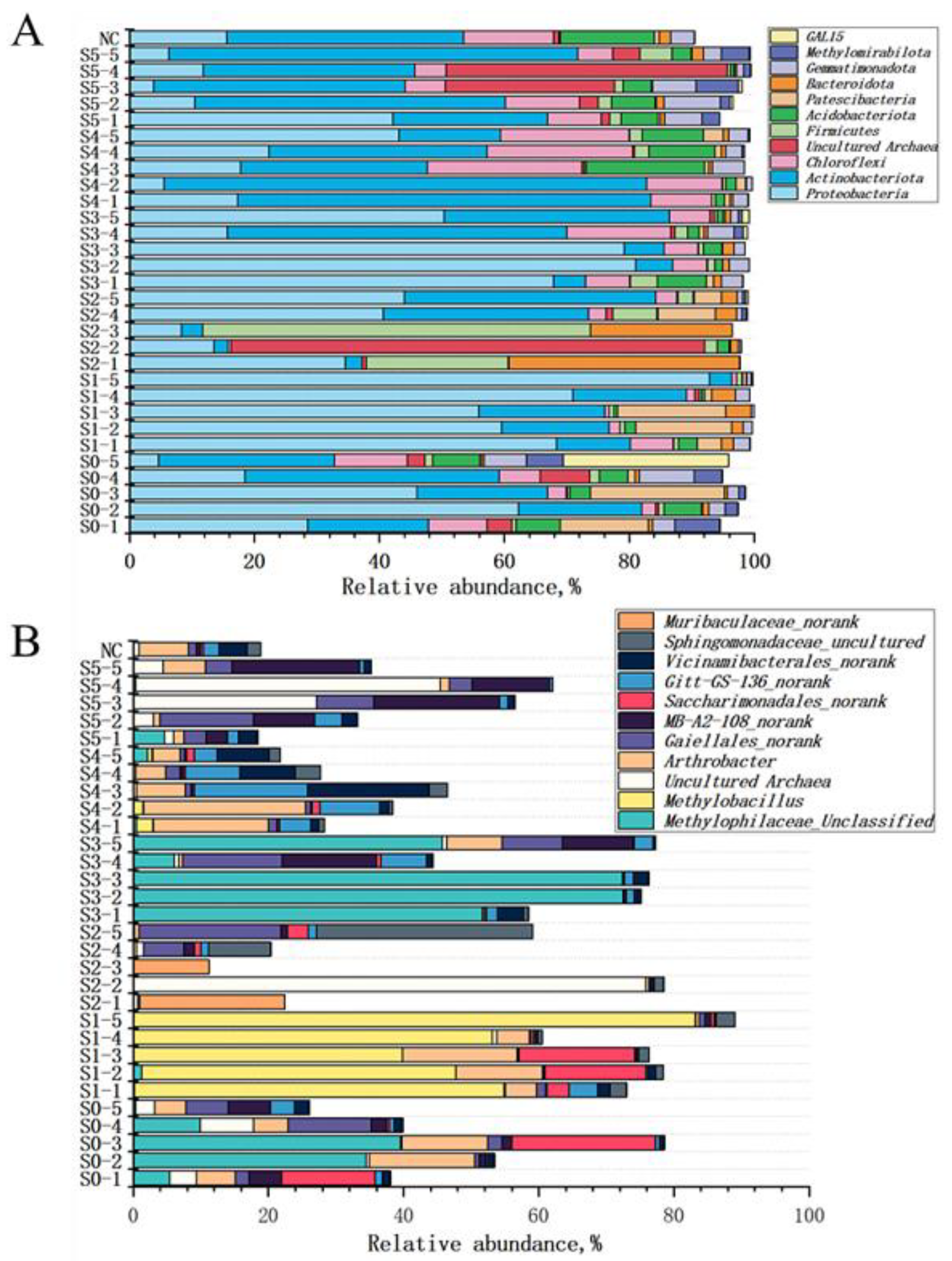
3.4. Relationship between Bacterial Community and HMs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hang, X.; Wang, H.; Zhou, J.; Du, C.; Chen, X. Characteristics and accumulation of heavy metals in sediments originated from an electroplating plant. J. Hazard. Mater. 2009, 163, 922–930. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, X.H.; Tran, H.; Wang, D.Q.; Zhu, Y.N. Heavy metal contamination and risk assessment in water, paddy soil, and rice around an electroplating plant. Environ. Sci. Pollut. Res. 2011, 18, 1623–1632. [Google Scholar] [CrossRef]
- Jordao, C.P.; Pereira MD, G.; Einloft, R.; Santana, M.B.; Bellato, C.R.; Vargas de Mello, J.W. Removal of Cu, Cr, Ni, Zn, and Cd from electroplating wastes and synthetic solutions by vermicompost of cattle manure. J. Environ. Sci. Health 2002, 37, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Kim, T.K.; Zoh, K.D. Removal mechanism of heavy metal (Cu, Ni, Zn, and Cr) in the presence of cyanide during electrocoagulation using Fe and Al electrodes. J. Water Process. Eng. 2020, 33, 101–109. [Google Scholar] [CrossRef]
- Pan, J.; Yu, L. Effects of Cd or/and Pb on soil enzyme activities and microbial community structure. Ecol. Eng. 2011, 37, 1889–1894. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, J.; Ren, L.; Zhou, Y.; Gao, J.; Luo, L.; Chen, A. Diagnosis of soil contamination using microbiological indices: A review on heavy metal pollution. J. Environ. Manag. 2019, 242, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Peng, B.; Yang, Z.; Chai, L.; Liao, Q.; Zhang, Z.; Li, C. Bacterial community dynamics during bioremediation of Cr (VI)-contaminated soil. Appl. Soil Ecol. 2015, 85, 50–55. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, S.; He, M. Bacterial community profile of contaminated soils in a typical antimony mining site. Environ. Sci. Pollut. Res. 2018, 25, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Ye, Y.; Hu, Y.; Shi, H. The variation in microbial community structure under different heavy metal contamination levels in paddy soils. Ecotoxicol. Environ. Saf. 2019, 180, 557–564. [Google Scholar] [CrossRef]
- Song, J.; Shen, Q.; Wang, L.; Qiu, G.; Shi, J.; Xu, J.; Liu, X. Effects of Cd, Cu, Zn and their combined action on microbial biomass and bacterial community structure. Environ. Pollut. 2018, 243, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Quan, Q.; Gan, Y.; Dong, J.; Fang, J.; Wang, L.; Liu, J. Effects of heavy metals on microbial communities in sediments and establishment of bioindicators based on microbial taxa and function for environmental monitoring and management. Sci. Total Environ. 2020, 749, 141–555. [Google Scholar] [CrossRef]
- Huang, A.; Xu, Y.; Sun, P.; Zhou, G.; Liu, C.; Lu, L.; Wang, H. Land use/land cover changes and its impact on ecosystem services in ecologically fragile zone: A case study of Zhangjiakou City, Hebei Province, China. Ecol. Indic. 2019, 104, 604–614. [Google Scholar] [CrossRef]
- Liu, C.; Gong, Z.; Teo, K.L.; Sun, J.; Caccetta, L. Robust multi-objective optimal switching control arising in 1, 3-propanediol microbial fed-batch process. Nonlinear Anal. Hybrid. Syst. 2017, 25, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Shi, J.; Wang, H.; Lin, Q.; Chen, X.; Chen, Y. The influence of soil heavy metals pollution on soil microbial biomass, enzyme activity, and community composition near a copper smelter. Ecotoxicol. Environ. Saf. 2007, 67, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Wong, Y.S.; Wong, M.H.; Tam, N.F.Y. Biosorption and bioreduction of Cr (VI) by a microalgal isolate, Chlorella miniata. J. Hazard. Mater. 2007, 146, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Li, F.D. Analysis of soil enzyme activity. In Experimental Techniques in Agricultural Microbiology; Chinese Agricultural Press: Beijing, China, 1996; pp. 137–139. [Google Scholar]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Zhao, H.P.; Ontiveros-Valencia, A.; Tang, Y.; Kim, B.O.; Ilhan, Z.E.; Krajmalnik-Brown, R.; Rittmann, B. Using a two-stage hydrogen-based membrane biofilm reactor (MBfR) to achieve complete perchlorate reduction in the presence of nitrate and sulfate. Environ. Sci. Technol. 2013, 47, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Ontiveros-Valencia, A.; Tang, Y.; Zhao, H.P.; Friese, D.; Overstreet, R.; Smith, J.; Krajmalnik-Brown, R. Pyrosequencing analysis yields comprehensive assessment of microbial communities in pilot-scale two-stage membrane biofilm reactors. Environ. Sci. Technol. 2014, 48, 7511–7518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Luo, Y.H.; Chen, J.X.; Zhang, Y.; Wen, L.L.; Shi, L.D.; Zhao, H.P. Evolution of the microbial community of the biofilm in a methane-based membrane biofilm reactor reducing multiple electron acceptors. Environ. Sci. Pollut. Res. 2016, 23, 9540–9548. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Lin, X.Y.; Chen, J.J.; Cai, C.Y.; Abbas, G.; Hu, Z.Q.; Zheng, P. Enrichment of denitratating bacteria from a methylotrophic denitrifying culture. Appl. Microbiol. Biotechnol. 2016, 100, 10203–10213. [Google Scholar] [CrossRef]
- Zhou, C.; Wang, Z.; Ontiveros-Valencia, A.; Long, M.; Lai, C.Y.; Zhao, H.P.; Rittmann, B.E. Coupling of Pd nanoparticles and denitrifying biofilm promotes H2-based nitrate removal with greater selectivity towards N2. Appl. Catal. B Environ. 2017, 206, 461–470. [Google Scholar] [CrossRef]
- Lai, C.Y.; Dong, Q.Y.; Rittmann, B.E.; Zhao, H.P. Bioreduction of antimonate by anaerobic methane oxidation in a membrane biofilm batch reactor. Environ. Sci. Technol. 2018, 52, 8693–8700. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.Y.; Lv, P.L.; Dong, Q.Y.; Yeo, S.L.; Rittmann, B.E.; Zhao, H.P. Bromate and nitrate bioreduction coupled with poly-β-hydroxybutyrate production in a methane-based membrane biofilm reactor. Environ. Sci. Technol. 2018, 52, 7024–7031. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, Y.; Lin, S.; Liu, Y.; Xie, Y. Soil pollution management in china: A brief introduction. Sustainability 2019, 11, 556. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Li, H.; Guo, G.; Semple, K.T.; Jones, K.C. Soil contamination in China: Current priorities, defining background levels and standards for heavy metals. J. Environ. Manag. 2019, 251. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Ali, S.; Zhang, H.; Ouyang, Y.; Qiu, B.; Wu, F.; Zhang, G. The influence of pH and organic matter content in paddy soil on heavy metal availability and their uptake by rice plants. Environ. Pollut. 2011, 159, 84–91. [Google Scholar] [CrossRef]
- Pattnaik, B.K.; Equeenuddin, S.M. Potentially toxic metal contamination and enzyme activities in soil around chromite mines at Sukinda Ultramafic Complex, India. J. Geochem. Explor. 2016, 168, 127–136. [Google Scholar] [CrossRef]
- Tyler, G.; Olsson, T. Concentrations of 60 elements in the soil solution as related to the soil acidity. Eur. J. Soil Sci. 2001, 52, 151–165. [Google Scholar] [CrossRef]
- Baena, C.W.; Andrés-Abellán, M.; Lucas-Borja, M.E.; Martínez-García, E.; García-Morote, F.A.; Rubio, E.; López-Serrano, F.R. Thinning and recovery effects on soil properties in two sites of a Mediterranean forest, in Cuenca Mountain (South-eastern of Spain). For. Ecol. Manag. 2013, 308, 223–230. [Google Scholar] [CrossRef]
- Jorge, I.; Soler, P.; Casermeiro, M.Á.; Cruz, M.T.; Polo, A. Seasonal changes in microbial activity in a semiarid soil after application of a high dose of different organic amendments. Geoderma 2013, 206, 40–48. [Google Scholar] [CrossRef]
- De Prada, P.; Brenchley, J.E. Purification and characterization of two extracellular alkaline phosphatases from a psychrophilic arthrobacter isolate. Appl. Environ. Microbiol. 1997, 63, 2928–2931. [Google Scholar] [CrossRef] [Green Version]
- Heyrman, J.; Verbeeren, J.; Schumann, P.; Swings, J.; De Vos, P. Six novel Arthrobacter species isolated from deteriorated mural paintings. Int. J. Syst. Evol. Microbiol. 2005, 55, 1457–1464. [Google Scholar] [CrossRef]
- Li, Y.T.; Zhang, H.H.; Sheng, H.M.; An, L.Z. Cloning, expression and characterization of trehalose-6-phosphate phosphatase from a psychrotrophic bacterium, Arthrobacter strain A3. World J. Microbiol. Biotechnol. 2012, 28, 2713–2721. [Google Scholar] [CrossRef]
- Aponte, H.; Herrera, W.; Cameron, C.; Black, H.; Meier, S.; Paolini, J.; Cornejo, P. Alteration of enzyme activities and functional diversity of a soil contaminated with copper and arsenic. Ecotoxicol. Environ. Saf. 2020, 192–200. [Google Scholar] [CrossRef]
- Mikanova, O. Effects of heavy metals on some soil biological parameters. J. Geochem. Explor. 2006, 88, 220–223. [Google Scholar] [CrossRef]
- Kumar, S.; Chaudhuri, S.; Maiti, S.K. Soil dehydrogenase enzyme activity in natural and mine soil-a review. Middle East. J. Sci. Res. 2013, 13, 898–906. [Google Scholar]
- Peng, B.; Huang, S.H.; Yang, Z.H.; Chai, L.Y.; Xu, Y.Z.; Su, C.Q. Inhibitory effect of Cr (VI) on activities of soil enzymes. J. Cent. South Univ. Technol. 2009, 16, 594–598. [Google Scholar] [CrossRef]
- Wyszkowska, J.; Kucharski, J.; Jastrzebska, E.; Hlasko, A. The biological properties of soil as influenced by chromium contamination. Pol. J. Environ. Stud. 2001, 10, 37–42. [Google Scholar]
- Zeiser, J.; Mühlenbeck, L.H.; Schweiger, P.; Deppenmeier, U. Characterization of a periplasmic quinoprotein from Sphingomonas wittichii that functions as aldehyde dehydrogenase. Appl. Microbiol. Biotechnol. 2014, 98, 2067–2079. [Google Scholar] [CrossRef] [PubMed]
- Bastida, F.; Jehmlich, N.; Lima, K.; Morris BE, L.; Richnow, H.H.; Hernández, T.; García, C. The ecological and physiological responses of the microbial community from a semiarid soil to hydrocarbon contamination and its bioremediation using compost amendment. J. Proteom. 2016, 135, 162–169. [Google Scholar] [CrossRef]
- Liang, Y.; Yang, Y.; Yang, C.; Shen, Q.; Zhou, J.; Yang, L. Soil enzymatic activity and growth of rice and barley as influenced by organic manure in an anthropogenic soil. Geoderma 2003, 115, 149–160. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, L.; Liu, Y.; Xie, X.; Zhou, J.; Jia, H.; Wei, P. Enhancing Cu-Zn-Cr-Ni Co-extraction from electroplating sludge in acid leaching process by optimizing Fe3+ addition and redox potential. Environ. Eng. Sci. 2019, 36, 1244–1257. [Google Scholar] [CrossRef]
- Samborska, A.; Stepniewska, Z.; Stepniewski, W. Influence of different oxidation states of chromium (VI, III) on soil urease activity. Geoderma 2004, 122, 317–322. [Google Scholar] [CrossRef]
- Hu, X.F.; Jiang, Y.; Shu, Y.; Hu, X.; Liu, L.; Luo, F. Effects of mining wastewater discharges on heavy metal pollution and soil enzyme activity of the paddy fields. J. Geochem. Explor. 2014, 147, 139–150. [Google Scholar] [CrossRef]
- Huber, K.J.; Overmann, J. Luteitalea. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc., in Association with Bergey’s Manual Trust: Hoboken, NJ, USA, 2015; pp. 1–5. [Google Scholar]
- Sheu, C.; Cai, C.Y.; Sheu, S.Y.; Li, Z.H.; Chen, W.M. Pseudomethylobacillus aquaticus gen. nov., sp. nov., a new member of the family Methylophilaceae isolated from an artificial reservoir. Int. J. Syst. Evol. Microbiol. 2019, 69, 3551–3559. [Google Scholar] [CrossRef] [PubMed]
- Editorial Board. Methylophilus. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc., in Association with Bergey’s Manual Trust: Hoboken, NJ, USA, 2015; pp. 1–3. [Google Scholar]
- Charlton, A.; Sakrabani, R.; Tyrrel, S.; Casado, M.R.; McGrath, S.P.; Crooks, B.; Campbell, C.D. Long-term impact of sewage sludge application on soil microbial biomass: An evaluation using meta-analysis. Environ. Pollut. 2016, 219, 1021–1035. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.M.; Qi, L.I.; Liang, W.J.; Jiang, Y. Distribution of soil enzyme activities and microbial biomass along a latitudinal gradient in farmlands of Songliao Plain, Northeast China. Pedosphere 2008, 18, 431–440. [Google Scholar] [CrossRef]
- Yu, Z.; Li, Y.; Wang, G.; Liu, J.; Liu, J.; Liu, X.; Herbert, S.J.; Jin, J. Effectiveness of elevated CO2 mediating bacterial communities in the soybean rhizosphere depends on genotypes. Agric. Ecosyst. Environ. 2016, 231, 229–232. [Google Scholar] [CrossRef]
- Zhang, C.; Nie, S.; Liang, J.; Zeng, G.; Wu, H.; Hua, S.; Xiang, H. Effects of heavy metals and soil physicochemical properties on wetland soil microbial biomass and bacterial community structure. Sci. Total Environ. 2016, 557, 785–790. [Google Scholar] [CrossRef]
- Abaye, D.A.; Lawlor, K.; Hirsch, P.R.; Brookes, P.C. Changes in the microbial community of an arable soil caused by long-term metal contamination. Eur. J. Soil Sci. 2005, 56, 93–102. [Google Scholar] [CrossRef]
- Muhammad, A.; Xu, J.; Li, Z.; Wang, H.; Yao, H. Effects of lead and cadmium nitrate on biomass and substrate utilization pattern of soil microbial communities. Chemosphere 2005, 60, 508–514. [Google Scholar] [CrossRef]
- Xu, Y.; Seshadri, B.; Sarkar, B.; Wang, H.; Rumpel, C.; Sparks, D.; Bolan, N. Biochar modulates heavy metal toxicity and improves microbial carbon use efficiency in soil. Sci. Total Environ. 2018, 621, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.D.; Zhu, Y.G.; Fu, B.J.; Marschner, P.; He, J.Z. The veterinary antibiotic oxytetracycline and Cu influence functional diversity of the soil microbial community. Environ. Pollut. 2006, 143, 129–137. [Google Scholar] [CrossRef]
- Zhao, X.; Huang, J.; Lu, J.; Sun, Y. Study on the influence of soil microbial community on the long-term heavy metal pollution of different land use types and depth layers in mine. Ecotoxicol. Environ. Saf. 2019, 170, 218–226. [Google Scholar] [CrossRef]
- De Oliveira, A.J.F.C.; Hollnagel, H.C.; Mesquita, H.D.S.L.; Fontes, R.F.C. Physical, chemical and microbiological characterization of the intertidal sediments of Pereque Beach, Guarujá (SP), Brazil. Mar. Pollut. Bull. 2007, 54, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Sheik, C.S.; Mitchell, T.W.; Rizvi, F.Z.; Rehman, Y.; Faisal, M.; Hasnain, S.; Krumholz, L.R. Exposure of soil microbial communities to chromium and arsenic alters their diversity and structure. PLoS ONE 2012, 7, 40–59. [Google Scholar]
- Schneider, A.R.; Gommeaux, M.; Duclercq, J.; Fanin, N.; Conreux, A.; Alahmad, A.; Marin, B. Response of bacterial communities to Pb smelter pollution in contrasting soils. Sci. Total Environ. 2017, 605, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Jiang, K.; Zhou, J.; Liu, J.; Wu, B. Responses of soil N-fixing bacterial communities to redroot pigweed (Amaranthus retroflexus L.) invasion under Cu and Cd heavy metal soil pollution. Agric. Ecosyst. Environ. 2018, 267, 15–22. [Google Scholar] [CrossRef]
- Li, Q.; You, P.; Hu, Q.; Leng, B.; Wang, J.; Chen, J.; Ouyang, K. Effects of co-contamination of heavy metals and total petroleum hydrocarbons on soil bacterial community and function network reconstitution. Ecotoxicol. Environ. Saf. 2020, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Viti, C.; Decorosi, F.; Mini, A.; Tatti, E.; Giovannetti, L. Involvement of the oscA gene in the sulphur starvation response and in Cr (VI) resistance in Pseudomonas corrugata 28. Microbiology 2009, 155, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Ye, B.; Xia, S. Structural characteristics of cake layer in membrane bioreactor with chromate exposure. Ecotoxicol. Environ. Saf. 2019, 169, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Su, G.; Yang, Y.; Yao, Y.; Huang, Y.; Hu, L.; He, Z. Vertical distribution of microbial communities in chromium-contaminated soil and isolation of Cr (Ⅵ)-Reducing strains. Ecotoxicol. Environ. Saf. 2019, 180, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Tao, Q.; Wu, K.; Li, J.; Qian, J.; Liang, Y.; Li, T. Structural and functional variability in root-associated bacterial microbiomes of Cd/Zn hyperaccumulator Sedum alfredii. Appl. Microbiol. Biotechnol. 2017, 101, 7961–7976. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wu, J.; Huo, Y.; Zhao, X.; Xue, L. Profiling multiple heavy metal contamination and bacterial communities surrounding an iron tailing pond in Northwest China. Sci. Total Environ. 2020, 752–761. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Ni (mg/kg) | Total Cu (mg/kg) | Total Cr (mg/kg) | Total Zn (mg/kg) | Total Cr (VI) (mg/kg) | |
|---|---|---|---|---|---|
| S0-1 | 24.8 ± 0.72 | 38.0 ± 1.04 | 411.2 ± 0.72 | 83.6 ± 1.20 | 1.4 ± 0.28 |
| S0-2 | 22.0 ± 0.40 | 20.0 ± 0.28 | 172.4 ± 0.44 | 83.6 ± 0.52 | 3.2 ± 0.34 |
| S0-3 | 28.0 ± 0.16 | 29.2 ± 0.72 | 232.4 ± 0.80 | 105.6 ± 0.56 | 1.3 ± 0.37 |
| S0-4 | 28.0 ± 0.52 | 20.0 ± 0.04 | 292.0 ± 0.68 | 91.2 ± 0.16 | 2.2 ± 0.22 |
| S0-5 | 34.0 ± 1.52 | 35.2 ± 0.28 | 292.0 ± 0.20 | 124.8 ± 0.36 | 5.0 ± 0.64 |
| S1-1 | 30.8 ± 0.56 | 16.8 ± 0.36 | 292.0 ± 0.20 | 96.4 ± 0.60 | 6.5 ± 0.68 |
| S1-2 | 15.6 ± 0.20 | 16.8 ± 0.16 | 292.0 ± 0.36 | 143.6 ± 0.40 | 4.2 ± 0.20 |
| S1-3 | 24.8 ± 0.76 | 10.8 ± 0.04 | 351.6 ± 0.44 | 69.6 ± 0.60 | 13.0 ± 0.22 |
| S1-4 | 30.8 ± 0.72 | 486.0 ± 0.60 | 5786.0 ± 7.76 | 590.4 ± 0.52 | 104.5 ± 2.60 |
| S1-5 | 24.8 ± 0.84 | 16.8 ± 0.16 | 351.6 ± 0.32 | 165.2 ± 0.16 | 82.0 ± 1.60 |
| S2-1 | 18.8 ± 1.12 | 126.0 ± 0.08 | 7338.8 ± 4.76 | 258.0 ± 0.72 | 302.6 ± 1.04 |
| S2-2 | 37.2 ± 0.40 | 132.0 ± 0.12 | 9727.2 ± 4.80 | 245.2 ± 2.40 | 861.9 ± 2.80 |
| S2-3 | 28.0 ± 0.52 | 147.2 ± 0.28 | 9249.6 ± 6.44 | 251.2 ± 1.04 | 861.9 ± 3.40 |
| S2-4 | 22.0 ± 0.40 | 219.6 ± 0.36 | 6681.6 ± 3.80 | 304.4 ± 0.60 | 1083.3 ± 4.40 |
| S2-5 | 28.0 ± 0.52 | 23.2 ± 0.36 | 471.2 ± 0.24 | 133.2 ± 1.04 | 47.5 ± 1.04 |
| S3-1 | 30.8 ± 0.32 | 258.8 ± 0.52 | 530.8 ± 0.20 | 156.0 ± 1.20 | 3.5 ± 0.24 |
| S3-2 | 40.0 ± 0.28 | 376.8 ± 0.64 | 530.8 ± 0.24 | 191.6 ± 0.24 | 5.2 ± 0.04 |
| S3-3 | 30.8 ± 0.24 | 510.0 ± 1.12 | 351.6 ± 0.24 | 121.6 ± 1.16 | 2.3 ± 0.04 |
| S3-4 | 28.0 ± 0.20 | 86.4 ± 0.16 | 351.6 ± 0.52 | 98.8 ± 0.40 | 3.3 ± 0.68 |
| S3-5 | 22.0 ± 0.76 | 20.0 ± 0.08 | 172.4 ± 0.44 | 123.2 ± 1.64 | 0.5 ± 0.44 |
| S4-1 | 46.4 ± 0.56 | 189.2 ± 0.40 | 2859.6 ± 2.48 | 631.6 ± 2.08 | 0.1 ± 0.20 |
| S4-2 | 28.0 ± 0.52 | 98.8 ± 0.12 | 590.4 ± 0.16 | 310.0 ± 2.60 | 23.3 ± 0.72 |
| S4-3 | 40.0 ± 0.76 | 92.4 ± 0.24 | 590.4 ± 0.32 | 327.6 ± 0.88 | 1.8 ± 0.22 |
| S4-4 | 22.0 ± 0.72 | 122.8 ± 0.76 | 351.6 ± 0.72 | 192.4 ± 0.88 | 7.4 ± 0.80 |
| S4-5 | 30.8 ± 0.72 | 77.6 ± 3.48 | 411.2 ± 0.32 | 192.4 ± 0.40 | 8.4 ± 0.96 |
| S5-1 | 24.8 ± 0.12 | 16.8 ± 0.20 | 112.8 ± 1.04 | 128.4 ± 2.08 | 3.5 ± 0.34 |
| S5-2 | 30.8 ± 0.56 | 16.8 ± 0.16 | 292.0 ± 0.36 | 106.4 ± 1.80 | 1.0 ± 0.28 |
| S5-3 | 37.2 ± 0.32 | 16.8 ± 0.08 | 351.6 ± 0.28 | 104.0 ± 0.20 | 2.1 ± 0.28 |
| S5-4 | 58.4 ± 0.36 | 20.0 ± 0.04 | 232.4 ± 0.52 | 98.8 ± 0.40 | 0 |
| S5-5 | 34.0 ± 0.52 | 10.8 ± 0.08 | 172.4 ± 0.80 | 148.4 ± 3.92 | 0 |
| NC | 43.2 ± 0.76 | 20.0 ± 0.03 | 292.0 ± 0.52 | 114.8 ± 0.19 | 0 |
| Methylophilaceae_Unclassified | Methylobacillus | Arthrobacter | MB-A2-108_Norank | Vicinamibacterales_Norank | Sphingomonadaceae_Uncultured | Muribaculaceae_Norank | |
|---|---|---|---|---|---|---|---|
| Ni | 0.100 | −0.219 | −0.058 | 0.236 | 0.095 | −0.290 | −0.112 |
| Cu | 0.107 | 0.132 | −0.266 | −0.152 | −0.157 | 0.250 | 0.363 * |
| Cr | −0.338 | 0.294 | −0.285 | −0.426 * | −0.377 * | 0.406 * | 0.402 * |
| Zn | −0.345 | 0.259 | −0.246 | −0.367 * | 0.097 | 0.574 ** | 0.312 |
| Cr (VI) | −0.444 * | 0.487 ** | −0.278 | −0.500 ** | −0.416 * | 0.456 * | 0.458 ** |
| pH | 0.558 ** | −0.167 | 0.404 * | 0.444 * | 0.393 * | −0.154 | −0.365 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, W.-J.; Niu, Z.-F.; Wang, X.-R.; Zhao, H.-P. How the Soil Microbial Communities and Activities Respond to Long-Term Heavy Metal Contamination in Electroplating Contaminated Site. Microorganisms 2021, 9, 362. https://doi.org/10.3390/microorganisms9020362
Gong W-J, Niu Z-F, Wang X-R, Zhao H-P. How the Soil Microbial Communities and Activities Respond to Long-Term Heavy Metal Contamination in Electroplating Contaminated Site. Microorganisms. 2021; 9(2):362. https://doi.org/10.3390/microorganisms9020362
Chicago/Turabian StyleGong, Wen-Jing, Zi-Fan Niu, Xing-Run Wang, and He-Ping Zhao. 2021. "How the Soil Microbial Communities and Activities Respond to Long-Term Heavy Metal Contamination in Electroplating Contaminated Site" Microorganisms 9, no. 2: 362. https://doi.org/10.3390/microorganisms9020362
APA StyleGong, W.-J., Niu, Z.-F., Wang, X.-R., & Zhao, H.-P. (2021). How the Soil Microbial Communities and Activities Respond to Long-Term Heavy Metal Contamination in Electroplating Contaminated Site. Microorganisms, 9(2), 362. https://doi.org/10.3390/microorganisms9020362